
In Silico Screening of Potential Phytocompounds from Several Herbs against SARS-CoV-2 Indian Delta Variant B.1.617.2 to Inhibit the Spike Glycoprotein Trimer

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
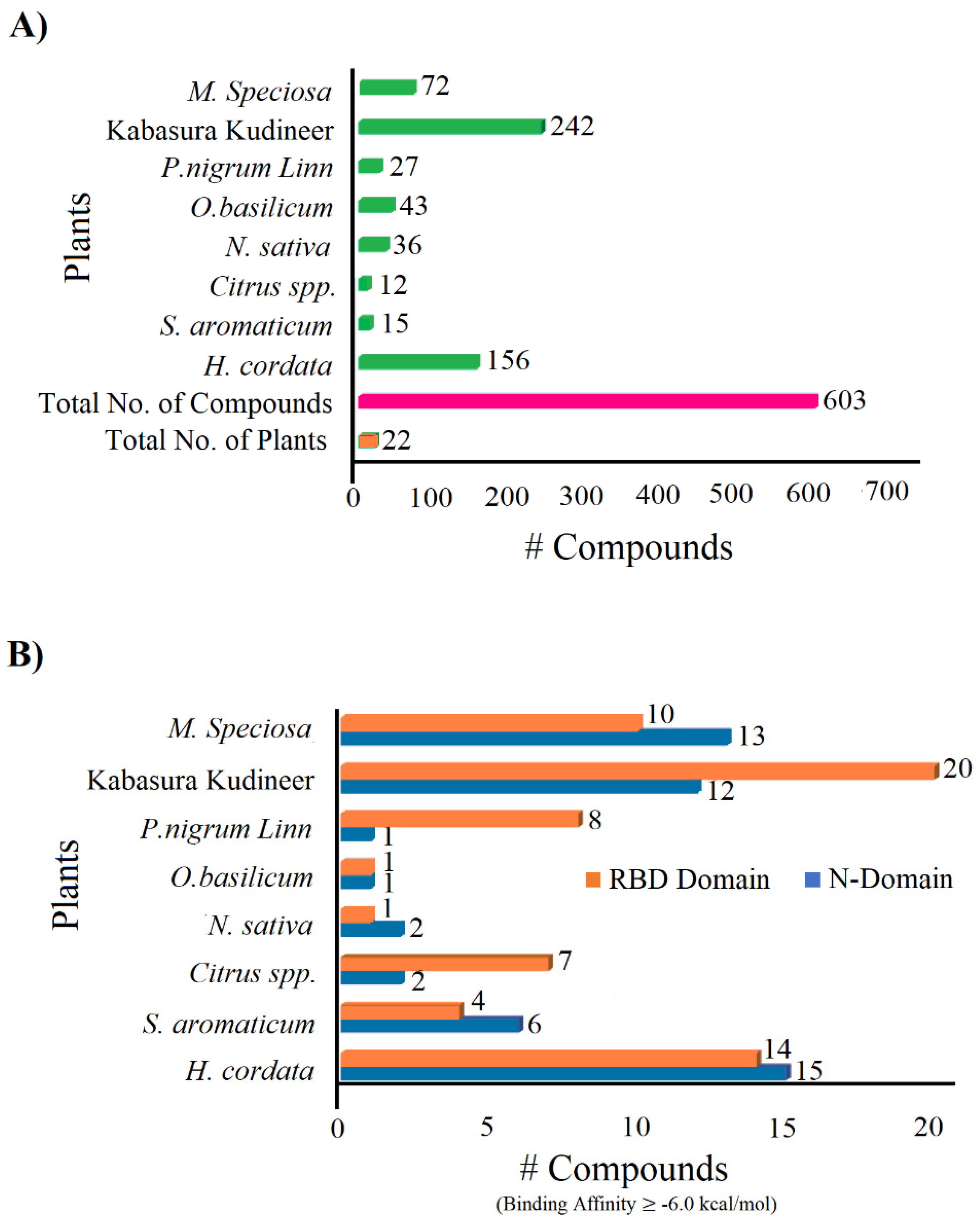
2.1. Phytocompounds from Kabasura Kudineer Choornam and Herbal Plants
2.2. Target Preparation and Ligand Library
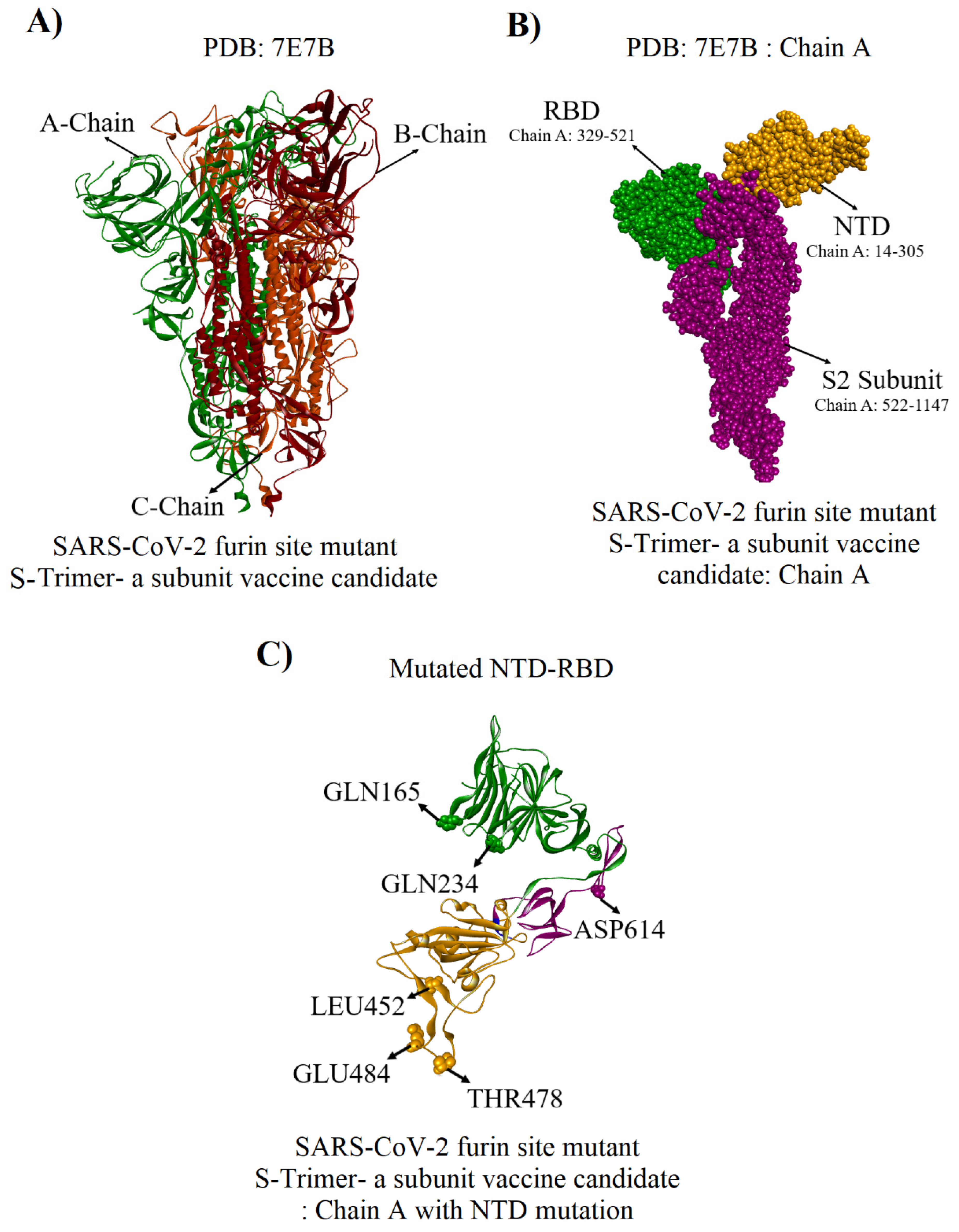
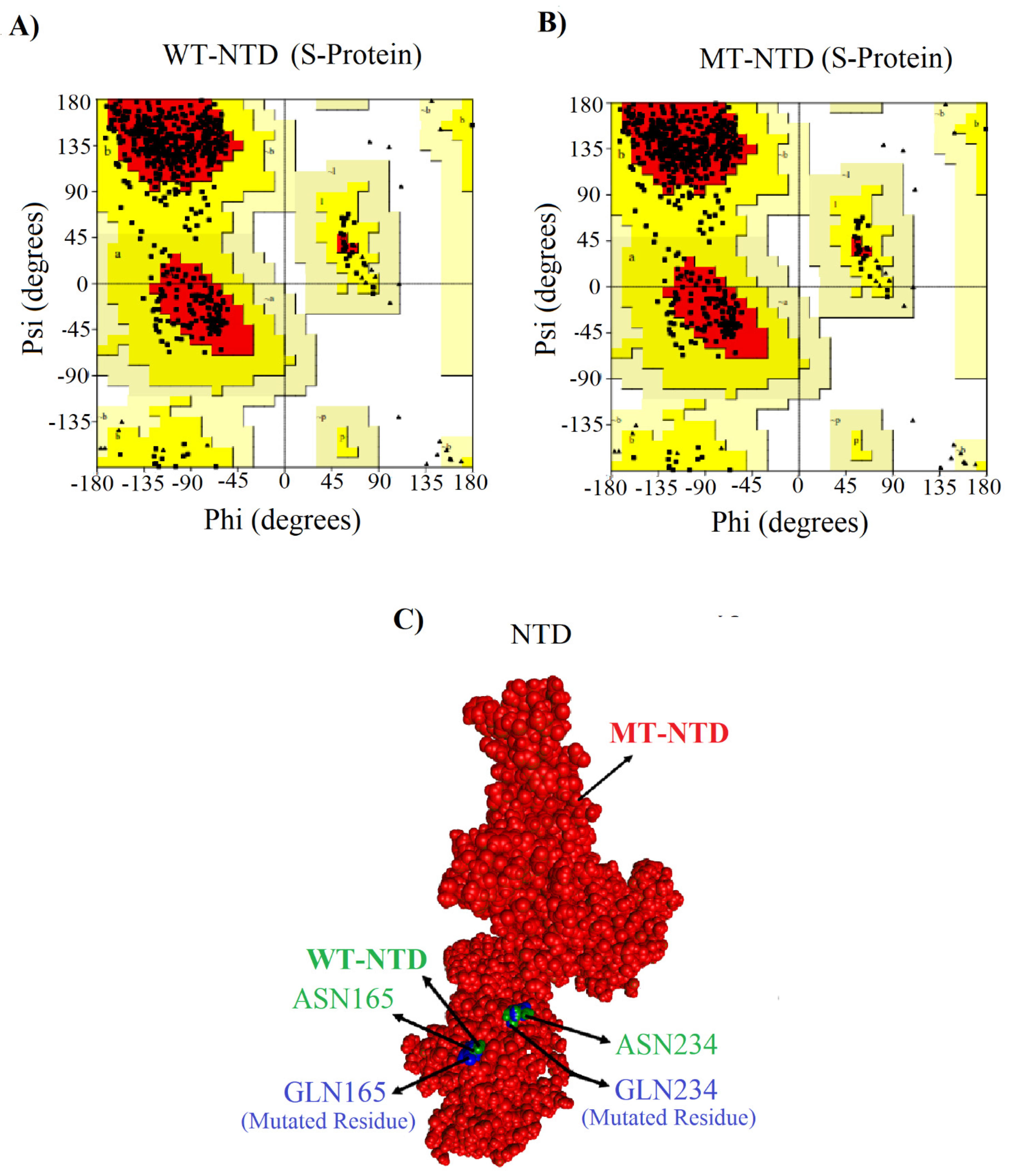
2.3. Mutated NTD Model
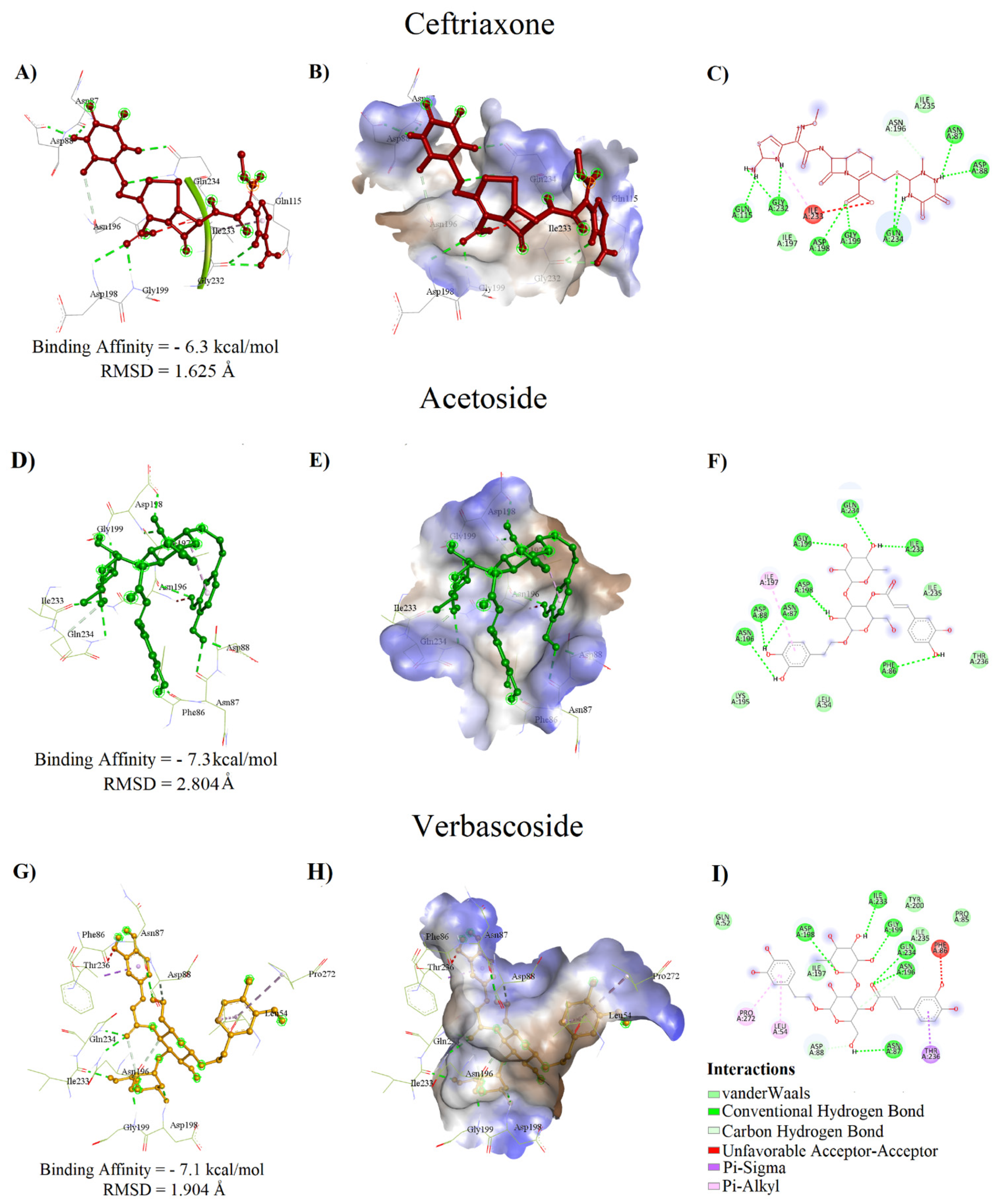
2.4. Molecular Docking
2.5. Evaluation of Ligands Drug Likeness and Toxicity
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Florindo, H.F.; Kleiner, R.; Vaskovich-Koubi, D.; Acúrcio, R.C.; Carreira, B.; Yeini, E.; Tiram, G.; Liubomirski, Y.; Satchi-Fainaro, R. Immune-mediated approaches against COVID-19. Nat. Nanotechnol. 2020, 15, 630–645. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Al-Ahmed, S.H.; Sah, R.; Tiwari, R.; Yatoo, M.I.; Patel, S.K.; Pathak, M.; Malik, Y.S.; Dhama, K.; Singh, K.P.; et al. SARS-CoV-2/COVID-19 and advances in developing potential therapeutics and vaccines to counter this emerging pandemic. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- WHO Solidarity Trial Consortium; Pan, H.; Peto, R.; Henao-Restrepo, A.M.; Preziosi, M.P.; Sathiyamoorthy, V.; Karim, A.Q.; Alejandria, M.M.; García, H.C.; Kieny, M.P.; et al. Repurposed Antiviral Drugs for COVID-19—Interim WHO Solidarity Trial Results. N. Engl. J. Med. 2021, 384, 497–511. [Google Scholar] [PubMed]
- Hu, X.; Zhou, Z.; Li, F.; Xiao, Y.; Wang, Z.; Xu, J.; Dong, F.; Zheng, H.; Yu, R. The study of anti-viral drugs targeting SARS-CoV-2 nucleocapsid and spike proteins through large-scale compound repurposing. Heliyon 2021, 7, e06387. [Google Scholar] [CrossRef] [PubMed]
- Feldman, C.; Anderson, R. The role of co-infections and secondary infections in patients with COVID-19. Pneumonia (Nathan) 2021, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Nitulescu, G.M.; Paunescu, H.; Moschos, S.A.; Petrakis, D.; Nitulescu, G.; Ion, G.N.D.; Spandidos, D.A.; Nikolouzakis, T.K.; Drakoulis, N.; Tsatsakis, A. Comprehensive analysis of drugs to treat SARS–CoV–2 infection: Mechanistic insights into current COVID–19 therapies (Review). Int. J. Mol. Med. 2020, 46, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Iacob, S.; Iacob, D.G. SARS-CoV-2 Treatment Approaches: Numerous Options, No Certainty for a Versatile Virus. Front. Pharmacol. 2020, 11, 1224. [Google Scholar] [CrossRef]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin B 2020, 10, 766–788. [Google Scholar] [CrossRef]
- Wondmkun, Y.T.; Mohammed, O.A. A Review on Novel Drug Targets and Future Directions for COVID-19 Treatment. Biologics 2020, 14, 77–82. [Google Scholar] [CrossRef]
- Faheem, B.K.; Sekhar, K.V.G.C.; Kunjiappan, S.; Jamalis, J.; Balaña-Fouce, R.; Tekwani, B.L.; Sankaranarayanan, M. Druggable targets of SARS-CoV-2 and treatment opportunities for COVID-19. Bioorg. Chem. 2020, 104, 104269. [Google Scholar] [CrossRef]
- Eastman, R.T.; Roth, J.S.; Brimacombe, K.R.; Simeonov, A.; Shen, M.; Patnaik, S.; Hall, M.D. Remdesivir: A Review of Its Discovery and Development Leading to Emergency Use Authorization for Treatment of COVID-19. ACS Cent. Sci. 2020, 6, 672–683. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Ye, S.; Zhang, Y.; Zhao, X.; Yu, Z.; Song, Y.; Tan, Z.; Tang, Y.; Chen, S.; Wang, M.; Ling, H. Emerging Variants of B.1.617 Lineage Identified Among Returning Chinese Employees Working in India—Chongqing Municipality, China, April 2021. China CDC Wkly. 2021, 3, 409–410. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Xia, H.; Zou, J.; Weaver, S.C.; Swanson, K.A.; Cai, H.; Cutler, M.; Cooper, D.; Muik, A.; et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature 2021, 596, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.J.; Fletcher, J.G.; Priya, M.T.; Veeraraghavan, B.; Mutreja, A. Relevance of immune response and vaccination strategies of SARS-CoV-2 in the phase of viral red queen dynamics. Indian J. Med. Microbiol. 2021, 39, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Pandit, P.; McArthur, A.G.; Banerjee, A.; Mossman, K. Evolutionary trajectory of SARS-CoV-2 and emerging variants. Virol. J. 2021, 18, 166. [Google Scholar]
