
Integrating Different Scales into Species Distribution Models: A Case for Evaluating the Risk of Plant Invasion in Chinese Protected Areas under Climate Change

Abstract
:1. Introduction
2. Materials and Methods
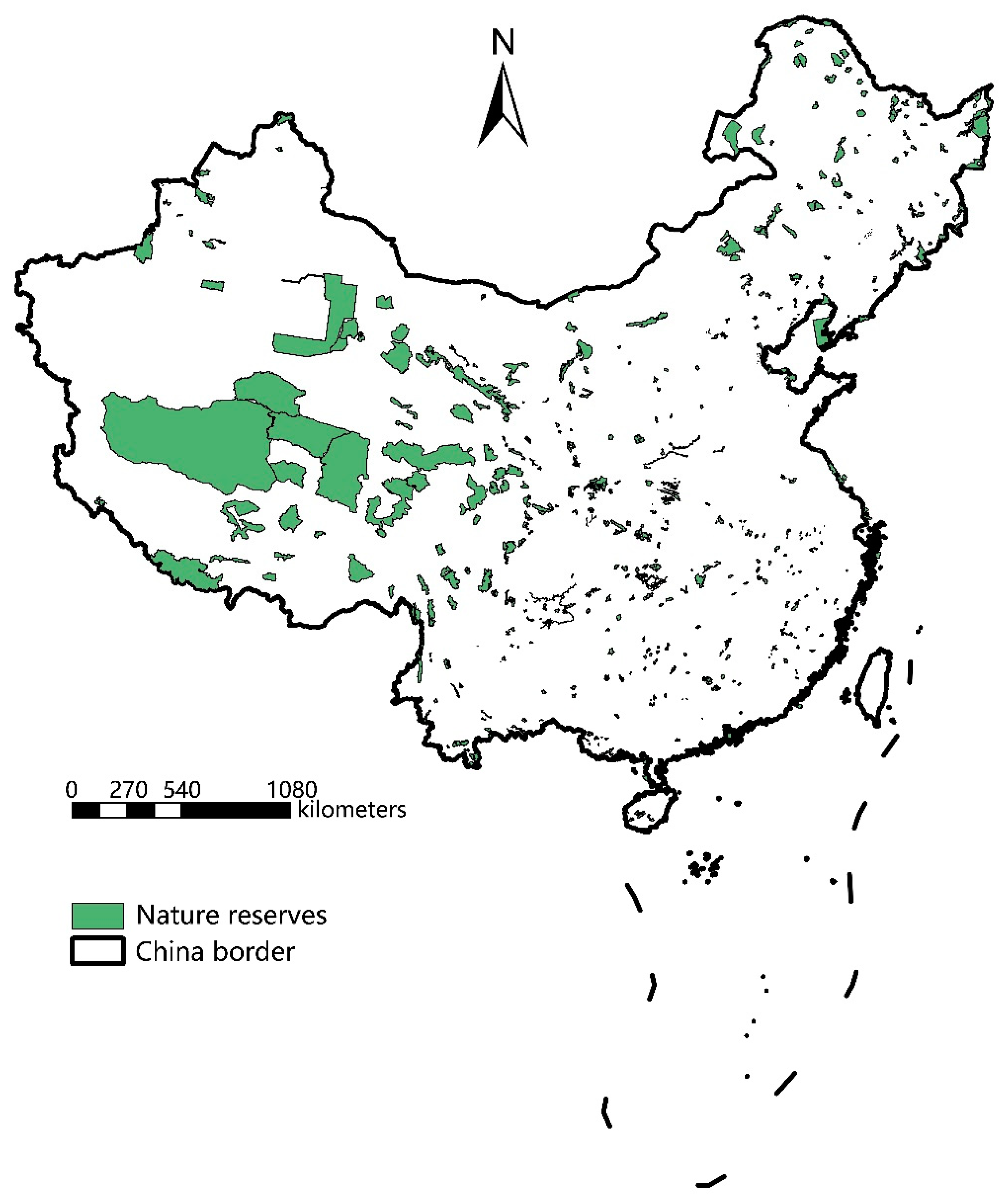
2.1. Data on Species and Protected Areas
2.2. Environmental Data
2.3. Species Distribution Models (SDMs)
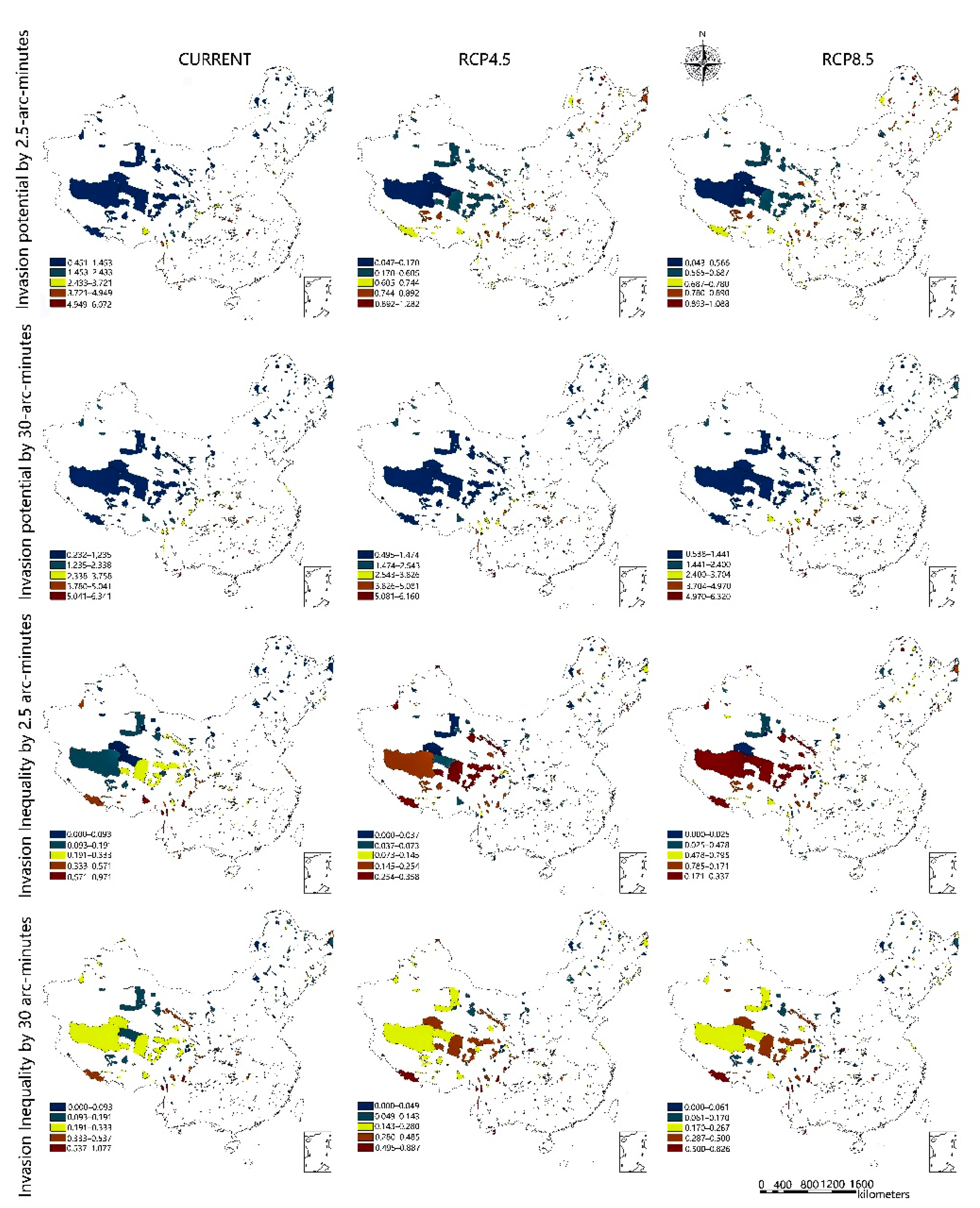
2.4. Evaluating the Risk of IAPS for Protected Areas
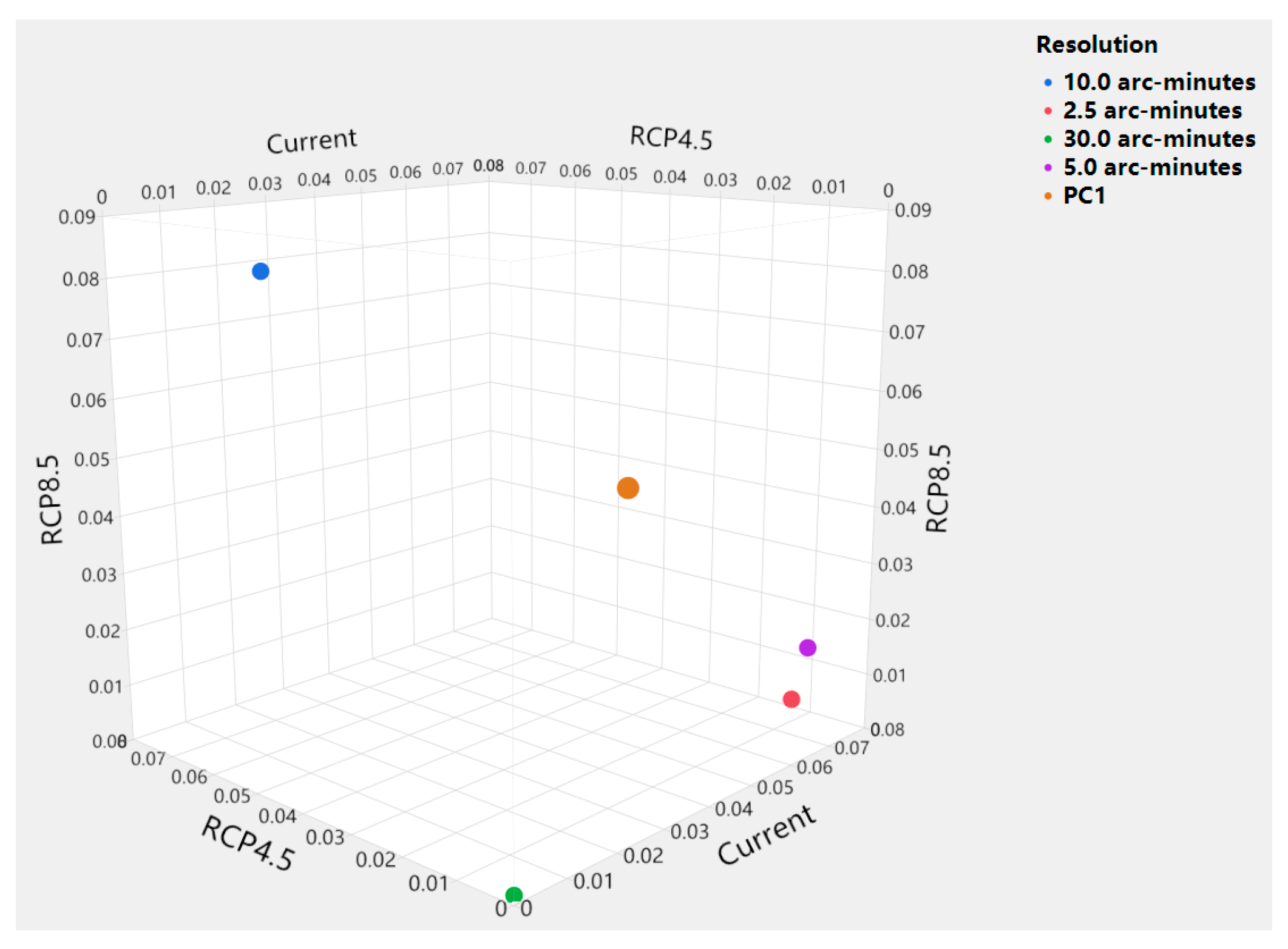
2.5. Scale-Balancing Method
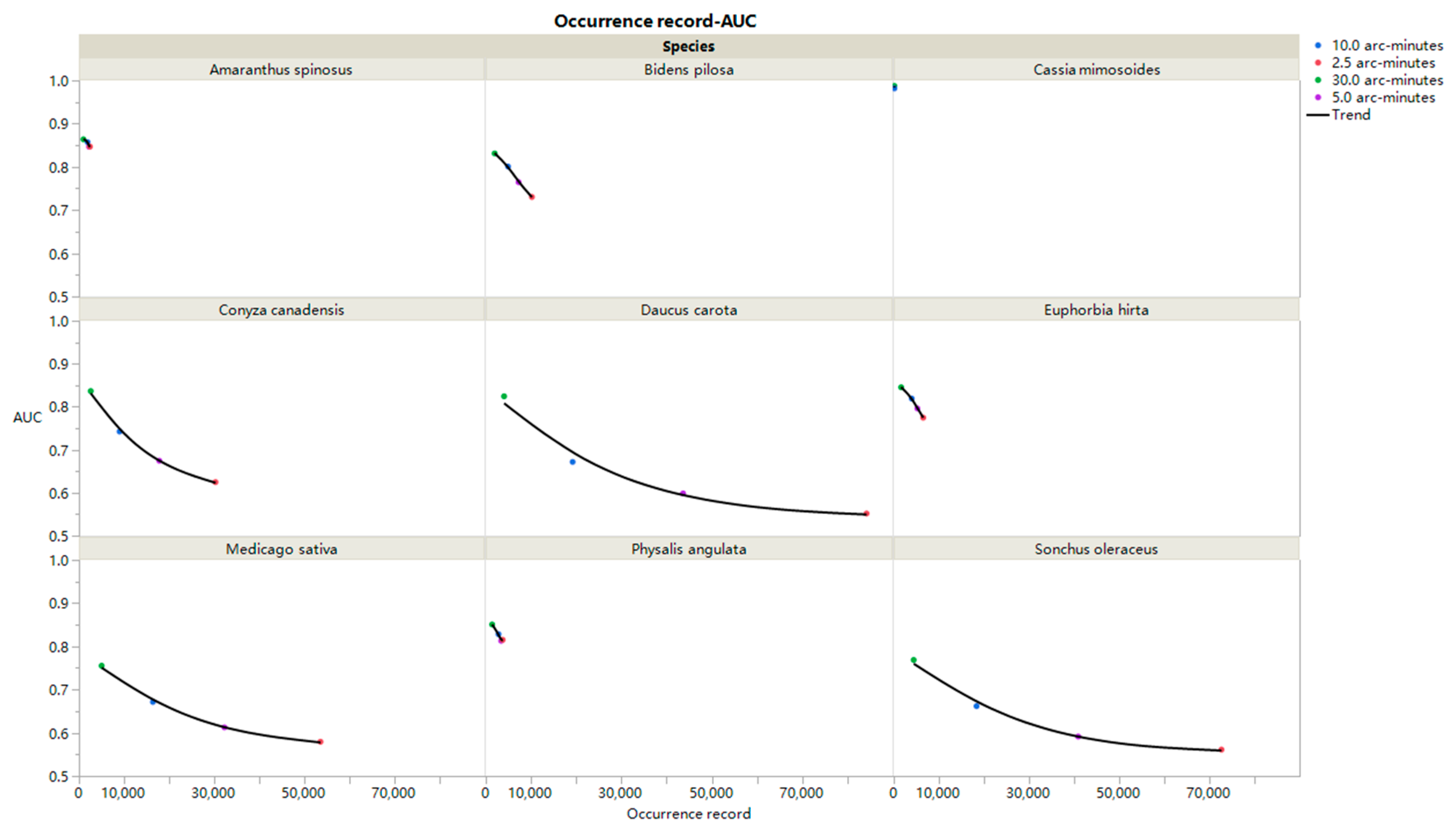
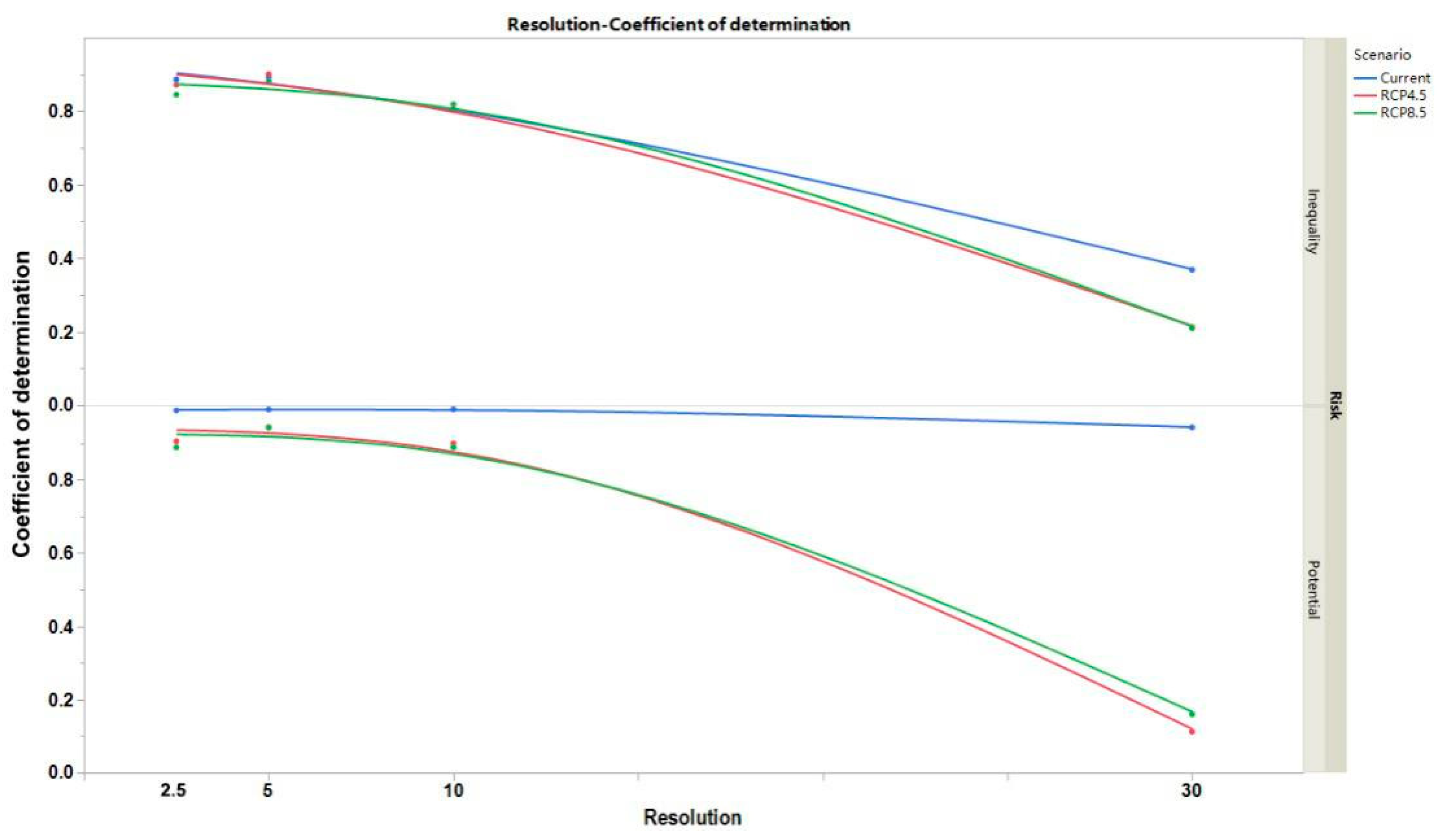
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Dubey, S.; Pike, D.A.; Shine, R. Predicting the impacts of climate change on genetic diversity in an endangered lizard species. Clim. Chang. 2014, 117, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Melton, A.E.; Soltis, D.E.; Soltis, P.S. Potential distributional shifts in North America of allelopathic invasive plant species under climate change models. Plant Divers. 2022, 44, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellard, C.; Thuiller, W.; Leroy, B.; Genovesi, P.; Bakkenes, M.; Courchamp, F. Will climate change promote future invasions? Glob. Chang. Biol. 2013, 19, 3740–3748. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Wang, R.; Yu, C.M.; Dang, X.P.; Sun, W.G.; Li, Q.F.; Wang, X.T.; Wan, J.Z. Risk assessment of insect pest expansion in alpine ecosystems under climate change. Pest Manag. Sci. 2021, 77, 3165–3178. [Google Scholar] [CrossRef]
