
Impact of Pheidole fallax (Hymenoptera: Formicidae) as an Ecosystem Engineer in Rehabilitated Coal Mine Areas

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of Ant Nests
2.2. Characterization and Biomass of Organic Debris of Refuse Piles
2.3. Chemical Properties of Ant Nets
2.4. Internal Structure of Nests and Calculation of Nest Volume
2.5. Data Analysis
3. Results
3.1. Characterization and Biomass of Organic Debris in Refuse Piles
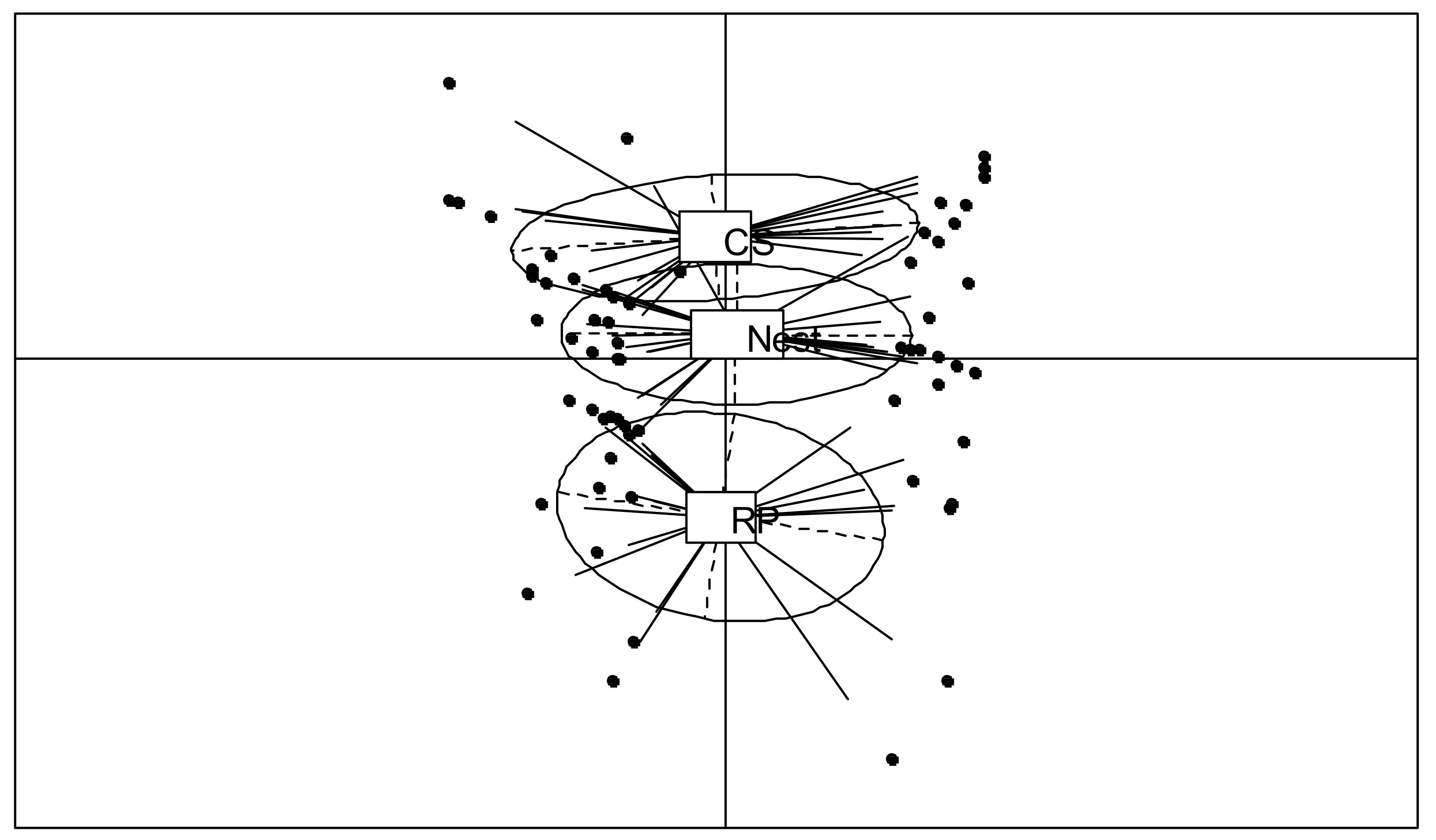
3.2. Chemical and NIRS Analyses
3.3. Nest Architecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mansourian, S.; Vallauri, D.; Dudley, N. Forest Restoration in Landscapes: Beyond Planting Trees; Springer: New York, NY, USA, 2005; ISBN 978-0-387-25525-5. [Google Scholar]
- Dominati, E.; Patterson, M.; Mackay, A. A framework for classifying and quantifying the natural capital and ecosystem services of soils. Ecol. Econ. 2010, 69, 1858–1868. [Google Scholar] [CrossRef]
- Prach, K.; Hobbs, R.J. Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor. Ecol. 2008, 16, 363–366. [Google Scholar] [CrossRef]
- Lavelle, P.; Spain, A.; Blouin, M.; Brown, G.; Decaëns, T.; Grimaldi, M.; Jiménez, J.J.; McKey, D.; Mathieu, J.; Velasquez, E.; et al. Ecosystem engineers in a self-organized soil: A Review of concepts and future research questions. Soil Sci. 2016, 181, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Lozano-Baez, S.E.; Domínguez-Haydar, Y.; Meli, P.; Meerveld, I.; Vásquez, K.V.; Castellini, M. Key gaps in soil monitoring during forest restoration in Colombia. Restor. Ecol. 2021, 29, e13391. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373. [Google Scholar] [CrossRef]
- Lee, K.; Foster, R. Soil Fauna and Soil Structure. Soil Res. 1991, 29, 745. [Google Scholar] [CrossRef]
- Dauber, J.; Niechoj, R.; Baltruschat, H.; Wolters, V. Soil engineering ants increase grass root arbuscular mycorrhizal colonization. Biol. Fertil. Soils 2008, 44, 791–796. [Google Scholar] [CrossRef]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Eldridge, D. Effect of ants on sandy soils in semi-arid eastern Australia—Local distribution of nest entrances and their effect on infiltration of water. Soil Res. 1993, 31, 509. [Google Scholar] [CrossRef]
