
Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Retrieval of Rubber Tree MicroRNAs
2.2. RTV1 Genome Retrieval and Annotation
2.3. miRanda
2.4. RNA22
2.5. RNAhybrid
2.6. Tapirhybrid
2.7. psRNATarget
2.8. Mapping of miRNA–Target Interaction
2.9. RNAfold and RNAcofold
2.10. Graphical Representation
3. Results
3.1. Rubber Tree miRNA’s Loci on RTV1 Genome
3.2. ORF1 Encoding Polyprotein
3.3. ORF2 Encoding Movement Protein
3.4. Visualization of miRNA–Target Interaction Network
3.5. Predicting Common Rubber Tree miRNAs
3.6. Prediction of Consensual Rubber Tree miRNAs
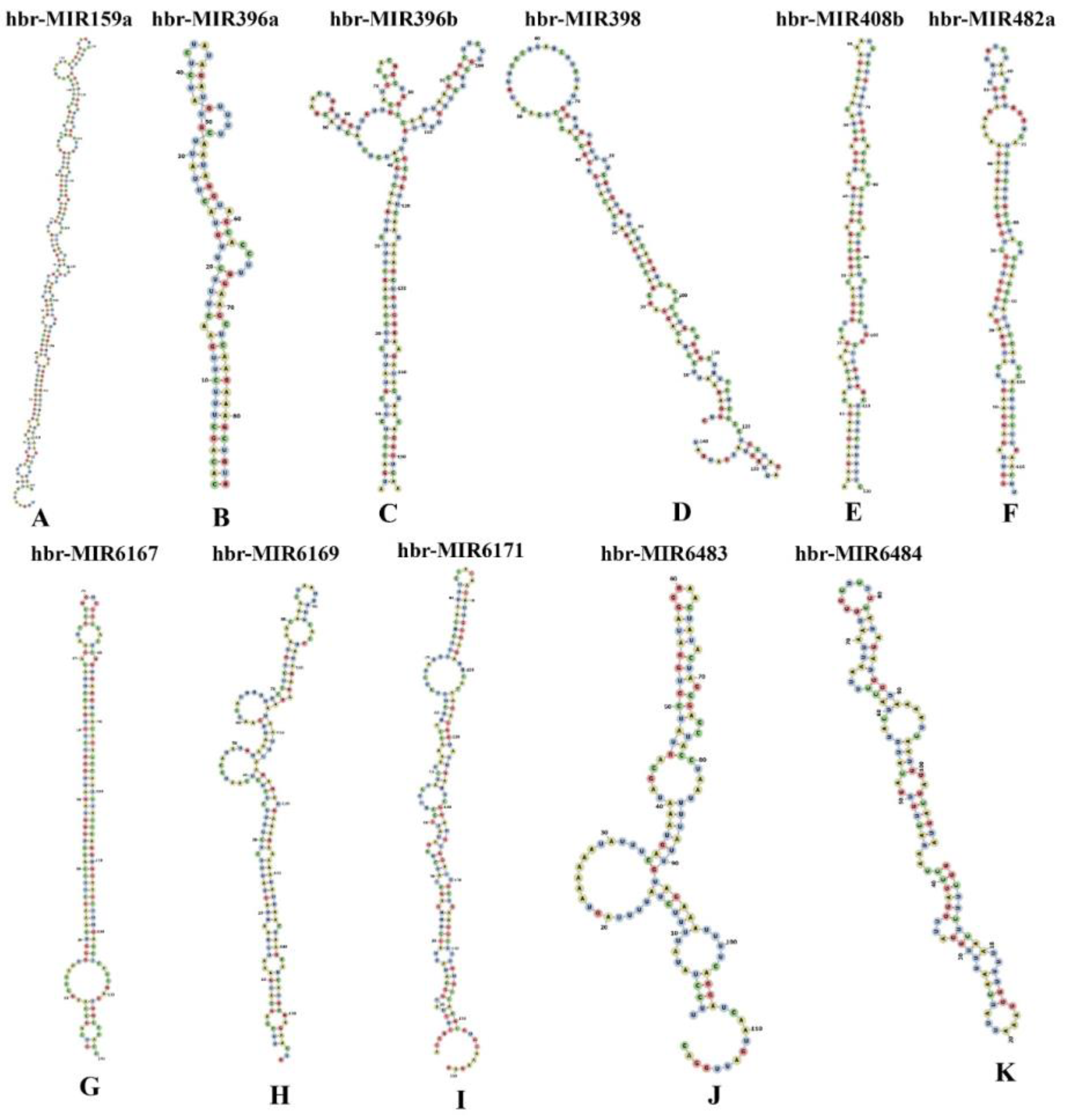
3.7. Prediction of Consensual Secondary Structures
3.8. Evaluation of Free Energy (ΔG) of mRNA-miRNA Interaction
4. Discussion
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nair, K.P. The Agronomy and Economy of Important Tree Crops of the Developing World; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Rahman, A.Y.A.; Usharraj, A.O.; Misra, B.B.; Thottathil, G.P.; Jayasekaran, K.; Feng, Y.; Hou, S.; Ong, S.Y.; Ng, F.L.; Lee, L.S. Draft genome sequence of the rubber tree Hevea brasiliensis. BMC Genom. 2013, 14, 75. [Google Scholar] [CrossRef] [Green Version]
- Go, W.Z.; Chin, K.L.; H’ng, P.S.; Wong, M.Y.; Luqman, C.A.; Surendran, A.; Tan, G.H.; Lee, C.L.; Khoo, P.S.; Kong, W.J. Virulence of Rigidoporus microporus Isolates Causing White Root Rot Disease on Rubber Trees (Hevea brasiliensis) in Malaysia. Plants 2021, 10, 2123. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, S.; Yu, F.; Li, Z.; Zhang, Y.; Wang, H.; Zhai, J. Complete Genome Sequence of a Novel Latent Tobamovirus Infecting Hevea Brasiliensis in China. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Chen, S.; Yu, F.; Li, Z.; Zhang, Y.; Wang, H.; Zhai, J.; Huang, X. Complete genome sequence of a novel virga-like virus infecting Hevea brasiliensis. Arch. Virol. 2022, 167, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, H.; Zhao, R.; Zhang, Z.; Xia, Z.; Zhai, J.; Huang, X. Complete genome sequence of a novel capillovirus infecting Hevea brasiliensis in China. Arch. Virol. 2020, 165, 249–252. [Google Scholar] [CrossRef]
- Fonseca, P.L.; Badotti, F.; De Oliveira, T.F.; Fonseca, A.; Vaz, A.; Tomé, L.M.; Abrahão, J.S.; Marques, J.T.; Trindade, G.S.; Chaverri, P. Virome analyses of Hevea brasiliensis using small RNA deep sequencing and PCR techniques reveal the presence of a potential new virus. Virol. J. 2018, 15, 184. [Google Scholar] [CrossRef]
- Akbar, S.; Wei, Y.; Zhang, M.-Q. RNA Interference: Promising Approach to Combat Plant Viruses. Int. J. Mol. Sci. 2022, 23, 5312. [Google Scholar] [CrossRef]