- Singh, J.; Rahman, S.A.; Ehtesham, N.Z.; Hira, S.; Hasnain, S.E. SARS-CoV-2 variants of concern are emerging in India. Nat. Med. 2021, 27, 1131–1133. [Google Scholar] [CrossRef]
- Hasan, M.S.; Islam, M.T.; Alam, A.S.M.R.U.; Sarkar, S.L.; Rahman, M.S.; Islam, O.K.; Setu, M.A.A.; Chakrovarty, T.; Al-Emran, H.M.; Jahid, I.K.; et al. Initial reports of the SARS-CoV-2 Delta variant (B.1.617.2 lineage) in Bangladeshi patients: Risks of cross-border transmission from India. Health Sci. Rep. 2021, 4, e366. [Google Scholar] [CrossRef]
- Winger, A.; Caspari, T. The Spike of Concern-The Novel Variants of SARS-CoV-2. Viruses 2021, 13, 1002. [Google Scholar] [CrossRef]
- Mlcochova, P.; Kemp, S.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.T.M.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S.; et al. Indian SARS-CoV-2 Genomics Consortium (INSACOG); Genotype to Phenotype Japan (G2P-Japan) Consortium; CITIID-NIHR BioResource COVID-19 Collaboration. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Worldometers. 2021. Available online: https://www.worldometers.info/coronavirus/ (accessed on 9 June 2021).
- Schumaker, E. What to Kow about the Delta COVID-19 Variant First Detected in India. ABC News; 12 June 2021. Available online: https://abcnews.go.com/Health/delta-covid-19-variant-detectedindia/story?id=78197319 (accessed on 12 July 2021).
- European Centre for Disease Prevention and Control (ECDPC). SARS-CoV-2 Variants of Concern as of 7 October 2021. Available online: https://cov-lineages.org/lineage.html?lineage=B.1.617.2 (accessed on 16 December 2021).
- Mukherji, B.; Mcgregor, G. Countries Enact Travel Bans to Contain India’s COVID Variant—And Protect against ‘a Global Resurgence all over again. 2021. Available online: https://fortune.com/2021/04/28/india-travel-ban-flight-ban-covid-variant-cases-global-resurgence/ (accessed on 10 October 2021).
- Phadnis, A. India Suspends UK Flights to Prevent the Spread of New COVID-19 Strain. Business Standard News. 2021. Available online: https://www.business-standard.com/article/current-affairs/india-suspends-uk-flights-to-prevent-the-spread-of-new-covid-19-strain-120122100645_1.html (accessed on 10 October 2021).
- Sanches, P.R.S.; Charlie-Silva, I.; Braz, H.L.B.; Bittar, C.; Calmon, F.M.; Rahal, P.; Cilli, E.M. Recent advances in SARS-CoV-2 Spike protein and RBD mutations comparison between new variants Alpha (B.1.1.7, United Kingdom), Beta (B.1.351, South Africa), Gamma (P.1, Brazil) and Delta (B.1.617.2, India). J. Virus Erad. 2021, 7, 100054. [Google Scholar] [CrossRef]
- Yin, R.; Guest, J.D.; Taherzadeh, G.; Gowthaman, R.; Mittra, I.; Quackenbush, J.; Pierce, B.G. Structural and energetic profiling of SARS-CoV-2 receptor binding domain antibody recognition and the impact of circulating variants. PLoS Comput. Biol. 2021, 17, e1009380. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Rakshit, P.; Singh, S.; Abraham, P.; Panda, S.; et al. SARS-CoV-2 Spike Mutations, L452R, T478K, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. Microorganisms 2021, 9, 1542. [Google Scholar] [CrossRef]
- Cheng, M.H.; Krieger, J.M.; James, M.; Yufei, X.; Burak, K.; Yi, S.; Moshe, A.; Ivet, B. Impact of New Variants on SAR-CoV-2 Infectivity and Neutralization: A Molecular Assessment of the Alterations in the Spike-Host Protein Interactions. Lancet Preprints. Available online: https://ssrn.com/abstract=3907841 (accessed on 10 October 2021).
