1. Introduction
Heavy metals and metalloids (HMs) such as cadmium (Cd), arsenic (As), chromium (Cr), copper (Cu), mercury (Hg), nickel (Ni), lead (Pb) and zinc (Zn) are dispersed in the environmental media [
1,
2,
3], and are persistent pollutants which may be transported from one environmental medium to another, or may be absorbed by the apoplast of plant roots from the pedosphere. They wend their way therefrom to the edible/non-edible parts of the plants, enter the trophic chain, and imperil human health [
4,
5]. Human beings are commonly exposed to Pb, Cd, Cu, As and Cr through the ingestion of contaminated food items [
6,
7]. For lead, cadmium and copper—the three metals of interest to the authors of this study—the Food and Agricultural Organisation and the World Health Organisation [
8] have set the tolerable upper limits in food commodities as 10, 0.3 and 5 ppm respectively. Recently-published papers have documented the occurrence of toxic HMs in both cultivated [
9] and non-cultivated soils [
10,
11] in many regions of the world. Some heavy metals are essential micronutrients for plant growth (Cu, Fe, Zn etc.) and are indispensable for some of the metabolic processes (enzymatic activity, electron transfer, redox catalysis etc.) associated with plant growth [
3,
4]. On the other hand, As, Pb, Cd and Hg are detrimental to plant metabolism and are among the top 20 toxic substances, as per United States Environmental Protection Agency (USEPA) and the Agency for Toxic Substances and Disease Registry (ATSDR) [
5,
6].
In the arable pedosphere, the high concentrations of HMs and their subsequent bio-availability can be attributed to anthropogenic agricultural routines, which include the inefficient use of synthetic fertilizers and pesticides [
7]. In many arid and semi-arid regions of the world, wastewater (partially treated) is used for irrigation, and this is a conduit for HMs to reach the soil [
8]. Sewage sludge with varying concentrations of HMs is considered as a fertilizer in some parts of the world, and while being a source of essential macronutrients, it also introduces HMs to the soil. Nowadays, phytoremediation (cleaning up contaminated soils with the aid of HM-absorbing plants) is a green strategy that comprises phytoextraction, rhizo-filtration, phytodegradation, phytostabilization, and phytovolatilization for HMs in soil–food crop subsystems [
9,
10].
Heavy metals are significant environmental pollutants, and their toxicity is a problem of increasing significance for ecological, evolutionary, nutritional and environmental reasons. The environment is defined as the totality of circumstances surrounding an organism or group of organisms, especially the combination of external physical conditions that affect and influence the growth, development and survival of organisms [
11]. A pollutant is any substance in the environment which causes objectionable effects, impairing the welfare of the environment, reducing the quality of life and which may eventually cause death. Such a substance has to be present in the environment beyond a set or tolerance limit. Hence, environmental pollution is the presence of a pollutant in the environment in air, water or soil, which may be poisonous or toxic and will cause harm to living things in the polluted environment.
The regulatory limit of cadmium (Cd) in agricultural soil is 100 mg/kg soil. Plants grown in soil containing high levels of Cd show visible symptoms of injury reflected in terms of chlorosis, growth inhibition, browning of root tips and finally death [
11]. Cd has been shown to interfere with the uptake, transport and use of several elements (Ca, Mg, P and K) and water by plants [
12]. Cd also reduced the absorption of nitrate and its transport from roots to shoots, by inhibiting the nitrate reductase activity in the shoots [
13]. Copper (Cu) is considered as a micronutrient for plants and plays an important role in CO
2 assimilation and ATP synthesis. Cu is also an essential component of various proteins such as plastocyanin, of the photosynthetic system and of cytochrome oxidase of the respiratory electron transport chain [
14], but an excess of Cu in soil plays a cytotoxic role, induces stress and causes injury to plants. This leads to plant growth retardation and leaf chlorosis. Exposure of plants to excess Cu generates oxidative stress and ROS [
15]. Oxidative stress causes disturbance of metabolic pathways and damage to macromolecules [
16].