- Zhang, F.X.; Wang, C.J.; Wan, J.Z. Using Consensus Land Cover Data to Model Global Invasive Tree Species Distributions. Plants 2022, 11, 981. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Borja, J.; David, D.; David, N. Modeling the potential area of occupancy at fine resolution may reduce uncertainty in species range estimates. Biol. Conserv. 2012, 147, 190–196. [Google Scholar]
- Pineda, E.; Lobo, J.M. The performance of range maps and species distribution models representing the geographic variation of species richness at different resolutions. Glob. Ecol. Biogeogr. 2012, 21, 935–944. [Google Scholar] [CrossRef]
- Lázaro-Lobo, A.; Ramirez-Reyes, C.; Lucardi, R.D.; Ervin, G.N. Multivariate analysis of invasive plant species distributions in southern US forests. Landsc. Ecol. 2021, 36, 3539–3555. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Yu, F.H. Effects of occurrence record number, environmental variable number, and spatial scales on MaxEnt distribution modelling for invasive plants. Biologia 2019, 74, 757–766. [Google Scholar] [CrossRef]
- Connor, T.; Hull, V.; Viña, A.; Shortridge, A.; Tang, Y.; Zhang, J.; Liu, J. Effects of grain size and niche breadth on species distribution modeling. Ecography 2018, 41, 1270–1282. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, S.A.; Griffiths, G.; Lukac, M. Species distribution model transferability and model grain size–finer may not always be better. Sci. Rep. 2018, 8, 7168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, M. Insight into distribution patterns and conservation planning in relation to woody species diversity in Xinjiang, arid northwestern China. Biol. Conserv. 2014, 177, 165–173. [Google Scholar] [CrossRef]