- Muhammad, T.; Zhang, F.; Zhang, Y.; Liang, Y. RNA interference: A natural immune system of plants to counteract biotic stressors. Cells 2019, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Khvorova, A.; Reynolds, A.; Jayasena, S.D. Functional siRNAs and miRNAs exhibit strand bias. Cell 2003, 115, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Niu, Q.-W.; Lin, S.-S.; Reyes, J.L.; Chen, K.-C.; Wu, H.-W.; Yeh, S.-D.; Chua, N.-H. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat. Biotechnol. 2006, 24, 1420–1428. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M.; Carrington, J.C. Principles of microRNA–target recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Li, Y.; Cao, X.; Qi, Y. MicroRNAs and their regulatory roles in plant–environment interactions. Annu. Rev. Plant Biol. 2019, 70, 489–525. [Google Scholar] [CrossRef] [PubMed]
- Lertpanyasampatha, M.; Gao, L.; Kongsawadworakul, P.; Viboonjun, U.; Chrestin, H.; Liu, R.; Chen, X.; Narangajavana, J. Genome-wide analysis of microRNAs in rubber tree (Hevea brasiliensis L.) using high-throughput sequencing. Planta 2012, 236, 437–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gébelin, V.; Leclercq, J.; Hu, S.; Tang, C.; Montoro, P. Regulation of MIR genes in response to abiotic stress in Hevea brasiliensis. Int. J. Mol. Sci. 2013, 14, 19587–19604. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [Green Version]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S.; Carrington, J.C. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [Green Version]
- Miranda, K.C.; Huynh, T.; Tay, Y.; Ang, Y.-S.; Tam, W.-L.; Thomson, A.M.; Lim, B.; Rigoutsos, I. A pattern-based method for the identification of MicroRNA binding sites and their corresponding heteroduplexes. Cell 2006, 126, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Bonnet, E.; He, Y.; Billiau, K.; Van de Peer, Y. TAPIR, a web server for the prediction of plant microRNA targets, including target mimics. Bioinformatics 2010, 26, 1566–1568. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhuang, Z.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Bernhart, S.H.; Tafer, H.; Mückstein, U.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. Partition function and base pairing probabilities of RNA heterodimers. Algorithms Mol. Biol. 2006, 1, 3. [Google Scholar] [CrossRef] [PubMed]
- Gandrud, C. Getting Started with R, RStudio, and knitr. In Reproducible Research with R and R Studio; Chapman and Hall, CRC Press: Boca Raton, FL, USA, 2013; pp. 53–84. [Google Scholar]
- Oliveira, A.C.; Bovolenta, L.A.; Nachtigall, P.G.; Herkenhoff, M.E.; Lemke, N.; Pinhal, D. Combining results from distinct microRNA target prediction tools enhances the performance of analyses. Front. Genet. 2017, 8, 59. [Google Scholar] [CrossRef] [Green Version]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. miRNA targets: From prediction tools to experimental validation. Methods Protoc. 2020, 4, 1. [Google Scholar] [CrossRef]