- WHO. Tracking SARS-CoV-2 Variants. 2021. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 10 October 2021).
- Kirola, L. Genetic emergence of B.1.617.2 in COVID-19. New Microbes New Infect. 2021, 43, 100929. [Google Scholar] [CrossRef]
- Wilhelm, A.; Toptan, T.; Pallas, C.; Wolf, T.; Goetsch, U.; Gottschalk, R.; Vehreschild, M.J.G.T.; Ciesek, S.; Widera, M. Antibody-Mediated Neutralization of Authentic SARS-CoV-2 B.1.617 Variants Harboring L452R and T478K/E484Q. Viruses 2021, 13, 1693. [Google Scholar] [CrossRef]
- Ding, C.; He, J.; Zhang, X.; Jiang, C.; Sun, Y.; Zhang, Y.; Chen, Q.; He, H.; Li, W.; Xie, J.; et al. Crucial Mutations of Spike Protein on SARS-CoV-2 Evolved to Variant Strains Escaping Neutralization of Convalescent Plasmas and RBD-Specific Monoclonal Antibodies. Front. Immunol. 2021, 12, 693775. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Garcia-Knight, M.A.; Khalid, M.M.; Servellita, V.; Wang, C.; Morris, M.K.; Sotomayor-González, A.; Glasner, D.R.; Reyes, K.R.; Gliwa, A.S.; et al. Transmission, infectivity, and neutralization of a spike L452R SARS-CoV-2 variant. Cell 2021, 184, 3426–3437.e8. [Google Scholar] [CrossRef] [PubMed]
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R.; et al. Spike mutation D614G alters SARS-CoV-2 fitness. Nature 2021, 592, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Salleh, M.Z.; Derrick, J.P.; Deris, Z.Z. Structural Evaluation of the Spike Glycoprotein Variants on SARS-CoV-2 Transmission and Immune Evasion. Int. J. Mol. Sci. 2021, 22, 7425. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2021, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.P.; Gupta, V. COVID-19 Vaccine: A comprehensive status report. Virus Res. 2020, 288, 198114. [Google Scholar] [CrossRef]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef]
- Li, Y.D.; Chi, W.Y.; Su, J.H.; Ferrall, L.; Hung, C.F.; Wu, T.C. Coronavirus vaccine development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 104. [Google Scholar] [CrossRef]
- Anand, U.; Jakhmola, S.; Indari, O.; Jha, H.C.; Chen, Z.S.; Tripathi, V.; Lastra, P.J.M. Potential Therapeutic Targets and Vaccine Development for SARS-CoV-2/COVID-19 Pandemic Management: A Review on the Recent Update. Front. Immunol. 2021, 12, 658519. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, J.; Nie, J.; Zhang, L.; Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cells 2020, 182, 1284–1294.e9. [Google Scholar] [CrossRef]
- Chung, J.Y.; Thone, M.N.; Kwon, Y.J. COVID-19 vaccines: The status and perspectives in delivery points of view. Adv. Drug Deliv. Rev. 2021, 170, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Cevik, M.; Grubaugh, N.D.; Iwasaki, A.; Openshaw, P. COVID-19 vaccines: Keeping pace with SARS-CoV-2 variants. Cell 2021, 184, 5077–5081. [Google Scholar] [CrossRef]
- Koshy, J. Coronavirus | Single Dose Only 33% Effective against B.1.617.2 Variant: U.K. 23 May 2021. The Hindu News. Available online: https://www.thehindu.com/news/national/coronavirus-single-dose-only-33-effective-against-b16172-variant-uk/article34628435.ece. (accessed on 10 October 2021).
- Bernal, L.J.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Stowe, J.; Tessier, E.; Groves, N.; Dabrera, G.; et al. Effectiveness of COVID-19 Vaccines against the B.1.617.2 (Delta) Variant. N. Engl. J. Med. 2021, 385, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Riva, L.; Yuan, S.; Yin, X.; Martin-Sancho, L.; Matsunaga, N.; Pache, L.; Muehlbacher, B.S.; Jesus, D.P.D.; Teriete, P.; Hull, M.V.; et al. Discovery of SARS-CoV-2 anti-viral drugs through large-scale compound repurposing. Nature 2020, 586, 113–119. [Google Scholar] [CrossRef]
- Bhuiyan, F.R.; Howlader, S.; Raihan, T.; Hasan, M. Plants Metabolites: Possibility of Natural Therapeutics against the COVID-19 Pandemic. Front. Med. (Lausanne) 2020, 7, 444. [Google Scholar] [CrossRef]
- Mahmood, N.; Nasir, S.B.; Hefferon, K. Plant-Based Drugs and Vaccines for COVID-19. Vaccines (Basel) 2020, 9, 15. [Google Scholar] [CrossRef]
- Khanna, K.; Kohli, S.K.; Kaur, R.; Bhardwaj, A.; Bhardwaj, V.; Ohri, P.; Sharma, A.; Ahmad, A.; Bhardwaj, R.; Ahmad, P. Herbal immune-boosters: Substantial warriors of pandemic COVID-19 battle. Phytomedicine 2021, 85, 153361. [Google Scholar] [CrossRef]
- Omrani, M.; Keshavarz, M.; Ebrahimi, N.S.; Mehrabi, M.; McGaw, L.J.; Abdalla, A.M.; Mehrbod, P. Potential Natural Products Against Respiratory Viruses: A Perspective to Develop Anti-COVID-19 Medicines. Front. Pharmacol. 2021, 11, 586993. [Google Scholar] [CrossRef]
- Mei, M.; Tan, X. Current Strategies of Antiviral Drug Discovery for COVID-19. Front. Mol. Biosci. 2021, 8, 671263. [Google Scholar] [CrossRef]
- Rehman, M.F.U.; Akhter, S.; Batool, A.I.; Selamoglu, Z.; Sevindik, M.; Eman, R.; Mustaqeem, M.; Akram, M.S.; Kanwal, F.; Lu, C.; et al. Effectiveness of Natural Antioxidants against SARS-CoV-2? Insights from the In-Silico World. Antibiotics 2021, 10, 1011. [Google Scholar] [CrossRef]
- Joshi, C.; Chaudhari, A.; Joshi, C.; Joshi, M.; Bagatharia, S. Repurposing of the herbal formulations: Molecular docking and molecular dynamics simulation studies to validate the efficacy of phytocompounds against SARS-CoV-2 proteins. J. Biomol. Struct. Dyn. 2021, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ali, G.S.; Ozdemir, B.; Selamoglu, Z. A Review of Severe Acute Respiratory Syndrome Coronavirus 2 and Pathological Disorders in Patients. J. Pharm. Care 2021, 9, 141–147. [Google Scholar] [CrossRef]
- Thas, J.J. Siddha medicine—Background and principles and the application for skin diseases. Clin. Dermatol. 2008, 26, 62–78. [Google Scholar] [CrossRef]
- Kumar, D.; Arya, V.; Kaur, R.; Bhat, Z.A.; Gupta, V.K.; Kumar, V. A review of immunomodulators in the Indian traditional health care system. J. Microbiol. Immunol. Infect. 2012, 45, 165–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunamoorthi, K.; Jegajeevanram, K.; Xavier, J.; Vijayalakshmi, J.; Melita, L. Tamil traditional medicinal system—Siddha: An indigenous health practice in the international perspectives. Tang 2012, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, S.; Anbarasi, C.; Sathiyarajeswaran, P.; Manickam, P.; Geetha, S.; Kathiravan, R.; Prathiba, P.; Pitchiahkumar, M.; Parthiban, P.; Kanakavalli, K.; et al. Kabasura Kudineer (KSK), a poly-herbal Siddha medicine, reduced SARS-CoV-2 viral load in asymptomatic COVID-19 individuals as compared to vitamin C and zinc supplementation: Findings from a prospective, exploratory, open-labeled, comparative, randomized controlled trial, Tamil Nadu, India. Trials 2021, 22, 623. [Google Scholar] [PubMed]
- Natarajan, S.; Anbarasi, C.; Sathiyarajeswaran, P.; Manickam, P.; Geetha, S.; Kathiravan, R.; Prathiba, P.; Pitchiahkumar, M.; Parthiban, P.; Kanakavalli, K.; et al. The efficacy of Siddha Medicine, Kabasura Kudineer (KSK) compared to Vitamin C & Zinc (CZ) supplementation in the management of asymptomatic COVID-19 cases: A structured summary of a study protocol for a randomised controlled trial. Trials 2020, 21, 892. [Google Scholar] [PubMed]
- Srivastava, A.; Rengaraju, M.; Srivastava, S.; Narayanan, V.; Gupta, V.; Upadhayay, R.; Kumar, J.; Parameswaran, S.; Kadarkarai, K.; Velmurugan, A. Efficacy of two siddha polyherbal decoctions, Nilavembu Kudineer and Kaba Sura Kudineer, along with standard allopathy treatment in the management of mild to moderate symptomatic COVID-19 patients-a double-blind, placebo-controlled, clinical trial. Trials 2021, 22, 570. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.; Arokiyaraj, S.; Saravanan, M.; Dhanraj, M. Molecular Docking Studies on the Anti-viral Effects of Compounds from Kabasura Kudineer on SARS-CoV-2 3CLpro. Front. Mol. Biosci. 2020, 7, 613401. [Google Scholar] [CrossRef] [PubMed]
- Nallusamy, S.; Mannu, J.; Ravikumar, C.; Angamuthu, K.; Nathan, B.; Nachimuthu, K.; Ramasamy, G.; Muthurajan, R.; Subbarayalu, M.; Neelakandan, K. Exploring Phytochemicals of Traditional Medicinal Plants Exhibiting Inhibitory Activity Against Main Protease, Spike Glycoprotein, RNA-dependent RNA Polymerase and Non-Structural Proteins of SARS-CoV-2 Through Virtual Screening. Front. Pharmacol. 2021, 12, 667704. [Google Scholar] [CrossRef]
- Lau, K.M.; Lee, K.M.; Koon, C.M.; Cheung, C.S.; Lau, C.P.; Ho, H.M.; Lee, M.Y.; Au, S.W.; Cheng, C.H.; Lau, C.B.; et al. Immunomodulatory and anti-SARS activities of Houttuynia cordata. J. Ethnopharmacol. 2008, 118, 79–85. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Siriwattananon, K.; Wangkanont, K.; Phoolcharoen, W. Perspectives on monoclonal antibody therapy as potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Asian Pac. J. Allergy Immunol. 2020, 38, 10–18. [Google Scholar]
- Wu, Z.; Deng, X.; Hu, Q.; Xiao, X.; Jiang, J.; Ma, X.; Wu, M. Houttuynia cordata Thunb: An Ethnopharmacological Review. Front. Pharmacol. 2021, 12, 714694. [Google Scholar] [CrossRef]
- Das, S.K.; Mahanta, S.; Tanti, B.; Tag, H.; Hui, P.K. Identification of phytocompounds from Houttuynia cordata Thunb. as potential inhibitors for SARS-CoV-2 replication proteins through GC-MS/LC-MS characterization, molecular docking and molecular dynamics simulation. Mol. Divers. 2021, 7, 1–24. [Google Scholar] [CrossRef]
- Bahramsoltani, R.; Rahimi, R. An Evaluation of Traditional Persian Medicine for the Management of SARS-CoV-2. Front. Pharmacol. 2020, 11, 571434. [Google Scholar] [CrossRef] [PubMed]
- Vicidomini, C.; Roviello, V.; Roviello, G.N. Molecular Basis of the Therapeutical Potential of Clove (Syzygium aromaticum L.) and Clues to Its Anti-COVID-19 Utility. Molecules 2021, 26, 1880. [Google Scholar] [CrossRef] [PubMed]