- Wang, Z.; Rahbek, C.; Fang, J. Effects of geographical extent on the determinants of woody plant diversity. Ecography 2012, 35, 1160–1167. [Google Scholar] [CrossRef]
- Franklin, J.; Davis, F.W.; Ikegami, M.; Syphard, A.D.; Flint, L.E.; Flint, A.L.; Hannah, L. Modeling plant species distributions under future climates: How fine scale do climate projections need to be? Glob. Chang. Biol. 2013, 19, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Bean, W.T.; Prugh, L.R.; Stafford, R.; Butterfield, H.S.; Westphal, M.; Brashares, J.S. Species distribution models of an endangered rodent offer conflicting measures of habitat quality at multiple scales. J. Appl. Ecol. 2014, 51, 1116–1125. [Google Scholar] [CrossRef]
- Suárez-Seoane, S.; Virgós, E.; Terroba, O.; Pardavila, X.; Barea-Azcón, J.M. Scaling of species distribution models across spatial resolutions and extents along a biogeographic gradient. The case of the Iberian mole Talpa occidentalis. Ecography 2014, 37, 279–292. [Google Scholar] [CrossRef]
- Metzger, M.J.; Bunce, R.G.H.; Jongman, R.H.G.; Mücher, C.A.; Watkins, J.W. A climatic stratification of the environment of Europe. Glob. Ecol. Biogeogr. 2005, 14, 549–563. [Google Scholar] [CrossRef]
- Tomas, J.B.; Amanda, E.B.; Jonathan, S.L.; Nicole, A.H.; Russell, J.T.; Graham, J.E.; Rick, D.S.; Simon, W.; Martin, K.; Jemina, F.S.; et al. Statistical solutions for error and bias in global citizen science datasets. Biol. Conserv. 2014, 173, 144–154. [Google Scholar]