- Jiménez, J.J.; Decaëns, T.; Lavelle, P. C and N concentrations in biogenic structures of a soil-feeding termite and a fungus-growing ant in the Colombian savannas. Appl. Soil Ecol. 2008, 40, 120–128. [Google Scholar] [CrossRef]
- Lobry de Bruyn, L.A. Ants as bioindicators of soil function in rural environments. Agric. Ecosyst. Environ. 1999, 74, 425–441. [Google Scholar] [CrossRef]
- Andersen, A.N.; Majer, J.D. Ants show the way down under: Invertebrates as bioindicators in land management. Front. Ecol. Environ. 2004, 2, 291–298. [Google Scholar] [CrossRef]
- Buchori, D.; Rizali, A.; Rahayu, G.A.; Mansur, I. Insect diversity in post-mining areas: Investigating their potential role as bioindicator of reclamation success. Biodiversitas 2018, 19, 1696–1702. [Google Scholar] [CrossRef]
- Domínguez-Haydar, Y.; Armbrecht, I. Response of ants and their seed removal in rehabilitation areas and forests at El Cerrejón coal mine in Colombia. Restor. Ecol. 2011, 19, 178–184. [Google Scholar] [CrossRef]
- Menta, C.; Conti, F.D.; Pinto, S.; Leoni, A.; Lozano-Fondón, C. Monitoring soil restoration in an open-pit mine in northern italy. Appl. Soil Ecol. 2014, 83, 22–29. [Google Scholar] [CrossRef]
- Frouz, J.; Holec, M.; Kalčík, J. The Effect of Lasius Niger (Hymenoptera, Formicidae) ant nest on selected soil chemical properties. Pedobiologia 2003, 47, 205–212. [Google Scholar] [CrossRef]
- Philpott, S.M.; Perfecto, I.; Armbrecht, I.; Parr, C.L. Ant diversity and function in disturbed and changing habitats. In Ant Ecology; Lach, L., Parr, C., Abbott, K., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 137–156. ISBN 978-0-19-954463-9. [Google Scholar]
- Leal, I.R.; Wirth, R.; Tabarelli, M. Seed dispersal by ants in the Semi-Arid Caatinga of North-East Brazil. Ann. Bot. 2007, 99, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Lôbo, D.; Tabarelli, M.; Leal, I. Relocation of Croton sonderianus (Euphorbiaceae) seeds by Pheidole fallax Mayr (Formicidae): A case of post-dispersal seed protection by ants? Neotrop. Entomol. 2011, 40, 440–444. [Google Scholar] [CrossRef] [Green Version]