- Tallei, T.E.; Tumilaar, S.G.; Niode, N.J.; Kepel, F.B.J.; Idroes, R.; Effendi, Y.; Sakib, S.A.; Emran, T.B. Potential of Plant Bioactive Compounds as SARS-CoV-2 Main Protease (Mpro) and Spike (S) Glycoprotein Inhibitors: A Molecular Docking Study. Scientifica 2020, 2020, 6307457. [Google Scholar] [CrossRef]
- Khazdair, M.R.; Ghafari, S.; Sadeghi, M. Possible therapeutic effects of Nigella sativa and its thymoquinone on COVID-19. Pharm. Biol. 2021, 59, 696–703. [Google Scholar] [CrossRef]
- Kulyar, M.F.; Li, R.; Mehmood, K.; Waqas, M.; Li, K.; Li, J. Potential influence of Nagella sativa (Black cumin) in reinforcing immune system: A hope to decelerate the COVID-19 pandemic. Phytomedicine 2021, 85, 153277. [Google Scholar] [CrossRef] [PubMed]
- Ghoran, S.H.; El-Shazly, M.; Sekeroglu, N.; Kijjoa, A. Natural Products from Medicinal Plants with Anti-Human Coronavirus Activities. Molecules 2021, 26, 1754. [Google Scholar] [CrossRef]
- Shree, P.; Mishra, P.; Selvaraj, C.; Singh, S.K.; Chaube, R.; Garg, N.; Tripathi, Y.B. Targeting COVID-19 (SARS-CoV-2) main protease through active phytochemicals of ayurvedic medicinal plants—Withania somnifera (Ashwagandha), Tinospora cordifolia (Giloy) and Ocimum sanctum (Tulsi)—A molecular docking study. J. Biomol. Struct. Dyn. 2020, 27, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nag, A.; Chowdhury, R.R. Piperine, an alkaloid of black pepper seeds can effectively inhibit the anti-viral enzymes of Dengue and Ebola viruses, an in silico molecular docking study. Virusdisease 2020, 31, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Davella, R.; Gurrapu, S.; Mamidala, E. Phenolic compounds as promising drug candidates against COVID-19—An integrated molecular docking and dynamics simulation study. Mater. Today Proc. 2021. [Google Scholar] [CrossRef]
- Khanal, P.; Duyu, T.; Patil, B.M.; Dey, Y.N.; Pasha, I.; Wanjari, M.; Gurav, S.S.; Maity, A. Network pharmacology of AYUSH recommended immune-boosting medicinal plants against COVID-19. J. Ayurveda Integr. Med. 2020, 13, 100374. [Google Scholar] [CrossRef] [PubMed]
- Metastasio, A.; Prevete, E.; Singh, D.; Grundmann, O.; Prozialeck, W.C.; Veltri, C.; Bersani, G.; Corazza, O. Can Kratom (Mitragyna speciosa) Alleviate COVID-19 Pain? A Case Study. Front. Psychiatry 2020, 11, 594816. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [PubMed]
- Baugh, E.H.; Lyskov, S.; Weitzner, B.D.; Gray, J.J. Real-time PyMOL visualization for Rosetta and PyRosetta. PLoS ONE 2011, 6, e21931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Yin, M.; Zeng, X.; Wu, C.; Lu, A.; et al. ADMETlab 2.0: An integrated online platform for accurate and comprehensive predictions of ADMET properties. Nucleic Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A webserver for the prediction of toxicity of chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, J.; Devasia, N.; Gopalasatheeshkumar, K.; Devan, S.V.; Koikila, T.K.; Sanjay, M. Anti-inflammatory, antipyretic and antibacterial study of Kabasura kudineer choornam. Int. J. Curr. Adv. Res. 2018, 7, 9992–9997. [Google Scholar]
- Meenakumari, R.; Thangaraj, K.; Sundaram, A.; Sundaram, M.M.; Shanmugapriya, P.; Mariappan, A.; George, M.; Suba, V.; Rajalakshmi, E.; Sendhilkumar, M. Clinical outcomes among COVID-19 patients managed with modern and traditional Siddha medicine -A retrospective cohort study, Chennai, Tamil Nadu, India, 2020. J. Ayurveda Integr. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Su, D.; Sun, Y.; Huang, X.; Liang, Y.; Fang, L.; Ma, Y.; Li, W.; Liang, P.; Zheng, S. Cryo-EM structure of S-Trimer, a subunit vaccine candidate for COVID-19. J. Virol. 2021, 95, e00194-e21. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking witha new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed] [Green Version]
- Hirschhorn, H.H. Botanical remedies of the former Dutch East Indies (Indonesia): Part I: Eumycetes, pteridophyta, gymnospermae, angiospermae (monocotyledones only). J. Ethnopharm. 1983, 7, 123–156. [Google Scholar] [CrossRef]
- Ren, X.; Sui, X.; Yin, J. The effect of Houttuynia cordata injection on pseudorabies herpes virus (PrV) infection in vitro. Pharma. Biol. 2011, 49, 161–166. [Google Scholar] [CrossRef]
- Leardkamolkarn, V.; Sirigulpanit, W.; Phurimsak, C.; Kumkate, S.; Himakoun, L.; Sripanidkulchai, B. The inhibitory actions of Houttuynia cordata aqueous extract on dengue virus and dengue infected cells. J. Food Biochem. 2012, 36, 86–92. [Google Scholar] [CrossRef]
- Jiangang, F.; Ling, D.; Zhang, L.; Hongmei, L. Houttuynia cordata Thunb: A review of phytochemistry and pharmacology and quality control. Chin. Med. 2013, 4, 101–123. [Google Scholar]
- Hung, P.Y.; Ho, B.C.; Lee, S.Y.; Chang, S.Y.; Kao, C.L.; Lee, S.S.; Lee, C.N. Houttuynia cordata targets the beginning stage of herpes simplex virus infection. PLoS ONE 2015, 10, e0115475. [Google Scholar] [CrossRef] [Green Version]
- Satthakarn, S.; Chung, W.O.; Promsong, A.; Nittayananta, W. Houttuynia cordata modulates oral innate immune mediators: Potential role of herbal plant on oral health. Oral Dis. 2015, 21, 512–518. [Google Scholar] [CrossRef]
- Sit, N.W.; Chan, Y.S.; Chuah, B.L.; Cheng, R.J.; Leong, W.M.; Khoo, K.S. Anti-viral, antifungal and antibacterial activities of the Chinese medicinal plants, Houttuynia cordata, Lobelia chinensis and Selaginella uncinata. Southeast Asian J. Trop. Med. Public Health 2018, 48, 616–627. [Google Scholar]
- Cheng, D.; Sun, L.; Zou, S.; Chen, J.; Mao, H.; Zhang, Y. Anti-viral effects of Houttuynia cordata polysaccharide extract on Murine Norovirus-1 (MNV-1): A human norovirus surrogate. Molecules 2019, 24, 1835. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.J.; Lu, Y.; Zhang, Y.Y.; Zhu, H.Y.; Tu, P.; Li, H.; Chen, D.F. Flavonoids from Houttuynia cordata attenuate H1N1-induced acute lung injury in mice via inhibition of influenza virus and Toll-like receptor signalling. Phytomedicine 2020, 67, 153150. [Google Scholar] [CrossRef]
- Fawe, A.; Zaid, M.A.; Menzies, J.G.; Bélanger, R.R. Silicon-Mediated Accumulation of flavonoid Phytoalexins in Cucumber. J. Phytopathol. 1998, 88, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Panda, C.K.; Das, S. Clove (Syzygium aromaticum L.), a potential chemopreventive agent for lung cancer. Carcinogenesis 2006, 27, 1645–1654. [Google Scholar] [CrossRef] [Green Version]
- Horiuchi, K.; Shiota, S.; Hatano, T.; Yoshida, T.; Kuroda, T.; Tsuchiva, T. Antimicrobial activity of oleanolic acid from Salvia officinalis and related compounds on vancomycin-resistant enterococci (VRE). J. Biol. Pharm. Bull. 2007, 30, 1147–1149. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.H.; Chen, W.; Zhao, Y.; Ju, X.L. Antitumor activity of Oleanolic, Ursolic and Glycyrrhetinic acid. Open Nat. Prod. J. 2009, 2, 48–52. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, M.N.I.; Begum, J.; Nandi, N.C.; Akter, F. Constituents of the essential oil from leaves and buds of clove (Syzygium caryophyllatum (L.) Alston). Afr. J. Plant. Sci. 2010, 4, 451–454. [Google Scholar]
- Vasconellos, M.C.; Benzerra, D.P.; Fonseca, A.M.; Pessoa, C.; Lemos, T.L.; Lotufo, C.L.V.; de Moraes, M.O.; Montenegro, R.C. The in vitro and in vivo inhibitory activity of biflorin in melanoma. J. Melanoma Res. 2011, 21, 106–114. [Google Scholar] [CrossRef]
- Sánchez-Tena, S.; Reyes-Zurita, F.J.; Díaz-Moralli, S.; Vinardell, M.P.; Reed, M.; García-García, F.; Dopazo, J.; Lupiáñez, J.A.; Günther, U.; Cascante, M. Maslinic acid-enriched diet decreases intestinal tumorigenesis in Apc (Min/+) mice through transcriptomic and metabolomic reprogramming. PLoS ONE 2013, 8, e59392. [Google Scholar]
- Castellano, J.M.; Guinda, A.; Deloado, T.; Rada, M.; Cavuela, J.A. Biochemical basis of the antidiabetic activity of oleanolic acid and related pentacyclic triterpenes. J. Diabetes 2013, 62, 1791–1799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Schmitz, J.C.; Wei, J.; Cao, S.; Beumer, J.H.; Strychor, S.; Cheng, L.; Liu, M.; Wang, C.; Wu, N.; et al. Clove extract inhibits tumor growth and promotes cell cycle arrest and apoptosis. Oncol. Res. 2014, 21, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Jnawali, H.N.; Lee, E.; Jeong, K.W.; Shin, A.; Heo, Y.S.; Kim, Y. Anti-inflammatory Activity of Rhamnetin and a Model of Its Binding to c-Jun NH-Terminal Kinase 1 and p38 MAPK. J. Nat. Prod. 2014, 77, 258–263. [Google Scholar] [CrossRef]
- Park, E.S.; Kang, J.C.; Jang, Y.C.; Park, J.S.; Jang, S.Y.; Kim, D.E.; Kim, B.; Shin, H.S. Cardioprotective effects of rhamnetin in H9c2 cardiomyoblast cells under H2O2-induced apoptosis. J. Ethnopharmacol. 2014, 153, 552–560. [Google Scholar] [CrossRef]
- Mossadeq, S.W.M.; Sulaiman, M.R.; Mohamad, T.T.A.; Chiong, H.S.; Zakaria, Z.A.; Jabit, M.L.; Israf, D.A. Anti-inflammatory and antinociceptive effects of Mitragyna speciosa Korth methanolic extract. Med. Princ. Pract. 2009, 18, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, S.; Azizi, J.; Ramanathan, B.S.; Ismail, S.; Sasidharan, S.; Mohd, M.I.; Mansor, S.M. Evaluation of antioxidant and antibacterial activities of aqueous, methanolic and alkaloid extracts from Mitragyna speciosa (rubiaceae family) leaves. Molecules 2009, 14, 3964–3974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizi, J.; Ismail, S.; Mordi, M.N.; Ramanathan, S.; Said, M.I.M.; Mansor, S.M. In vitro and in vivo effects of three different mitragyna speciosa korth leaf extracts on phase II drug metabolizing enzymes-glutathione transferases (GSTs). Molecules 2010, 15, 432–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, T.B.; Yian, K.R.; Mordi, M.N.; Mansor, S.M. Antioxidant value and antiproliferative efficacy of mitragynine and a silane reduced analougue. Asian Pac. J. Cancer Prev. 2014, 15, 5659–5665. [Google Scholar] [CrossRef] [Green Version]