- Gabriele, C.; Paolo, G.; Renato, B.; Bruno, F.; Daniele, V.; Rossella, F.; Emmanuele, F.; Simonetta, B.; Stefania, P.; Mauro, G.M. Climate change hastens the urgency of conservation for range-restricted plant species in the central-northern Mediterranean region. Biol. Conserv. 2014, 179, 129–138. [Google Scholar]
- Gallien, L.; Münkemüller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting potential distributions of invasive species: Where to go from here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Kempen, B. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Hoffman, J.D.; Aguilar-Amuchastegui, N.; Tyre, A.J. Use of simulated data from a process-based habitat model to evaluate methods for predicting species occurrence. Ecography 2010, 33, 656–666. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A.; Simberloff, D. Current mismatch between research and conservation efforts: The need to study co-occurring invasive plant species. Biol. Conserv. 2013, 160, 121–129. [Google Scholar] [CrossRef]
- Calabrese, J.M.; Certain, G.; Kraan, C.; Dormann, C.F. Stacking species distribution models and adjusting bias by linking them to macroecological models. Glob. Ecol. Biogeogr. 2014, 23, 99–112. [Google Scholar] [CrossRef]
- Rahbek, C.; Graves, G.R. Multiscale assessment of patterns of avian species richness. Proc. Natl. Acad. Sci. USA 2001, 98, 4534–4539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Václavík, T.; Kupfer, J.A.; Meentemeyer, R.K. Accounting for multi-scale spatial autocorrelation improves performance of invasive species distribution modelling (iSDM). J. Biogeogr. 2012, 39, 42–55. [Google Scholar] [CrossRef]