- Itzkowitz, M.; Haley, M. The food retrieval tactics of the ant Pheidole fallax Mayr. Insectes Sociaux 1983, 30, 317–322. [Google Scholar] [CrossRef]
- Gutierrez Rapalino, B.P.; Domínguez Haydar, Y.D.C. Contribución de Pheidole fallax y Ectatomma ruidum (Hymenoptera: Formicidae) en la dispersión y germinación de semillas en áreas rehabilitadas de la mina de Carbón Del Cerrejón, Colombia. Rev. Biol. Tropical 2017, 65, 575–587. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.K.; Singh, H.; Rastogi, N.; Agarwal, V.M. Impact of abundant Pheidole ant species on soil nutrients in relation to the food biology of the species. Appl. Soil Ecol. 2013, 71, 15–23. [Google Scholar] [CrossRef]
- Gualdrón Acosta, R. Cerrejón: Hacia La Rehabilitación de Las Tierras Intervenidas Por La Minería a Cielo Abierto; Cerrejón: Bogotá, Colombia, 2011; ISBN 978-958-99731-8-9. [Google Scholar]
- Domínguez-Haydar, Y.; Gutierrez-Rapalino, B.; Jiménez, J.J. Density and spatial distribution of nests of Ectatomma ruidum and Pheidole fallax (Hymenoptera: Formicidae), as response to the recovery of coal mine areas. Sociobiology 2018, 65, 415. [Google Scholar] [CrossRef] [Green Version]
- Ben-Dor, E.; Banin, A. Near-infrared analysis as a rapid method to simultaneously evaluate several soil properties. Soil Sci. Soc. Am. J. 1995, 59, 364–372. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- EMBRAPA. Manual of methods of soil Analysis, 2nd ed.; Embrapa Soils: Rio de Janeiro, Brazil, 2011. [Google Scholar]
- Zangerlé, A.; Hissler, C.; Blouin, M.; Lavelle, P. Near infrared spectroscopy (nirs) to estimate earthworm cast age. Soil Biol. Biochem. 2014, 70, 47–53. [Google Scholar] [CrossRef]
- De Carvalho Guimarães, I.; Pereira, M.C.; Batista, N.R.; Rodrigues, C.A.P.; Antonialli, W.F. The complex nest architecture of the Ponerinae ant Odontomachus chelifer. PLoS ONE 2018, 13, e0189896. [Google Scholar] [CrossRef] [Green Version]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMRplus). Available online: https://cran.r-project.org/web/packages/PMCMRplus/PMCMRplus.pdf (accessed on 23 January 2022).