The beneficial phytoremediative potential of plant growth-promoting rhizobacteria (PGPR) has been proven in studies conducted on cereal (rice, maize and wheat) crops by Yuan et al. [
17], Nagajyoti et al. [
18] and Guo et al. [
19]. PGPR in the roots and rhizosphere greatly reduce heavy metal stress in plants by secreting organic acids, subsequent production of siderophores, 1-aminocyclopropane-1-carboxylic (ACC)-deaminase, phytohormones, chelation, immobilization, and enzymatic transformation [
13,
20]. Ramtek et al. [
21] have recommended the cultivation of certain plant germoplasms possessing the ability to accumulate HMs. Generating awareness among farmers (the primary stakeholders in this regard) will be a sine qua non to incorporate and popularize this phytoremediation technology.
Leguminous crops belonging to the Fabaceae group, such as
Vicia faba (broad beans),
Pisum sativum (garden peas),
Phasiolus vulgaris (common beans), and
Lens culinaris (lentils—white, green, red and yellow), are key components of human diets globally, as they are sources of carbohydrates, proteins, vitamins and minerals [
22]. It must not be forgotten that they were among the first outputs from poor and degraded soils. The adaptability of crops from the Fabaceae group and their ability to tolerate high salinity, temperatures and even droughts, yielded different varieties in different environments (pedo-climatic), and enabled the restoration of arid ecosystems, as noted by Belimov et al. [
23]. Legumes are noteworthy for their ability to capture and fix atmospheric nitrogen in the soil, through a mutualistic symbiotic relationship with soil-dwelling rhizobia bacteria. They also tend to absorb excessive amounts of HMs and accumulate them in their edible and non-edible parts [
24], thus posing a clear risk to animals and humans downstream in the trophic chain [
25]. Ironically, by virtue of this ability, legumes are looked upon as agents of soil-phytoremediation, when the pollutants of concern are HMs [
16,
26,
27]. It goes without saying that whenever they are called upon to fulfill this function, they cease to be food-crops.
It has been reported that the bioaccumulation of heavy metals in vegetables, especially in leafy vegetable [
28], is influenced by many factors, including climate, atmospheric deposition, soil HM concentrations, soil-type, and the degree of maturity of plants when they are harvested [
29]. The concentrations of heavy metals in the plant vary according to the type of plant [
30]. The ability of legumes to grow in marginal soils is often attributed to the symbiotic associations they have established with nitrogen-fixing rhizobia, and some of them tolerate extreme environmental conditions such as salinity, drought, or high temperatures. Khan et al. [
31] reported that cobalt (Co), lead (Pb) and chromium (Cr) are accumulated in faba bean leaves, Fe in pods, while Zn and Cu tend to be accumulated in the seeds. While a handful of studies have been carried out on the accumulation of Pb, Cd and Cu in legumes, it is worthwhile to pose new research questions and try to answer them in this study. Therefore, the authors have decided to investigate the locations and the degrees of accumulation (bifurcated as ‘accumulators’ and ‘hyper-accumulators’) of the three HMs selected for the analysis. Such an approach may considerably upgrade all procedures aimed at selecting heavy metal-tolerant varieties to be exploited for cultivation in contaminated soil. Alternative options should be carried out in order to prevent excessive accumulation of heavy metals and all vegetables should be washed properly before consumption, as washing can remove a significant amount of aerial contamination from the vegetable surface.
In Tunisia, soil polluted with heavy metals is gradually increasing due to the scarcity of rains and the use of recycled wastewater for irrigation as is often the case in semiarid regions. In legumes, heavy metals causes various physiological and biochemical alterations and diverse toxicity symptoms such as chlorosis and necrosis [
32]. Despite the acquired knowledge in relation to the response of species to heavy metals, there is a gap in relevant research fields for grain legumes. Considering that grain legumes act as accumulator plants for some heavy metals, this study aimed at investigating the response of two local varieties of grain legumes to lead, copper and cadmium at different growth stages. As such, two Tunisian varieties of common bean and faba bean were subjected to different concentrations of lead, copper and cadmium at development stage, flowering stage and pods stage, and their response was assessed on the basis of the translocation factor (TF) and the biochemical responses of the crops (chlorophyll and carotenoid contents).