- Alsamadisi, A.G.; Tran, L.T.; Papeş, M. Employing inferences across scales: Integrating spatial data with different resolutions to enhance Maxent models. Ecol. Model. 2020, 415, 108857. [Google Scholar] [CrossRef]
- Schmidt, H.; Radinger, J.; Teschlade, D.; Stoll, S. The role of spatial units in modelling freshwater fish distributions: Comparing a subcatchment and river network approach using MaxEnt. Ecol. Model. 2020, 418, 108937. [Google Scholar] [CrossRef]
- Zhang, M.G.; Zhou, Z.K.; Chen, W.Y.; Cannon, C.H.; Raes, N.; Slik, J.W. Major declines of woody plant species ranges under climate change in Yunnan, China. Divers. Distrib. 2014, 20, 405–415. [Google Scholar] [CrossRef]
- Gallagher, R.V.; Hughes, L.; Leishman, M.R. Species loss and gain in communities under future climate change: Consequences for functional diversity. Ecography 2013, 36, 531–540. [Google Scholar] [CrossRef]
- Velásquez-Tibatá, J.; Salaman, P.; Graham, C.H. Effects of climate change on species distribution, community structure, and conservation of birds in protected areas in Colombia. Reg. Environ. Chang. 2013, 13, 235–248. [Google Scholar] [CrossRef]
- Vicente, J.R.; Fernandes, R.F.; Randin, C.F.; Broennimann, O.; Gonçalves, J.; Marcos, B.; Pôcas, I.; Alves, P.; Guisan, A.; Honrado, J.P. Will climate change drive alien invasive plants into areas of high protection value? An improved model-based regional assessment to prioritise the management of invasions. J. Environ. Manag. 2013, 131, 185–195. [Google Scholar] [CrossRef]
- Cotrina Sánchez, A.; Rojas Briceño, N.B.; Bandopadhyay, S.; Ghosh, S.; Torres Guzmán, C.; Oliva, M.; Salas López, R. Biogeographic Distribution of Cedrela spp. Genus in Peru Using MaxEnt Modeling: A Conservation and Restoration Approach. Diversity 2021, 13, 261. [Google Scholar] [CrossRef]