- Ghoshal, A.; Shankar, R.; Bagchi, S.; Grama, A.; Chaterji, S. MicroRNA target prediction using thermodynamic and sequence curves. BMC Genom. 2015, 16, 999. [Google Scholar] [CrossRef] [Green Version]
- Akhter, Y.; Khan, J.A. Genome wide identification of cotton (Gossypium hirsutum)-encoded microRNA targets against Cotton leaf curl Burewala virus. Gene 2018, 638, 60–65. [Google Scholar]
- Ashraf, M.A.; Feng, X.; Hu, X.; Ashraf, F.; Shen, L.; Iqbal, M.S.; Zhang, S. In Silico identification of sugarcane (Saccharum officinarum L.) genome encoded microRNAs targeting sugarcane bacilliform virus. PLoS ONE 2022, 17, e0261807. [Google Scholar] [CrossRef]
- Golyshev, V.; Pyshnyi, D.; Lomzov, A. Calculation of Energy for RNA/RNA and DNA/RNA Duplex Formation by Molecular Dynamics Simulation. Mol. Biol. 2021, 55, 927–940. [Google Scholar] [CrossRef]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for sequence-based miRNA target prediction: What to choose? Int. J. Mol. Sci. 2016, 17, 1987. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.Y.; Sieburth, L.; Voinnet, O. Widespread translational inhibition by plant miRNAs and siRNAs. Science 2008, 320, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- M Witkos, T.; Koscianska, E.; J Krzyzosiak, W. Practical aspects of microRNA target prediction. Curr. Mol. Med. 2011, 11, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Zhang, N.; Shen, W.; Li, J.-F. Engineered artificial microRNA precursors facilitate cloning and gene silencing in arabidopsis and rice. Int. J. Mol. Sci. 2019, 20, 5620. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhang, D.; Chen, S.L.; Gong, B.-Q.; Guo, Y.; Xu, L.; Zhang, X.-N.; Li, J.-F. Engineering artificial microRNAs for multiplex gene silencing and simplified transgenic screen. Plant Physiol. 2018, 178, 989–1001. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Yuan, Q.; Ai, X.; Chen, J.; Lu, Y.; Yan, F. Transgenic Rice Plants Expressing Artificial miRNA Targeting the Rice Stripe Virus MP Gene Are Highly Resistant to the Virus. Biology 2022, 11, 332. [Google Scholar] [CrossRef]
- Petchthai, U.; Yee, C.S.L.; Wong, S.-M. Resistance to CymMV and ORSV in artificial microRNA transgenic Nicotiana benthamiana plants. Sci. Rep. 2018, 8, 9958. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Hao, J.; Li, J.; Baker, B.; Luo, L. Artificial microRNA-mediated resistance to cucumber green mottle mosaic virus in Nicotiana benthamiana. Planta 2019, 250, 1591–1601. [Google Scholar] [CrossRef]