4. Discussion
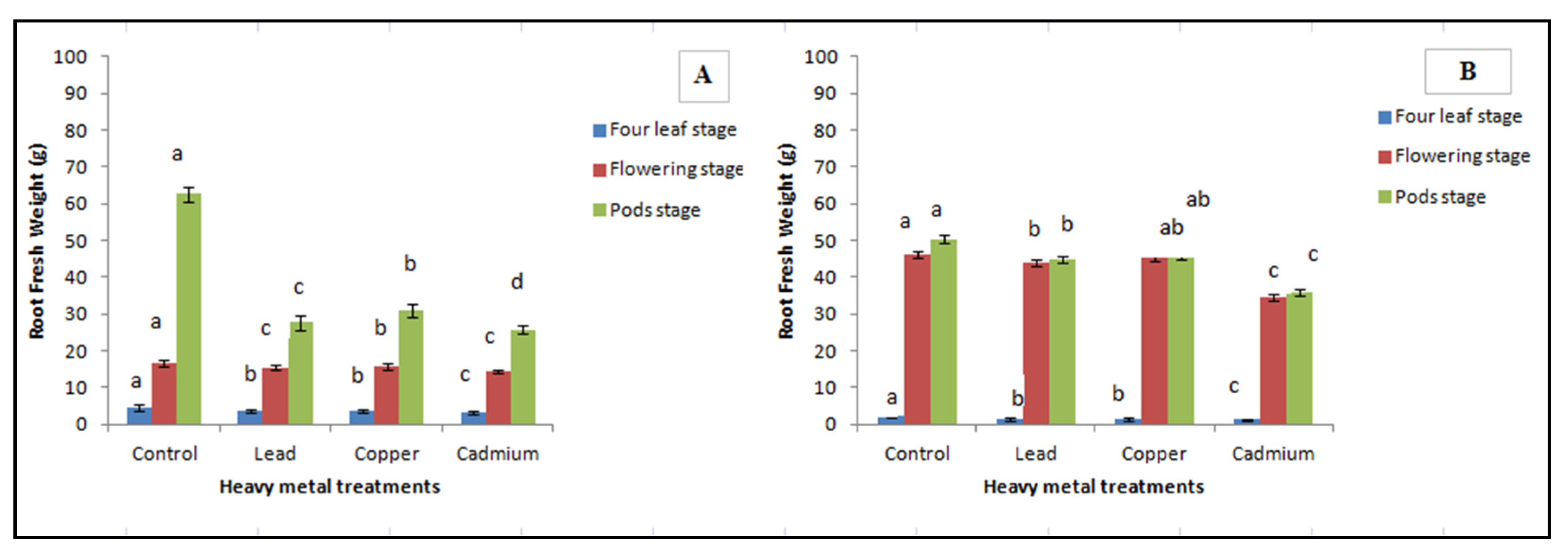
In this experimental analysis, the authors focused on two varieties of bean in Tunisia—the common bean and the faba bean, and embarked on investigating the effect of three HMs—Pb, Cu and Cd—on a range of plant-growth aspects, at different stages of plant growth. The addition of HMs to the soil had a detrimental effect on plant growth, with the RFW and SFW being lower vis-à-vis the uncontaminated control samples, in accordance with the results published in [
34] for the effects of toxic metals on Arabidopsis. Baldi et al. [
16] have noted that the root growth of common bean significantly decreased (
p < 0.05) with the increase in soil Pb concentration. In this study, the authors considered two other HMs—Cd and Cu—and showed that the adverse growth-inhibitory effects varied among the HMs [
34] (
Figure 1 and
Figure 2). Cadmium turned out to have the most negative effect and copper the least. The negative effect could be due to the suppression of the elongation growth rate of cells, resulting in an irreversible inhibition exerted by Cd on the proton pump responsible for the process [
35].