- Srichana, K.; Janchawee, B.; Prutipanlai, S.; Raungrut, P.; Keawpradub, N. Effects of mitragynine and a crude alkaloid extract derived from Mitragyna speciosa korth. On permethrin elimination in rats. Pharmaceutics 2015, 7, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Goh, Y.S.; Karunakaran, T.; Murugaiyah, V.; Santhanam, R.; Abu Bakar, M.H.; Ramanathan, S. Accelerated Solvent Extractions (ASE) of Mitragyna speciosa Korth. (Kratom) Leaves: Evaluation of Its Cytotoxicity and Antinociceptive Activity. Molecules 2021, 26, 3704. [Google Scholar] [CrossRef]
- Singh, M.K.; Khare, G.; Iyer, S.K.; Sharwan, G.; Tripathi, D.K. Clerodendrum serratum: A clinical approach. J. App. Pharm. Sci. 2012, 2, 11–15. [Google Scholar]
- Patel, J.J.; Acharya, S.R.; Acharya, N.S. Clerodendrum serratum (L.) Moon.—A review on traditional uses, phytochemistry and pharmacological activities. Ethnopharmacology 2014, 154, 268–285. [Google Scholar] [CrossRef]
- Kshirsagar, A.D.; Ingale, K.G.; Vyawahare, N.S.; Thorve, V.S. Hygrophila spinosa: A comprehensive review. Pharmacogn. Rev. 2010, 4, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Salve, S.D.; Bhuktar, A.S. Pharmacognosy and phytochemical evaluation of Hygrophila auriculata (Schumach.) heine root. J. Phytopharm. 2017, 6, 210–216. [Google Scholar] [CrossRef]
- Sodhi, T. Ayurveda in Veterinary Medicine. In Proceedings of the AHVMA Annual Conference, Durham, NC, USA, 20–23 September 2003. [Google Scholar]
- Kirtikar, K.R.; Basu, B.D. Indian Medicinal Plants, 1st ed.; Bio-green Books: Delhi, India, 1987; pp. 757–759. [Google Scholar]
- Shobana, S.; Naidu, A.K. Antioxidant activity of selected Indian spices. Prostaglandins Leukot. Essent. Fat. Acids. 2000, 62, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Waisundara, V.Y.; Watawana, M.I.; Jayawardena, N. Costus speciosus and Coccinia grandis: Traditional medicinal remedies for diabetes. S. Afr. J. Bot. 2015, 98, 1–5. [Google Scholar] [CrossRef]
- Selim, S.; Jaouni, S.A. Anticancer and apoptotic effects on cell proliferation of diosgenin isolated from Costus speciosus (Koen.) Sm. BMC Complementary Altern. Med. 2015, 15, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Far, A.H.; Shaheen, H.M.; Alsenosy, A.W.; El-Sayed, Y.S.; Al Jaouni, S.K.; Mousa, S.A. Costus speciosus: Traditional uses, phytochemistry, and therapeutic potentials. Pharmacogn. Rev. 2018, 12, 120–127. [Google Scholar] [CrossRef]
- Ma, S.C.; Du, J.; But, P.P.; Deng, X.L.; Zhang, Y.W.; Ooi, V.E.; Xu, H.X.; Lee, S.H.; Lee, S.F. Antiviral Chinese medicinal herbs against respiratory syncytial virus. J. Ethnopharmacol. 2002, 79, 205–211. [Google Scholar] [CrossRef]
- Jadhav, A.K.; Karuppayil, S.M. Andrographis paniculata (Burm. F) Wall ex Nees: Anti-viral properties. Phytother. Res. 2021, 127, 5365–5373. [Google Scholar] [CrossRef]
- Aglin, A.A. Medicinal effects of Mollugo cerviana—A Review. Int. J. Sci. Res. Multidiscip. Stud. 2018, 4, 34–37. [Google Scholar]
- Napagoda, M.; Gerstmeier, J.; Butschek, H.; De Soyza, S.; Pace, S.; Lorenz, S.; Qader, M.; Witharana, S.; Nagahawatte, A.; Wijayaratne, G.; et al. The Anti-Inflammatory and Antimicrobial Potential of Selected Ethnomedicinal Plants from Sri Lanka. Molecules 2020, 25, 1894. [Google Scholar] [CrossRef]
- Reddy, B.S.; Rao, N.R.; Vijeepallam, K.; Pandy, V. Phytochemical, pharmacological and biological profiles of Tragia species (Family: Euphorbiaceae). Afr. J. Tradit. Complement Altern. Med. 2017, 14, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, H.; Wang, H. Glycans of SARS-CoV-2 Spike Protein in Virus Infection and Antibody Production. Front. Mol. Biosci. 2021, 8, 629873. [Google Scholar] [CrossRef] [PubMed]
- Baral, P.; Bhattarai, N.; Hossen, M.L.; Stebliankin, V.; Gerstman, B.S.; Narasimhan, G.; Chapagain, P.P. Mutation-induced changes in the receptor-binding interface of the SARS-CoV-2 Delta variant B.1.617.2 and implications for immune evasion. Biochem. Biophys. Res. Commun. 2021, 574, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Motozono, C.; Toyoda, M.; Zahradnik, J.; Saito, A.; Nasser, H.; Tan, T.S.; Ngare, I.; Kimura, I.; Uriu, K.; Kosugi, Y.; et al. SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microbe 2021, 29, 1124–1136.e11. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokova, V.; Kulakesara, H.; Larson, L.; Bowers, V.; Rechkina, E.; Kisiela, D.; Sledneva, Y.; Choudhury, D.; Maslova, I.; Deng, K.; et al. Acquisition of the L452R mutation in the ACE2-binding interface of Spike protein triggers recent massive expansion of SARS-CoV-2 variants. J. Clin. Microbiol. 2021, 11, JCM0092121. [Google Scholar] [CrossRef]
- Mouffouk, C.; Mouffouk, S.; Mouffouk, S.; Hambaba, L.; Haba, H. Flavonols as potential anti-viral drugs targeting SARS-CoV-2 proteases (3CLpro and PLpro), spike protein, RNA-dependent RNA polymerase (RdRp) and angiotensin-converting enzyme II receptor (ACE2). Eur. J. Pharmacol. 2021, 891, 173759. [Google Scholar] [CrossRef]
- Rosales-Mendoza, S.; Márquez-Escobar, V.A.; González-Ortega, O.; Nieto-Gómez, R.; Arévalo-Villalobos, J.I. What Does Plant-Based Vaccine Technology Offer to the Fight against COVID-19? Vaccines 2020, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, Z.; Waterhouse, P.; Bally, J. Plant-Based Vaccines: The Way Ahead? Viruses 2020, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Pandita, D.; Jain, G.K.; Agarwal, P.; Grewal, A.S.; Khar, R.K.; Lather, V. Role of phytoconstituents in the management of COVID-19. Chem. Biol. Interact. 2021, 341, 109449. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Witek-Krowiak, A.; Skrzypczak, D.; Mikula, K.; Młynarz, P. Phytochemicals containing biologically active polyphenols as an effective agent against COVID-19-inducing coronavirus. J. Funct. Foods. 2020, 73, 104146. [Google Scholar] [CrossRef]
- Anand, A.V.; Balamuralikrishnan, B.; Kaviya, M.; Bharathi, K.; Parithathvi, A.; Arun, M.; Senthilkumar, N.; Velayuthaprabhu, S.; Saradhadevi, M.; Al-Dhabi, N.A.; et al. Medicinal Plants, Phytochemicals, and Herbs to Combat Viral Pathogens Including SARS-CoV-2. Molecules 2021, 26, 1775. [Google Scholar] [CrossRef]
- RECOVERY Collaborative Group. Lopinavir-ritonavir in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2020, 396, 1345–1352. [Google Scholar] [CrossRef]
- Jorgensen, S.C.J.; Kebriaei, R.; Dresser, L.D. Remdesivir: Review of Pharmacology, Pre-clinical Data, and Emerging Clinical Experience for COVID-19. Pharmacotherapy 2020, 40, 659–671. [Google Scholar] [CrossRef]
- Touret, F.; de Lamballerie, X. Of chloroquine and COVID-19. Antivir. Res. 2020, 177, 104762. [Google Scholar] [CrossRef]
- Nojomi, M.; Yassin, Z.; Keyvani, H.; Makiani, M.J.; Roham, M.; Laali, A.; Dehghan, N.; Navaei, M.; Ranjbar, M. Effect of Arbidol (Umifenovir) on COVID-19: A randomized controlled trial. BMC Infect. Dis. 2020, 20, 954. [Google Scholar] [CrossRef]
- Joshi, S.; Parkar, J.; Ansari, A.; Vora, A.; Talwar, D.; Tiwaskar, M.; Patil, S.; Barkate, H. Role of favipiravir in the treatment of COVID-19. Int. J. Infect. Dis. 2021, 102, 501–508. [Google Scholar] [CrossRef]
- Hung, I.F.; Lung, K.C.; Tso, E.Y.; Liu, R.; Chung, T.W.; Chu, M.Y.; Ng, Y.Y.; Lo, J.; Chan, J.; Tam, A.R.; et al. Triple combination of interferon beta-1b, lopinavir-ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID-19: An open-label, randomised, phase 2 trial. Lancet 2020, 395, 1695–1704. [Google Scholar] [CrossRef]
- Boulware, D.R.; Pullen, M.F.; Bangdiwala, A.S.; Pastick, K.A.; Lofgren, S.M.; Okafor, E.C.; Skipper, C.P.; Nascene, A.A.; Nicol, M.R.; Abassi, M.; et al. A Randomized Trial of Hydroxychloroquine as Postexposure Prophylaxis for COVID-19. N. Engl J. Med. 2020, 383, 517–525. [Google Scholar] [CrossRef]
- Nourian, A.; Khalili, H. Sofosbuvir as a potential option for the treatment of COVID-19. Acta Biomed. 2020, 91, 236–238. [Google Scholar]
- Kallingal, A.; Thachan Kundil, V.; Ayyolath, A.; Karlapudi, A.P.; Muringayil Joseph, T.; E, J.V. Molecular modeling study of tectoquinone and acteoside from Tectona grandis linn: A new SARS-CoV-2 main protease inhibitor against COVID-19. J. Biomol. Struct. Dyn. 2020, 9, 1–12. [Google Scholar] [CrossRef]
- Shawky, E.; Nada, A.A.; Ibrahim, R.S. Potential role of medicinal plants and their constituents in the mitigation of SARS-CoV-2: Identifying related therapeutic targets using network pharmacology and molecular docking analyses. RSC Adv. 2020, 10, 27961–27983. [Google Scholar] [CrossRef]
- Mbaveng, A.T.; Ngameni, B.; Kuete, V.; Simo, I.K.; Ambassa, P.; Roy, R.; Bezabih, M.; Etoa, F.X.; Ngadjui, B.T.; Abegaz, B.M.; et al. Antimicrobial activity of the crude extracts and five flavonoids from the twigs of Dorstenia barteri (Moraceae). J. Ethnopharmacol. 2008, 116, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benet, L.Z.; Hosey, C.M.; Ursu, O.; Oprea, T.I. BDDCS, the Rule of 5 and drugability. Adv. Drug Deliv. Rev. 2016, 101, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidelines for Ayurveda Practitioners for COVID 19. Available online: https://vikaspedia.in/health/ayush/guidelines-for-ayush-practitioners-for-covid-19/guidelines-for-ayurveda-practitioners-for-covid19 (accessed on 11 November 2021).
- World Health Organization. 2020. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update–-19-january-2021 (accessed on 11 November 2021).