- Wittkowski, K.M.; Song, T. Package MuStat: Prentice Rank Sum Test and McNemar Test. Available online: https://cran.r-project.org/web/packages/muStat/muStat.pdf (accessed on 23 January 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Viena, Austria, 2021. [Google Scholar]
- Majer, J.D. Animals in the restoration process-progressing the trends. Restor. Ecol. 2009, 17, 315–319. [Google Scholar] [CrossRef]
- Agarwal, V.M.; Rastogi, N.; Raju, S.V.S. Impact of predatory ants on two Lepidopteran insect pests in Indian cauliflower agroecosystems. J. Appl. Entomol. 2007, 131, 493–500. [Google Scholar] [CrossRef]
- Cammeraat, L.H.; Willott, S.J.; Compton, S.G.; Incoll, L.D. The effects of ants’ nests on the physical, chemical and hydrological properties of a rangeland Soil in Semi-Arid Spain. Geoderma 2002, 105, 1–20. [Google Scholar] [CrossRef]
- Moutinho, P.; Nepstad, D.C.; Davidson, E.A. Influence of leaf-cutting ant nests on secondary forest growth and soil properties in amazonia. Ecology 2003, 84, 1265–1276. [Google Scholar] [CrossRef]
- Wagner, D.; Brown, M.J.F.; Gordon, D.M. Harvester Ant nests, soil biota and soil chemistry. Oecologia 1997, 112, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Holec, M.; Frouz, J. The effect of two ant species lasius niger and lasius flavus on soil properties in two contrasting habitats. Eur. J. Soil Biol. 2006, 42, S213–S217. [Google Scholar] [CrossRef]
- Levan, M.A.; Stone, E.L. Soil modification by colonies of black meadow ants in a New York old field. Soil Sci. Soc. Am. J. 1983, 47, 1192. [Google Scholar] [CrossRef]
- Domínguez-Haydar, Y.; Velásquez, E.; Zangerlé, A.; Lavelle, P.; Gutiérrez-Eisman, S.; Jiménez, J.J. Unveiling the age and origin of biogenic aggregates produced by earthworm species with their NIRS Fingerprint in a Subalpine Meadow of Central Pyrenees. PLoS ONE 2020, 15, e0237115. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, E.; Pelosi, C.; Brunet, D.; Grimaldi, M.; Martins, M.; Rendeiro, A.C.; Barrios, E.; Lavelle, P. This ped is my ped: Visual separation and near infrared spectra allow determination of the origins of soil macroaggregates. Pedobiologia 2007, 51, 75–87. [Google Scholar] [CrossRef]
- Tschinkel, W.R. The nest architecture of the ant, camponotus socius. J. Insect Sci. 2005, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Forti, L.C.; Camargo, R.S.; Fujihara, R.T.; Lopes, J.F.S. The nest architecture of the ant, Pheidole oxyops Forel (Hymenoptera: Formicidae). Insect Sci. 2007, 14, 437–442. [Google Scholar] [CrossRef]
- Domínguez-Haydar, Y.; Velásquez, E.; Carmona, J.; Lavelle, P.; Chavez, L.F.; Jiménez, J.J. Evaluation of reclamation success in an open-pit coal mine using integrated soil physical, chemical and biological quality indicators. Ecol. Indic. 2019, 103, 182–193. [Google Scholar] [CrossRef]
- Farji-Brener, A.G.; Werenkraut, V. The effects of ant nests on soil fertility and plant performance: A meta-analysis. J. Anim. Ecol. 2017, 86, 866–877. [Google Scholar] [CrossRef]