- Chen, K.; Wang, B.; Chen, C.; Zhou, G. MaxEnt Modeling to Predict the Current and Future Distribution of Pomatosace filicula under Climate Change Scenarios on the Qinghai–Tibet Plateau. Plants 2022, 11, 670. [Google Scholar] [CrossRef]
- Caplat, P.; Cheptou, P.O.; Diez, J.; Guisan, A.; Larson, B.M.H.; Macdougall, A.S.; Peltzer, D.A.; Richardson, D.M.; Shea, K.; van Kleunen, M.; et al. Movement, impacts and management of plant distributions in response to climate change: Insights from invasions. Oikos 2013, 122, 1265–1274. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, M.W.; Bradley, B.A.; Allen, J.M. Hotspots of invasive plant abundance are geographically distinct from hotspots of establishment. Biol. Invasions 2021, 23, 1249–1261. [Google Scholar] [CrossRef]
- Yu, J.; Wang, C.; Wan, J.; Han, S.; Wang, Q.; Nie, S. A model-based method to evaluate the ability of nature reserves to protect endangered tree species in the context of climate change. For. Ecol. Manag. 2014, 327, 48–54. [Google Scholar] [CrossRef]
- Wang, C.J.; Wan, J.Z.; Zhang, G.M.; Zhang, Z.X.; Zhang, J. Protected areas may not effectively support conservation of endangered forest plants under climate change. Environ. Earth Sci. 2016, 75, 46. [Google Scholar]
- Zhang, L.; Wang, S.L.; Liu, S.W.; Liu, X.J.; Zou, J.W.; Sieman, E. Perennial forb invasions alter greenhouse gas balance between ecosystem and atmosphere in an annual grassland in China. Sci. Total Environ. 2018, 642, 781–788. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Yu, F.H. Impacts of the spatial scale of climate data on the modeled distribution probabilities of invasive tree species throughout the world. Ecol. Inform. 2016, 36, 42–49. [Google Scholar] [CrossRef]