- Gautam, S.; Gautam, A.; Chhetri, S.; Bhattarai, U. Immunity against COVID-19: Potential Role of Ayush Kwath. J. Ayurveda Integr. Med. 2020, 100350. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Phytocompounds | Binding Affinity | RMSD (Å) | H/C-H Bond Interaction | Bond Length | Hydrophobic Interaction | Alkyl Interaction | Pi-Sigma/Cation Stacked Interaction |
|---|---|---|---|---|---|---|---|---|
| B.1.617.2. S-Protein—N-Domain (Mutant Type) | ||||||||
| Standard Drug | Ceftriaxone | −6.3 | 1.625 | ASP88, ASN87, GLN115, ASN156* ASP198, GLY199, GLY232, GLN234*, | 2.57, 2.23, 2.10, 2.54, 2.03, 2.76, 2.28. 2.93 | ASN196, ILE197 | ILE233 | - |
| H. cordata | Cholest-4,14-dien-15,20-diol-3,16-dione | −6 | 2.414 | ASN196, GLN234 | 5.03, 4.72 | ASP88, ASP198, GLY199, GLY232, ILE233, ILE235 | LEU54, ILE197 | - |
| Dihydrocelastrol | −6.6 | 1.41 | ASP53, ASN196, GLN234, ILE235 | 4.56, 3.63, 4.74, 4.38 | ASN87, ASP88, LYS195, ILE233 | LEU54, ILE197 | ||
| Isoquercitrin | −6.4 | 2.445 | ASN196, ILE233, GLN234, ILE235, ASN87* | 4.40, 5.36, 4.45, 5.41 | LEU54, PHE86, ILE197, ASP198, GLY199, THR236 | ASP88 | ||
| Naltrindole | −6.3 | 2.726 | ASN87, ILE197, ASP198, ILE233, ILE235, THR236 | GLN234, ASN1196, LEU54, PRO272 | ASP88 | |||
| Pirenperone | −8.7 | 1.395 | ILE233 | - | ASN87, ASP88, ASN196, GLT199, GLY232, GLN234, PRO272 | ILE197 | LEU54 | |
| Quercitrin | −6.3 | 1.588 | ASN87, ASP88, ASN196, GLN234, ILE235 | 4.54, 3.67, 4.74, 4.47, 5.69, 3.46 | LEU54, PHE86, ASP198, GLY199, ILE233 | - | - | |
| Rhodoxanthin | −6.8 | 2.258 | ASP198 | - | THR114, GLN115, GLU132, GLN165, CYS166, THR167, ASN196, ILE197, GLY199, GLY232, ILE233, GLN234 | - | - | |
| Sesamin | −9.1 | 0.046 | ASN196*, ILE197*, ASP198 | - | ASP88, ILE233, GLN234, ILE235, PRO272 | LEU54 | ||
| Usambarensine | −6.8 | 1.9 | ASN196, ASP198*, ILE233* | 4.57 | ASP88, GLY199, GLN234, ILE235 | LEU54 | ASP197, ASP198 | |
| S. aromaticum | Biflorin | −6 | 1.436 | ASP88, ASN196, GLN234, ILE235 GLN234* | PHE86, ILE197, ASP198, GLY199, TYR200, ILE233, THR236 | GLN234 | - | |
| Crategolic acid | −6.4 | 1.764 | ASN196, GLN234 | 3.96, 4.57 | LEU54, ASP88, ILE197, ASP198, GLY199, ILE235, PRO272 | |||
| Oleanolic acid | −6.2 | 2.01 | ASN87*, ILE235 | ASP53, LEU54, PHE86, ASP88, ASN196, ILE197, GLN234, THR236 | - | - | ||
| Rhamnetin | −6 | 1.785 | ASN87, ASP88, GLY199 | 3.29, 6.01, 3.40 | PHE86, ASN196, ASP198, GLY232, ILE233, GLN234, ILE235 | |||
| M. speciosa Korthi | Chlorogenic acid | −6.1 | 2.307 | PHE86, ASN196, ILE235 | 4.34, 2.65, 5.55 | LEU54, ASP88, ASN87, ILE197, ASP198, GLY199, TYR200, GLY232, ILE233, GLN234, THR236 | ||
| Isoquercitrin | −6.4 | 2.484 | ASN196, ILE233, GLN234, ILE235, ASN87* | 6.26*, 4.25, 4.43, 5.59, 4.47 | LEU54, PHE86, ILE197, ASP198, GLY199, THR236 | - | ASP88 | |
| Rutin | −6.1 | 2.149 | ASN196, ASN196*, ASP198, GLN234*, ILE235 | 3.79, 4.05*, 3.85, 3.67*, 5.01 | PHE86, ASN87, ASP88, GLY199, GLY232, ILE233, THR236 | LEU54, ILE197 | - | |
| Kabasura kudineer | ||||||||
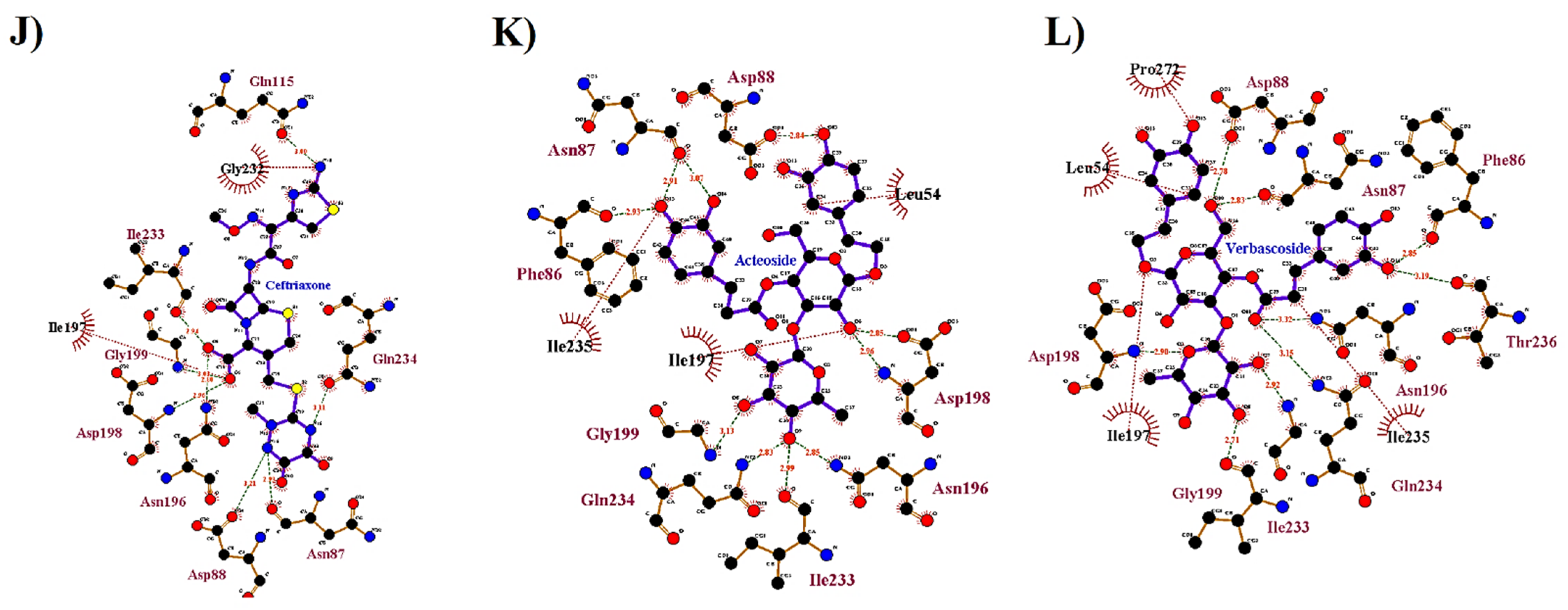
| C. serratum | Acteoside | −7.3 | 2.804 | PHE86, ASN87, ASP88, ASN196, ASP198, GLY199, ILE233, GLN234 | 4.45, 6.04, 3.09, 4.34, 3.47, 5.84, 3.78 | LEU54, LYS195, THR236 | ILE197 | |
| Serratagenic acid | −6.8 | 2.404 | ASP88, GLN234 | 4.33, 4.54 | - | - | - | |
| Verbascoside | −7.1 | 1.904 | ASN87, ASP88*, ASN196, ASP198, GLY199, ILE233, GLN234 | 6.07, 3.99, 3.49, 4.04, 5.37, 4.73 | GLN52, PRO85, ILE235 | LEU54, PRO272 | THR236 | |
| H. auriculata | Apigenin 7-O-glucoside | −6.5 | 2.588 | ASN87, ASN196, GLY199, GLN234, ILE235 | 5.13, 3.54, 3.65, 3.42, 3.95, 4.52 | PHE86, ASP88, ILE233 | ASP198 | |
| Cucurbitacin B | −6.2 | 2.838 | GLN234, GLY199 | 3.34, 4.84 | PHE86, ASN87, ASP88, THR108, THR114, ILE197, ASP198, ILE233, ILE235, THR236 | - | - | |
| A. paniculata | Neoandrographolide | −6.3 | 1.466 | ASN87, ASN196, GLN234, ILE235, PRO272* | 3.79, 4.2, 5.13, 4.25, 5.55 | GLN52, ASP53, LEU54, PHE86, THR236 | ASP88 | |
| M. cerviana | Orientin | −6.2 | 1.189 | ASN87*, ASP88, ASN196, ILE233, GLN234, ILE235 | 4.32, 421, 5.42, 4.98, 3.95 | PHE86, ASN87, ASP198, GLY199, GLY232, THR236 | ILE197 | - |
| Vitexin | −6.2 | 1.562 | ASN87, ASP88, ASN196, GLN234, GLN234*, ILE235, ASP198 | 5.44, 3.01, 3.24, 3.52, 4.21 | - | - | - | |
| T. involucrate L. | Rutin | −6.4 | 1.702 | ASP88*, ASN196, ASP198, GLY199, ILE233, GLN234 | 4.39, 2.93, 4.48, 4.79 | PHE86, ASN87, GLY89, ILE231, GLY232, ILE235, PRO272 | LEU54, ILE197 | |
| T. cordifolia | Tinosporide | −6.1 | 2.551 | ASN87*, ASP198, GLY199, ILE233. ILE235* | 4.50, 3.59, 5.90 | PHE86, ASP88, ASN196, TYR200, GLN234 | THR236 | |
| Plant | Phytocompounds | Binding Affinity | RMSD | H/C-H Bond Interaction | Bond Length | Hydrophobic Interaction | Alkyl Interaction | Pi-Sigma/Cation Stacked |
|---|---|---|---|---|---|---|---|---|
| (Å) | Interaction | |||||||
| B.1.617.2. S-Protein—RBD-Domain | ||||||||
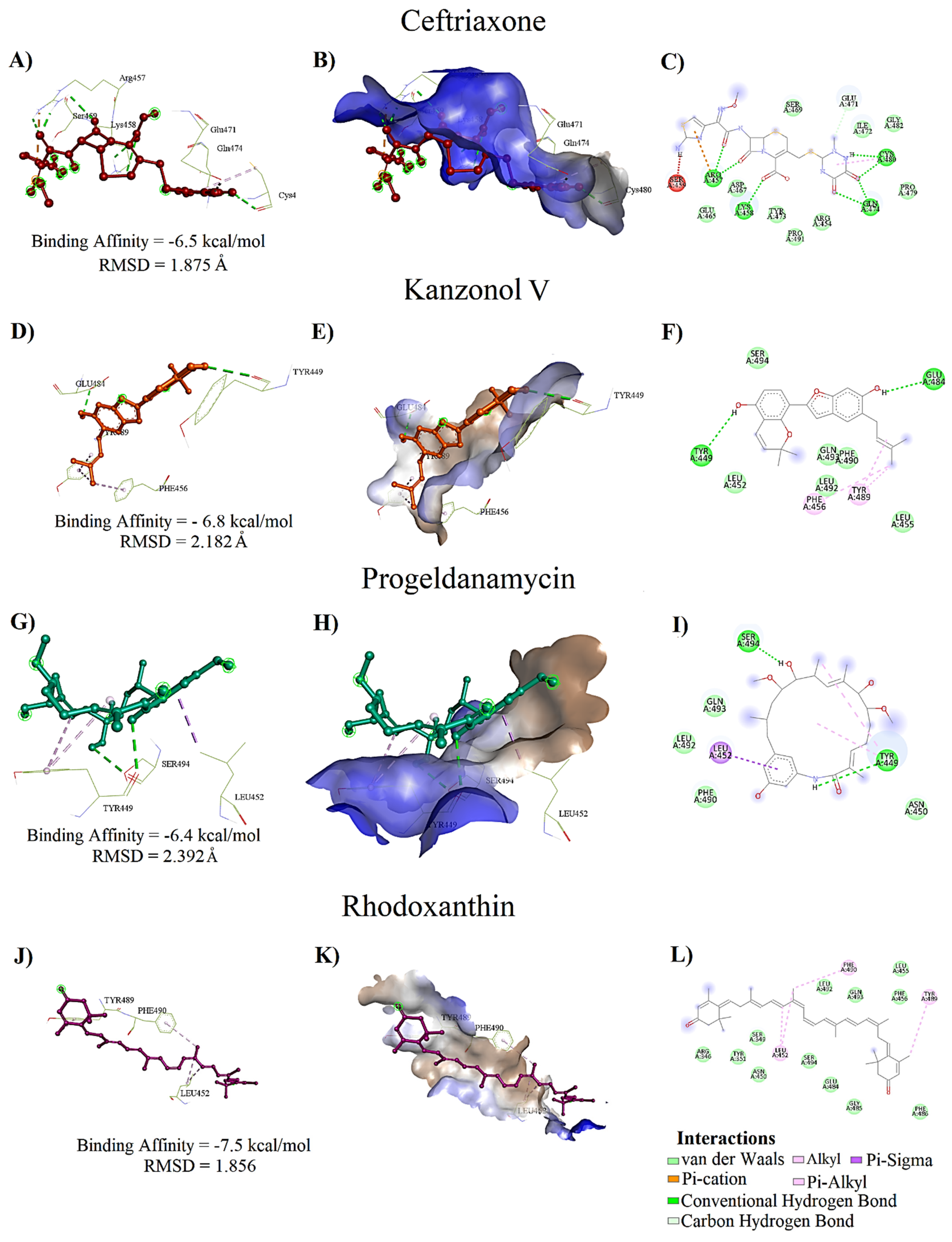
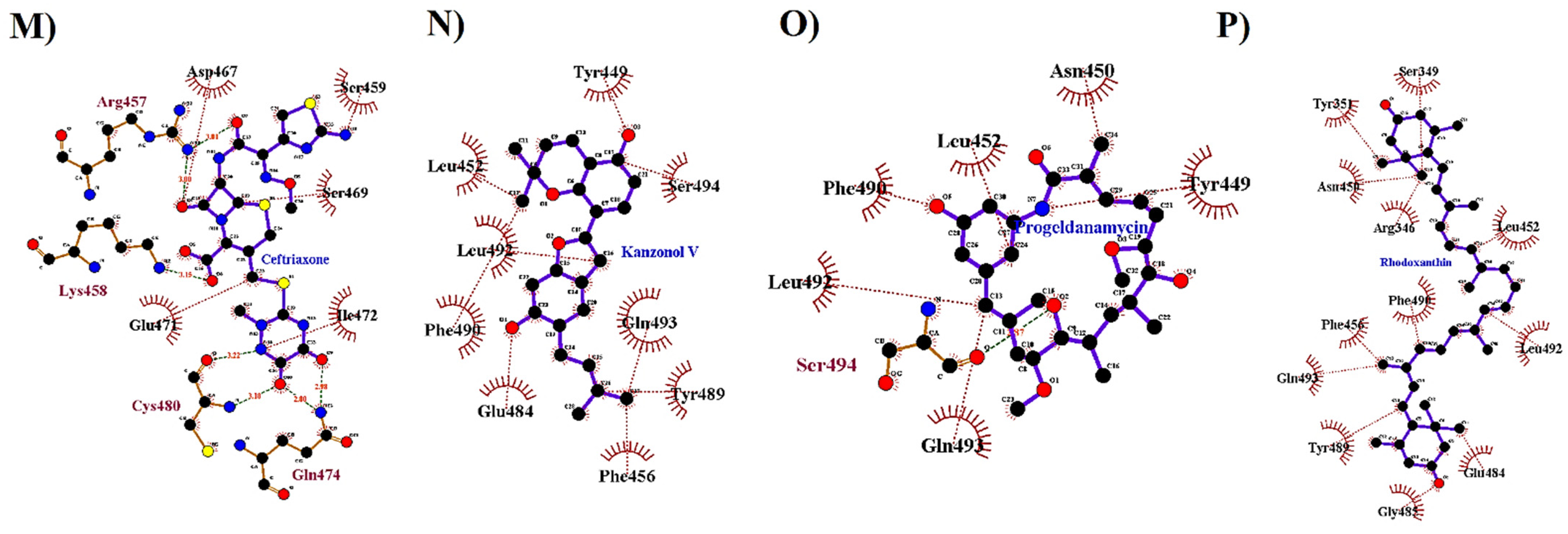
| Standard Drug | Ceftriaxone | −6.5 | 1.625 | ARG457, LYS458, GLU471*, GLN474, CYS480 | 3.45, 5.86; 3.40, 4.14, 3.52, 3.59, 3.60 | ARG454, GLU465, ASP567, SER469, TYR473, PRO479, GLY482, PRO491 | CYS480 | ARG457 |
| H. cordata | Canthaxanthin | −7.2 | 1.543 | - | - | ARG403, TYR449, GLU484, GLN493, SER494, TYR495, GLY496, THR500, ASN501, GLY502, TYR505 | - | PHE490 |
| Cholest-4,14-dien-15,20-diol-3,16-dione | −6 | 2.764 | SER494 | 4.08 | LEU452, GLU484, GLY485, CYS488, TYR489, LEU492, GLN493 | - | PHE490 | |
| Fluorometholone 17-acetate | −6.1 | 3.672 | PHE490, GLN493 | 4.54, 5.00 | LEU452, PHE456, GLU484, TYR489, SER494 | - | LEU492 | |
| Kanzonol V | −6.8 | 2.182 | GLU484, TYR449 | 4.54, 4.55 | LEU452, LEU455, PHE490, LEU492, GLN493, SER494 | - | PHE456, TYR489 | |
| Progeldanamycin | −6.4 | 2.392 | TYR449, SER494 | 3.45, 2.62 | ASN450, PHE490, LEU492, GLN493 | - | LEU452 | |
| Rhodoxanthin | −7.5 | 1.856 | - | - | ARG346, SER349, TYR351, ASN450, LEU455, PHE456, GLU484, GLY485, PHE486, LEU492, GLN493, SER494 | LEU452, TYR489, PHE490 | - | |