Additionally, the exposure of the two varieties of beans to these three HMs resulted in a reduction in chlorophyll and carotenoid contents (
Table 2). Baek et al. [
34] have observed that abiotic stresses—which includes inhibition by toxic metals—on plants tend to affect their chlorophyll biosynthesis capability. Cd and Pb tend to inhibit enzymatic activity of δ-aminolevulinic acid dehydratase and protochlorophyllide reductase associated with chlorophyll biosynthesis. Copper, being an essential micronutrient, was found to be less toxic than the other two HMs, as expected [
36]. If one would compare common bean with faba bean, the former was affected more than the latter [
37].
However, Zhang et al. [
38] have reported that HMs can either enhance or decrease carotenoid production, and that would depend on the type of HM. Carotenoids protect chlorophyll pigments under heavy-metal-induced stress conditions [
39,
40], and thus if their biosynthesis is disturbed, it would directly hurt the chlorophyll content in the leaves of plants [
41,
42].
Bioaccumulation and translocation of the HMs were also studied in this experiment. In addition to the quantification of the accumulation of these HMs on roots, shoots, flowers and pods, the BAFs and TFs were also calculated. Roots accumulate a lot of HMs (in this case, Cd and Pb especially), when the plants are in the vegetative stages of growth (4th mature leaf stage) [
43]. While the point of entry for the HMs—from the soil to the roots—is the apoplast of the root-cortex and the endoderm [
44,
45], the upward transport from thereto the stem, branches, leaves, flowers and fruits is termed as translocation. This may or may not require complexing agents such as organic acids, amino acids and peptides [
46]. Suman et al. [
47] are of the view that metallic trace elements often preferentially accumulate in roots (as Pb does in the case of this study). The low TF values for lead are corroborated by what has been reported by [
48,
49]. Lead is tightly retained by the root tissues. Faba bean, which has shown higher TF values in general for all the three HMs studied, can be posited as an effective phytoextracting agent in rhizofiltration facilities [
8]. However, post-extraction, they will not be available as food commodities. While HMs prefer in general to make the roots their permanent abodes’, Aboulroos et al. [
50] have reported that they prefer the leaves next and the stem thereafter.
The variation of the HM concentrations for common bean and faba bean at the 4th leaf stage can be explained by the sorption phenomenon exhibited by the HMs in the substrate. The exchanges between the liquid and solid phases of the soil depend on the sorption mechanism referred to. After being adsorbed on the surface of soil particles, the HMs can be absorbed by the mineral particles. A TF value for Pb, Cu and Cd less than unity (
Table 4) in the 4th leaf stage, for both the varieties, suggests that the HMs are not effectively transferred from the roots to the above-ground parts of the plants [
50].
The results obtained in this study concur with those of [
51] who showed that with a Pb concentration in the range of 9 to 267 μM for
Arabispaniculata Franch, the TF ratio was less than unity. A TF value greater than one was reported for
Solanumnigrum by [
51], suggesting that this plant species can easily transfer lead from the soil to airborne organisms. To reiterate, hyper-accumulators are plants which are effective in moving HMs up from roots to the stems and leaves, and thereby have TF ratios greater than unity [
52].
In the vegetative stage, as mentioned earlier too, HMs (lead especially) tend to accumulate in the roots, and thus the two varieties of beans considered in this particular study cannot be hyper-accumulators of lead in this stage of growth. However, when it comes to copper, the TF is close to unity, indicating that copper is transferred from the soil to the roots to the aerial parts. This, as mentioned on a few occasions earlier in this article, is because copper is an essential micronutrient for plant growth, especially in the vegetative stage. The TF values for Cd obtained in this study are lower than those for copper in the vegetative stage. Zhao et al. [
53], however, have reported higher TF ratios for Cd in the vegetative stage.