- Shahid, M.N.; Rashid, S.; Iqbal, M.S.; Jamal, A.; Khalid, S. In Silico prediction of potential mirnas to target zymv in cucumis melo. Pak. J. Bot. 2022, 54, 1319–1325. [Google Scholar]
- Jabbar, B.; Iqbal, M.S.; Batcho, A.A.; Nasir, I.A.; Rashid, B.; Husnain, T.; Henry, R.J. Target prediction of candidate miRNAs from Oryza sativa for silencing the RYMV genome. Comput. Biol. Chem. 2019, 83, 107127. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Jabbar, B.; Sharif, M.N.; Ali, Q.; Husnain, T.; Nasir, I.A. In Silico MCMV silencing concludes potential host-derived miRNAs in maize. Front. Plant Sci. 2017, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.A.; Tariq, H.K.; Khan, J.; Cheng, H. Computational Biology and Machine Learning Approaches to Identify the Rubber Tree (Hevea brasiliensis L.) Genome Encoded Potential MicroRNAs Targeting Rubber Tree Virus 1. Plant Sci. 2022, 2022070334. [Google Scholar] [CrossRef]
- Gaafar, Y.Z.A.; Ziebell, H. Novel targets for engineering Physostegia chlorotic mottle and tomato brown rugose fruit virus-resistant tomatoes: In Silico prediction of tomato microRNA targets. PeerJ 2020, 8, e10096. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, F.; Ashraf, M.A.; Hu, X.; Zhang, S. A novel computational approach to the silencing of Sugarcane Bacilliform Guadeloupe a Virus determines potential host-derived MicroRNAs in sugarcane (Saccharum officinarum L.). PeerJ 2020, 8, e8359. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Ashraf, F.; Feng, X.; Hu, X.; Shen, L.; Khan, J.; Zhang, S. Potential targets for evaluation of sugarcane yellow leaf virus resistance in sugarcane cultivars: In Silico sugarcane miRNA and target network prediction. Biotechnol. Biotechnol. Equip. 2021, 35, 1980–1991. [Google Scholar] [CrossRef]
- Zhao, T.; Wang, W.; Bai, X.; Qi, Y. Gene silencing by artificial microRNAs in Chlamydomonas. Plant J. 2009, 58, 157–164. [Google Scholar] [CrossRef]
- Schwab, R.; Ossowski, S.; Warthmann, N.; Weigel, D. Directed gene silencing with artificial microRNAs. In Plant MicroRNAs; Springer: Berlin/Heidelberg, Germany, 2010; pp. 71–88. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rubber Tree miRNAs | Locus miRanda | Locus RNA22 | Locus RNAhybrid | Locus TAPIR | Position psRNATarget | MFE * miRanda | MFE ** RNA22 | MFE * RNAhybrid | MFE-Ratio TAPIR | Expectation psRNATarget |
|---|---|---|---|---|---|---|---|---|---|---|
| hbr-miR156 | 6492 | −20.6 | ||||||||
| hbr-miR159a | 2178 | 2177 | 2177 | −20.2 | −15.2 | −24.8 | 6.5 | |||
| hbr-miR159a(1) | 3505 | 6 | ||||||||
| hbr-miR159a(2) | 2549 | 6.5 | ||||||||
| hbr-miR159a(3) | 4786 | 7 | ||||||||
| hbr-miR166a | 1060 | −25 | ||||||||
| hbr-miR166b | 6125 | −28.5 | ||||||||
| hbr-miR319 | 2992 | 569 | 5323 | 2548 | −20.2 | −18.9 | −29.8 | 7 | ||
| hbr-miR319(1) | 3204 | −16.7 | ||||||||