| Stigmastane-3,6-dione, (5.alpha) | −6.8 | 3.213 | GLN493, SER494 | 4.98, 4.04 | TYR351, LEU452, THR470, GLU484, LEU492 | PHE490 | - | |
| S. aromaticum | Rhamnetin | −6.1 | 2.981 | ARG346, SER349, TRP353, SER349 | 4.12, 3.78, 4.73 | PHE347, ARG355, LEU452, ARG466 | TYR351, ALA352 | ALA348 |
| M. speciosa Korthi | Beta-Sitosterol | −6 | 1.586 | GLY485 | 3.45 | GLU484, PHE486, ASN487, LEU492, GLN493, SER494 | LEU452, TYR489, PHE490 | - |
| Stigmasterol | −6 | 3.456 | - | - | LEU455, GLN484, LEU492, GLN493 | LEU452, PHE456, TYR489, | PHE490 | |
| PHE490 | ||||||||
| Kabasura kudineer | ||||||||
| T. cordifolia | Berberine | −6.1 | 2.725 | PHE490, GLN493, SER494 | 4.73, 4.32, 4.15 | TYR449, TYR489, LEU492 | LEU452 | - |
| C. serratum | Clerodermic acid | −5.8 | 4.924 | GLU484 | 3.64, 4.37 | TYR449, LEU452, THR470, LEU492, GLN493, SER494 | - | PHE490 |
| C. speciosus | Diosgenin | −6.9 | 1.058 | SER349 | 4.28 | ARG346, PHE347, ALA348, ASN450, TRP353, ASN354, ARG355, ARG466, ILE468 | ALA352, LEU452 | - |
| Plant Name | Active Compounds | Plants Parts | Pharmacological Properties | References |
|---|---|---|---|---|
| H. cordata | Canthaxanthin, Cholest-4,14-dien-15,20-diol-3,16-dione, Dihydrocelastrol, Fluorometholone 17-acetate, Isoquercitrin, Kanzonol V, Naltrindole, Pirenperone, Progeldanamycin, Quercitrin, Rhodoxanthin, Sesamin, Stigmastane-3,6-dione, (5.alpha), Usambarensine | Whole Plant | Cough, pneumonia, bronchitis, dysentery, dropsy, leukorrhea, uteritis, eczema, herpes simplex, acne, chronic sinusitis, stomach ulcer, infection, control wrinkle, chapped skin, septic, febrifuge, heatstroke, malaria, lung disorder, tonsillitis, skin ulcer, diarrhea, dysentery arthritis, appendicitis, snake bite, stomach disorder, sinusitis, heart disorders, severe acute respiratory Syndrome (SARS), chikungunya, herpes simplex viruses, dengue virus serotype 2 (DEN-2), infuenza neuraminidase, pseudorabies herpes virus (prv), human noroviruses (hunovs), murine coronavirus and dengue virus infection, innate immune modulation activities, and inhibits the replication of SARS-CoV. | [65,90,91,92,93,94,95,96,97,98] |
| S. aromaticum | Biflorin, Crategolic acid, Oleanolic acid, Rhamnetin | Cloves buds (Oil) | Coughs, colds, asthma, respiratory and digestive disorders, sinusitis, modulatory effects of cell membrane permeability, acts against food borne gram-positive bacteria, promotion of Go/G1 cell cycle arrest, induction of apoptosis, anti-diabetic activity, antioxidant, antitumor, cardio protective, antifungal, and acts effectively against SARS-CoV-2. | [70,99,100,101,102,103,104,105,106,107,108,109] |
| M. speciosa Korthi | Chlorogenic acid, Isoquercitrin, Rutin, Beta-Sitosterol, Stigmasterol | Leaves | Tiredness and muscle fatigue, diarrhea, coughing, muscle pain, anti-diabetic, wound, hypertension, drug addiction, anti-inflammation, antinociceptive, anti-oxidant, antimicrobial activity, and reduction of muscle pain against SARS-CoV-2. | [79,110,111,112,113,114,115] |
| Kabasura kudineer | ||||
| C. serratum | Acteoside, Clerodermic acid, Serratagenic acid, Verbascoside | Root | Respiratory disease, fever, anti-inflammatory, anticancer, antinociceptive, liver disorders, anti-allergic, and acts as anti-oxidant. | [63,64,116,117] |
| H. auriculata | Apigenin 7-O-glucoside | Root | Anasarca, urinogenital tract disorder, hyperdipsia, vesical calculi, flatulence, diarrhea, leukorrhea, gonorrhea, gastrointestinal disorder, anti-tumor, arthritis, painful micturition, menorrhagia, and treats blood infection. | [63,64,118,119] |
| T. cordifolia | Berberine, Tinosporide | Stem | Immuno-modulation, pneumonia, asthma, cough, swelling lungs, colic, constipation, tetanus, anthrax, pox, fracture, antispasmodic, and antipyretic activity. | [63,64,75,120,121] |
| C. speciosus | Diosgenin | Root | Pneumonia, constipation, skin diseases, fever, asthma, bronchitis, inflammation, anaemia, dropsy, cough, urinary diseases, jaundice, improves insulin secretion, hypolipidemic, adaptogenic, anticancer, and hepatoprotective activity. | [63,64,122,123,124,125] |
| A. paniculata | Neoandrographolide | Whole plant | Colds, sinusitis, influenza, immunostimulant, anti-viral against hepatitis B, HIV, and respiratory syncytial virus. | [63,64,126,127] |
| M. cerviana | Orientin, Vitexin | Whole plant | Anti-inflammatory, anti-oxidant, antimicrobial, antidiabetic, hepatoprotective, photo-protective, uterine stimulant, antiseptic, antipyretic, and immunostimulant activity. | [63,64,128,129] |
| T. involucrata | Rutin | Root | High fever, inflammation, wounds, eczema, scabies, skin infections, bronchitis pain, and antimicrobial activity. | [63,64,121,130] |
| Phytocompounds | Drug-Likeness | Toxicity Analysis | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Name | Compound Name | miLogP | TPSA | Natoms | nON | nOHNH | No. of Violations | Intestinal Absorption | Oral Toxicity (−log kg/mol) | Hepato Toxicity | Carcino Genicity | Immuno Toxicity | Muta Genicity | Cyto Toxicity | LD50 (mg/kg) | TC |
| B.1.617.2. S-Protein—NTD Mutant Type | ||||||||||||||||
| Standard Drug | Ceftriaxone | −1.68 | 214.98 | 36 | 15 | 5 | 2 | 0.9477 | 1.486 | 0.55(mod) | 0.51(mod) | 0.99(-) | 0.68(mild) | 0.66(mild) | 10,000 | VI |
| H. cordata | Cholest-4,14-dien-15,20-diol-3,16-dione | 4.87 | 74.60 | 31 | 4 | 2 | 0 | 0.9931 | 4.137 | 0.87(-) | 0.62(mild) | 0.70 (+) | 0.77(-) | 0.66(-) | 5000 | V |
| Dihydrocelastrol | 6.15 | 77.75 | 33 | 4 | 3 | 1 | 0.9905 | 2.415 | 0.63(mild) | 0.51(mod) | 0.73 (+) | 0.88(-) | 0.84(-) | 1000 | IV | |
| Isoquercitrin | −0.36 | 210.50 | 33 | 12 | 8 | 2 | 0.6468 | 3.076 | 0.82(-) | 0.85(-) | 0.66(mild) | 0.76(-) | 0.69(mild) | 5000 | V | |
| Naltrindole | 3.80 | 68.72 | 31 | 5 | 3 | 0 | 0.9848 | 4.214 | 0.89(-) | 0.58(mod) | 0.96(-) | 0.57(mod) | 0.55(mod) | 402 | IV | |