- Leite, P.A.M.; Carvalho, M.C.; Wilcox, B.P. Good ant, bad ant? Soil engineering by ants in the brazilian caatinga differs by species. Geoderma 2018, 323, 65–73. [Google Scholar] [CrossRef]
- Wang, D.; Lowery, B.; Norman, J.M.; McSweeney, K. Ant burrow effects on water flow and soil hydraulic properties of sparta sand. Soil Tillage Res. 1996, 37, 83–93. [Google Scholar] [CrossRef]
- Nkem, J.N.; Lobry de Bruyn, L.A.; Grant, C.D.; Hulugalle, N.R. The impact of ant bioturbation and foraging activities on surrounding soil properties. Pedobiologia 2000, 44, 609–621. [Google Scholar] [CrossRef]
- O’Grady, A.; Breen, J.; Harrington, T.J.; Courtney, R. The seed bank in soil from the nests of grassland ants in a unique limestone grassland community in Ireland. Ecol. Eng. 2013, 61, 58–64. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | n | Seeds (g) | AD (g) | OAD (g) | p-Value |
|---|---|---|---|---|---|
| 16 years | 15 | 0.27 a | 0.36 b | 0.03 b | 0.001 |
| 0.09 | 0.07 | 0.01 | |||
| 20 years | 15 | 0.41 a | 0.64 a | 0.12 b | 0.001 |
| 0.20 | 0.21 | 0.03 |
| Refuse Pile | Nests | Control Soil | Increase/Decrease RP vs. CS (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | P | |||
| pH | 6.70 a | 0.17 | 7.83 b | 0.15 | 7.84 b | 0.06 | <0.01 | ↓ | 14.54 |
| OM (%) | 9.36 a | 1.30 | 3.32 b | 0.10 | 3.28 b | 0.31 | <0.01 | ↑ | 64.96 |
| N (%) | 0.98 a | 0.29 | 0.30 b | 0.10 | 0.29 b | 0.07 | <0.01 | ↑ | 70.41 |
| P (ppm) | 132.29 a | 7.85 | 46.54 b | 9.31 | 54.11 b | 13.03 | <0.01 | ↑ | 59.10 |
| Ca (meq) | 39.19 a | 7.28 | 56.60 b | 8.53 | 54.6 b | 8.31 | |||
| Mg (meq) | 10.13 a | 1.14 | 4.28 b | 0.19 | 4.36 b | 0.25 | <0.01 | ↑ | 56.96 |
| K (meq) | 5.43 a | 0.31 | 2.46 b | 0.16 | 2.53 b | 0.25 | <0.01 | ↑ | 53.41 |
| Nest | NC | TH (cm) | TV (cm3) | AVC |
|---|---|---|---|---|
| 1 | 7 | 4.7 | 30.4 | 16.66 ± 0.62 |
| 2 | 9 | 6.2 | 40.1 | 12.08 ± 0.29 |
| 3 | 12 | 8.4 | 55.6 | 15.94 ± 0.11 |
| 4 | 14 | 8.9 | 90.4 | 14.13 ± 0.15 |
| 5 | 17 | 10.3 | 105.7 | 17.41 ± 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Haydar, Y.; Gutierrez-Rapalino, B.P.; Barros-Torres, Y.; Jiménez, J.J.; Lozano-Baez, S.E.; Castellini, M. Impact of Pheidole fallax (Hymenoptera: Formicidae) as an Ecosystem Engineer in Rehabilitated Coal Mine Areas. Appl. Sci. 2022, 12, 1573. https://doi.org/10.3390/app12031573
Domínguez-Haydar Y, Gutierrez-Rapalino BP, Barros-Torres Y, Jiménez JJ, Lozano-Baez SE, Castellini M. Impact of Pheidole fallax (Hymenoptera: Formicidae) as an Ecosystem Engineer in Rehabilitated Coal Mine Areas. Applied Sciences. 2022; 12(3):1573. https://doi.org/10.3390/app12031573
Chicago/Turabian StyleDomínguez-Haydar, Yamileth, Bleydis Paola Gutierrez-Rapalino, Yair Barros-Torres, Juan José Jiménez, Sergio Esteban Lozano-Baez, and Mirko Castellini. 2022. "Impact of Pheidole fallax (Hymenoptera: Formicidae) as an Ecosystem Engineer in Rehabilitated Coal Mine Areas" Applied Sciences 12, no. 3: 1573. https://doi.org/10.3390/app12031573