For a plant to be an effective phytoremediating agent, its BAF must be greater than unity [
54]. In this study, the BAFs for Pb, Cu and Cd in the vegetative stage of common bean and faba bean were 0.43, 0.81, 0.12, and 0.15, 0.94, 0.14, respectively. One can conclude from these values that
Vicia faba cannot be an effective phytoremediating agent in the vegetative stage [
55].
Most HMs are insoluble in the vascular system of plants, and are thereby less mobile. This is owing to their presence as sulphate, phosphate or carbonates, which immobilises them in the apoplastic (extracellular) and symplastic compartments of the vascular system [
56]. The WHO proposes limits of 10 mg/kg biomass and 0.3 mg/kg biomass for Pb and Cd, respectively. As can be seen in
Table 4, the Pb and Cd concentrations in the roots, stems, leaves and flowers recorded in this study are above this tolerable upper limit prescribed by the WHO [
8].
When it comes to copper, its uptake by plants depends on the ability of plants to take it up through the soil–root interface as a divalent cation or chelated copper, and also on the total amount of Cu present in the soil. Faba bean, as this study showed, is a hyper-accumulator of copper. As reported by Memon et al. [
57], if there are free metals available for translocation from roots to shoots, it indicates that the sequestration of these metals in root vacuoles is limited. Copper, though necessary at low doses, is often present in soils at concentrations which may be quite toxic to plants growing there. Values of 0.4 to 45.8 mg Cu per kg of soil have been reported for uncontaminated soils, in [
58]. Plants which are tolerant to high concentrations of toxic heavy metals are usually capable of compartmentalizing metal ions, sequestering them in vacuoles and thus excluding them from cell-sites where processes such as cell division and respiration occur [
58].
Singh et al. [
59] reported that the process of translocating HMs in plant species is a crucial factor in determining the distribution of these elements in different plant tissues. The HMs are mobilized and removed by the root cells, bound by the cell wall and then transported through the plasma membrane, driven by proton pumps dependent on ATP. In addition to cationic nutrients, plant transporters also participate in the migration of potentially toxic cations between plant membranes [
60]. This study yielded a TF value of less than unity for lead in the full-flowering stage, indicating a propensity for lead to lodge itself in the roots preferentially, vis-à-vis Cu and Cd. This can lead to the conclusion that the common bean and faba bean are not hyper-accumulators for lead in the vegetative stage. The roots of leguminous plants such as common bean and faba bean secrete exudates necessary for the formation of nodules that facilitate the mobilization and translocation of metals. With TFs for Cu greater than unity in the flowering stage, faba bean plants are hyper-accumulators for the red metal, but just accumulators for Cd (the TF value being less than one). Farmers can thus avail themselves of common bean and faba bean crops as phyto-remediating agents to cleanse the soil of Cd (by locking it up in the roots). After removing these crops once enough Cd has been extracted, the soil is much less contaminated then (at least with respect to Cd). In the flowering stage, these two crops are hyper-accumulators of Pb and Cd (BAFs in the range of 1.08 to 2.42).
In the fruiting stage, the stems, leaves and pods of the plants accumulated more Cu than Pb and Cd, for the simple reason that Cu is a catalyst in electron transfer and redox reactions involving enzymatic reactions and protein synthesis [
60]. Environmental factors, plant types and the stage of growth of the plant influence the bioaccumulation of HMs. As already discussed before, in very high concentrations, even the essential micronutrients such as copper can be toxic to plants. In the pods stage, both common bean and faba bean are capable of taking up Pb, Cu and Cd, and subsequently sequestering them in the vacuoles of the root system. As the TFs for Cu and Cd are greater than unity in this stage of plant growth for both the varieties, they are hyper-accumulators for these two HMs in the pods-stage, as also concluded by [
31] for the Brassica species of plants.



 ,
, 



{kind=link}
{kind=link}