| Pirenperone | 3.49 | 54.69 | 29 | 5 | 0 | 0 | 0.9896 | 2.368 | 0.78(-) | 0.63(mild) | 0.99(-) | 0.54(mod) | 0.68(mild) | 1000 | IV | |
| Quercitrin | 1.68 | 131.35 | 22 | 7 | 5 | 0 | 0.9833 | 2.559 | 0.69(mild) | 0.68(mild) | 0.87(-) | 0.51(mod) | 0.99(-) | 159 | III | |
| Rhodoxanthin | 9.29 | 34.14 | 42 | 2 | 0 | 2 | 0.9902 | 2.26 | 0.63(mild) | 0.61(mild) | 0.60(mild) | 0.90(-) | 0.83(-) | 10,000 | VI | |
| Sesamin | 3.69 | 55.40 | 26 | 6 | 0 | 0 | 0.9871 | 0.967 | 0.81(-) | 0.65(mild) | 0.84(+) | 0.60(mild) | 0.94(-) | 1500 | III | |
| Usambarensine | 6.17 | 47.71 | 33 | 4 | 2 | 1 | 0.9970 | 2.689 | 0.91(-) | 0.71(-) | 0.86(+) | 0.50(mod) | 0.66(mild) | 370 | IV | |
| S. aromaticum | Biflorin | −0.70 | 160.81 | 25 | 9 | 6 | 1 | 0.9009 | 2.995 | 0.81(-) | 0.78(-) | 0.81(-) | 0.51(mod) | 0.83(-) | 562 | IV |
| Crategolic acid | 5.81 | 77.75 | 34 | 4 | 3 | 1 | 0.9643 | 2.316 | 0.65(mild) | 0.63(mild) | 0.61(mild) | 0.87(-) | 0.89(-) | 2000 | IV | |
| Oleanolic acid | 6.72 | 57.53 | 33 | 3 | 2 | 1 | 0.9853 | 2.034 | 0.52(mod) | 0.57(mod) | 0.79(+) | 0.85(-) | 0.99(-) | 2000 | IV | |
| Rhamnetin | 2.22 | 120.36 | 23 | 7 | 4 | 0 | 0.9840 | 2.542 | 0.73(mild) | 0.59(mod) | 0.55(mod) | 0.69(mild) | 0.91(-) | 5000 | V | |
| M. speciosa Korthi | Chlorogenic acid | −0.45 | 164.74 | 25 | 9 | 6 | 1 | 0.3251 | 2.277 | 0.72(-) | 0.68(mild) | 0.99(+) | 0.93(-) | 0.80(-) | 5000 | V |
| Kabasura kudineer | ||||||||||||||||
| C. serratum | Acteoside | −0.45 | 245.29 | 44 | 15 | 9 | 3 | 0.81(-) | 0.81(-) | 0.99(+) | 0.87(-) | 0.77(-) | 5000 | V | ||
| Serratagenic acid | 5.43 | 94.83 | 35 | 5 | 3 | 1 | 0.9853 | 2.233 | 0.69(mild) | 0.55(mod) | 0.79(+) | 0.90(-) | 0.91(-) | 6176 | VI | |
| Verbascoside | −0.45 | 245.29 | 44 | 15 | 9 | 3 | 0.6642 | 2.694 | 0.81(-) | 0.81(-) | 0.99(+) | 0.87(-) | 0.77(-) | 5000 | V | |
| H. auriculata | Cucurbitacin B | 2.83 | 138.20 | 40 | 8 | 3 | 1 | 0.9895 | 4.041 | 0.87(-) | 0.50(mod) | 0.90(+) | 0.72(-) | 0.66(mild) | 14 | II |
| A. paniculata | Neoandrographolide | 1.17 | 125.69 | 34 | 8 | 4 | 0 | 0.8124 | 3.165 | 0.92(-) | 0.88(-) | 0.97(+) | 0.69(mod) | 0.70(+) | 5 | I |
| M. cerviana | Orientin | 0.03 | 201.27 | 32 | 11 | 8 | 2 | 0.8864 | 3.207 | 0.81(-) | 0.72(-) | 0.52(mod) | 0.52(mod) | 0.87(-) | 1213 | IV |
| Vitexin | 0.52 | 181.04 | 31 | 10 | 7 | 1 | 0.8984 | 2.724 | 0.81(-) | 0.72(-) | 0.82(-) | 0.52(mod) | 0.87(-) | 832 | IV | |
| T. cordifolia | Tinosporide | 2.02 | 98.51 | 27 | 7 | 1 | 0 | 0.9589 | 2.775 | 0.82(-) | 0.6(mild) | 0.96(+) | 0.72(-) | 0.53(mod) | 280 | III |
| Phytocompounds | Drug-Likeness | Toxicity Analysis | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Name | Compound Name | miLogP | TPSA | Natoms | nON | nOHNH | No. of Violations | Intestinal Absorption | Oral Toxicity (−log kg/mol) | Hepato Toxicity | Carcino Genicity | Immuno Toxicity | Muta Genicity | Cyto Toxicity | LD50 (mg/kg) | TC |
| B.1.617.2. S-Protein—RBD Domain | ||||||||||||||||
| H. cordata | Canthaxanthin | 9.29 | 34.14 | 42 | 2 | 0 | 2 | 0.8350 | 3.016 | 0.63(mod) | 0.68(mod) | 0.92(-) | 0.98(-) | 0.85(-) | 10,000 | VI |
| Cholest-4,14-dien-15,20-diol-3,16-dione | 4.87 | 74.60 | 31 | 4 | 2 | 0 | 0.9931 | 4.137 | 0.87(-) | 0.62(mild) | 0.70(+) | 0.77(-) | 0.66(-) | 5000 | V | |
| Fluorometholone 17-acetate | 3.09 | 80.67 | 30 | 5 | 1 | 0 | 0.6371 | 3.218 | 0.86(-) | 0.61(mod) | 0.99(-) | 0.94(-) | 0.71(mild) | 4000 | V | |
| Kanzonol V | 7.05 | 62.83 | 28 | 4 | 2 | 1 | 0.6530 | 2.860 | 0.73(mild) | 0.59(mod) | 0.98(-) | 0.56(mod) | 0.84(-) | 2500 | V | |
| Progeldanamycin | 2.47 | 108.25 | 34 | 7 | 4 | 0 | 0.5100 | 3.001 | 0.63(mod) | 0.59(mod) | 0.99(-) | 0.719(mild) | 0.70(mild) | 1000 | IV | |
| Quercetin | 1.68 | 131.35 | 22 | 7 | 5 | 0 | 0.4381 | 2.636 | 0.69(mod) | 0.68(mod) | 0.87(-) | 0.51(+) | 0.99(-) | 159 | III | |
| Rhodoxanthin | 9.29 | 34.14 | 42 | 2 | 0 | 2 | 0.8190 | 2.660 | 0.63(mod) | 0.61(mod) | 0.60(mod) | 0.90(-) | 0.83(-) | 10,000 | VI | |
| Stigmastane-3,6-dione, (5.alpha.) | 7.76 | 34.14 | 31 | 2 | 0 | 1 | 0.8690 | 2.962 | 0.78(mild) | 0.62(mod) | 0.99(-) | 0.93(-) | 0.58(mod) | 775 | IV | |
| Rhamnetin | 2.22 | 120.36 | 23 | 7 | 4 | 0 | 0.5620 | 2.739 | 0.73(mild) | 0.59(mod) | 0.55(mod) | 0.69(mod) | 0.91(-) | 5000 | V | |
| Mitragyna speciosa Korthi | Beta-Sitosterol | 8.62 | 20.23 | 30 | 1 | 1 | 1 | 0.9241 | 3.181 | 0.87(-) | 0.60(mod) | 0.99(+) | 0.98(-) | 0.94(-) | 890 | IV |
| Stigmasterol | 7.87 | 20.23 | 30 | 1 | 1 | 1 | 0.9241 | 3.251 | 0.87(-) | 0.60(mod) | 0.99(+) | 0.98(-) | 0.94(-) | 890 | IV | |
| Kabasura kudineer | ||||||||||||||||
| T. cordifolia | Berberine | 0.20 | 40.82 | 25 | 5 | 0 | 0 | 0.4693 | 2.785 | 0.82(-) | 0.56(mod) | 0.99(-) | 0.62(mod) | 0.96(-) | 200 | III |
| C. serratum | Clerodermic acid | 2.73 | 63.60 | 24 | 4 | 1 | 0 | 0.7051 | 3.101 | 0.82(-) | 0.55(mod) | 0.81(-) | 0.89(-) | 0.80(-) | 3300 | V |
| C. speciosus | Diosgenin | 5.93 | 38.70 | 30 | 3 | 1 | 1 | 0.7820 | 3.364 | 0.69(mod) | 0.55(mod) | 0.99(-) | 0.96(-) | 0.99(-) | 8000 | VI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bharathi, M.; Sivamaruthi, B.S.; Kesika, P.; Thangaleela, S.; Chaiyasut, C. In Silico Screening of Potential Phytocompounds from Several Herbs against SARS-CoV-2 Indian Delta Variant B.1.617.2 to Inhibit the Spike Glycoprotein Trimer. Appl. Sci. 2022, 12, 665. https://doi.org/10.3390/app12020665
Bharathi M, Sivamaruthi BS, Kesika P, Thangaleela S, Chaiyasut C. In Silico Screening of Potential Phytocompounds from Several Herbs against SARS-CoV-2 Indian Delta Variant B.1.617.2 to Inhibit the Spike Glycoprotein Trimer. Applied Sciences. 2022; 12(2):665. https://doi.org/10.3390/app12020665
Chicago/Turabian StyleBharathi, Muruganantham, Bhagavathi Sundaram Sivamaruthi, Periyanaina Kesika, Subramanian Thangaleela, and Chaiyavat Chaiyasut. 2022. "In Silico Screening of Potential Phytocompounds from Several Herbs against SARS-CoV-2 Indian Delta Variant B.1.617.2 to Inhibit the Spike Glycoprotein Trimer" Applied Sciences 12, no. 2: 665. https://doi.org/10.3390/app12020665