- Guisan, A.; Graham, C.H.; Elith, J.; Huettmann, F. Sensitivity of predictive species distribution models to change in grain size. Divers. Distrib. 2007, 13, 332–340. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Gueta, T.; Carmel, Y. Quantifying the value of user-level data cleaning for big data: A case study using mammal distribution models. Ecol. Inform. 2016, 34, 139–145. [Google Scholar] [CrossRef]
- Thuiller, W.; Richardson, D.M.; Pyšek, P.; Midgley, G.F.; Hughes, G.O.; Rouget, M. Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob. Chang. Biol. 2005, 11, 2234–2250. [Google Scholar] [CrossRef]
- Donaldson, J.E.; Hui, C.; Richardson, D.M.; Robertson, M.P.; NWebber, B.L.; Wilson, J.R. Invasion trajectory of alien trees: The role of introduction pathway and planting history. Glob. Chang. Biol. 2014, 20, 1527–1537. [Google Scholar] [CrossRef] [PubMed]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, D.M. Conservation biogeography: What’s hot and what’s not? Divers. Distrib. 2012, 18, 319–322. [Google Scholar] [CrossRef]
- Miller, A.J.; Knouft, J.H. GIS-based characterization of the geographic distributions of wild and cultivated populations of the Mesoamerican fruit tree Spondias purpurea (Anacardiaceae). Am. J. Bot. 2006, 93, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Wan, J.Z.; Zhang, Z.X. Expansion potential of invasive tree plants in ecoregions under climate change scenarios: An assessment of 54 species at a global scale. Scand. J. For. Res. 2017, 32, 663–670. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Yu, F.H. Risk hotspots for terrestrial plant invaders under climate change at the global scale. Environ. Earth Sci. 2016, 75, 1–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potential | Inequality | |||||||
|---|---|---|---|---|---|---|---|---|
| 2.5 | 5 | 10 | 30 | 2.5 | 5 | 10 | 30 | |
| 2.5 | 1 | 0.9987 | 0.9935 | 0.9438 | 1 | 0.949 | 0.7711 | 0.4126 |
| 5 | 0.9987 | 1 | 0.9952 | 0.9473 | 0.949 | 1 | 0.7882 | 0.4057 |
| 10 | 0.9935 | 0.9952 | 1 | 0.9536 | 0.7711 | 0.7882 | 1 | 0.4616 |
| 30 | 0.9438 | 0.9473 | 0.9536 | 1 | 0.4126 | 0.4057 | 0.4616 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, D.-J.; Zhang, F.-X.; Wang, C.-J.; Wan, J.-Z. Integrating Different Scales into Species Distribution Models: A Case for Evaluating the Risk of Plant Invasion in Chinese Protected Areas under Climate Change. Appl. Sci. 2022, 12, 11108. https://doi.org/10.3390/app122111108
Xie D-J, Zhang F-X, Wang C-J, Wan J-Z. Integrating Different Scales into Species Distribution Models: A Case for Evaluating the Risk of Plant Invasion in Chinese Protected Areas under Climate Change. Applied Sciences. 2022; 12(21):11108. https://doi.org/10.3390/app122111108
Chicago/Turabian StyleXie, De-Juan, Fei-Xue Zhang, Chun-Jing Wang, and Ji-Zhong Wan. 2022. "Integrating Different Scales into Species Distribution Models: A Case for Evaluating the Risk of Plant Invasion in Chinese Protected Areas under Climate Change" Applied Sciences 12, no. 21: 11108. https://doi.org/10.3390/app122111108