| hbr-miR396a | 6676 | 6675 | 6675 | 6674 | −24.7 | −21.3 | −27.2 | 0.66 | ||
| hbr-miR396b | 6678 | 823 | 6677 | −20.2 | −25.1 | 7 | ||||
| hbr-miR396b(1) | 5836 | 7 | ||||||||
| hbr-miR398 | 1839 | 1838 | 1840 | 1838 | −21.3 | −18.1 | −25.1 | 7 | ||
| hbr-miR408a | 573 | −14.2 | −25.8 | |||||||
| hbr-miR408b | 6498 | 6497 | −23.2 | −14.9 | −25.3 | |||||
| hbr-miR476 | 6176 | −21.3 | ||||||||
| hbr-miR482a | 5332 | 3951 | 5334 | 5331 | −26.1 | −17.1 | −29.3 | 5.5 | ||
| hbr-miR482a(1) | 627 | 6.5 | ||||||||
| hbr-miR482b | 6500 | −24.3 | ||||||||
| hbr-miR2118 | 5480 | 6592 | −20 | −27.8 | ||||||
| hbr-miR6166 | 1515 | −21.7 | ||||||||
| hbr-miR6167 | 5985 | 1058 | 5984 | 2716 | 1459 | −23 | −16.9 | −26.9 | 0.5 | 6.5 |
| hbr-miR6167(1) | 980 | 7 | ||||||||
| hbr-miR6168 | 5056 | 145 | 1641 | −21.1 | −19.8 | −28.6 | ||||
| hbr-miR6168(1) | 647 | −18.1 | ||||||||
| hbr-miR6169 | 3882 | 5291 | 5291 | −14.5 | −23.3 | 0.55 | 5 | |||
| hbr-miR6169(1) | 1693 | 5 | ||||||||
| hbr-miR6170 | 6660 | −20.2 | ||||||||
| hbr-miR6171 | 1633 | 2043 | 1633 | 1633 | −20.4 | −15.8 | −25.8 | 0.64 | 5.5 | |
| hbr-miR6171(1) | 6066 | −15.8 | 5.5 | |||||||
| hbr-miR6171(2) | 4540 | −15.8 | 7 | |||||||
| hbr-miR6172 | 4500 | −15.3 | −22.3 | |||||||
| hbr-miR6173 | 3749 | −24.3 | ||||||||
| hbr-miR6174 | 4303 | 5542 | −24 | 7 | ||||||
| hbr-miR6175 | 5118 | −14.9 | −25.5 | |||||||
| hbr-miR6482 | 2163 | 399 | −24.9 | 7 | ||||||
| hbr-miR6483 | 1267 | 106 | 106 | −20.1 | 0.41 | 6 | ||||
| hbr-miR6483(1) | 5666 | 7 | ||||||||
| hbr-miR6484 | 5994 | 5993 | 4295 | −20.7 | −24.7 | 6 | ||||
| hbr-miR6485 | 3042 | −22.4 | ||||||||
| hbr-miR9386 | 370 | 4422 | −24 | 6.5 | ||||||
| hbr-miR9387 |
| miRNA ID | Length of miRNA | Length of Precursor | MFE * (Kcal/mol) | AMFE ** | MFEI *** | (G + C)% | ΔG **** (Kcal/mol) |
|---|---|---|---|---|---|---|---|
| hbr-miR396a | 21 | 86 | −35.40 | −41.16 | −1.14 | 36 | −22.50 |
| hbr-miR398 | 21 | 140 | −42.80 | −30.57 | −0.61 | 50 | −18.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashraf, M.A.; Tariq, H.K.; Hu, X.-W.; Khan, J.; Zou, Z. Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1. Appl. Sci. 2022, 12, 12908. https://doi.org/10.3390/app122412908
Ashraf MA, Tariq HK, Hu X-W, Khan J, Zou Z. Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1. Applied Sciences. 2022; 12(24):12908. https://doi.org/10.3390/app122412908
Chicago/Turabian StyleAshraf, Muhammad Aleem, Hafiza Kashaf Tariq, Xiao-Wen Hu, Jallat Khan, and Zhi Zou. 2022. "Computational Biology and Machine Learning Approaches Identify Rubber Tree (Hevea brasiliensis Muell. Arg.) Genome Encoded MicroRNAs Targeting Rubber Tree Virus 1" Applied Sciences 12, no. 24: 12908. https://doi.org/10.3390/app122412908