
Polysaccharides from Moroccan Green and Brown Seaweed and Their Derivatives Stimulate Natural Defenses in Olive Tree Leaves
 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Samples
2.2. Extraction and Purification of Polysaccharides and Preparation of Oligomers Extracts
2.3. Biochemical Characterization of Extracted Polysaccharides
2.3.1. Total Sugar Content
2.3.2. Neutral Sugar and Uronic Acid Content
2.3.3. Sulfates Content
2.3.4. Protein and Phenolic Content
2.4. Structural Analysis of Saccharides Fractions
2.5. Treatment of Leaf Discs of the Olive Tree
2.6. Measurement of Enzyme Activities and Protein Assays
2.7. Quantification of Secondary Metabolites: Phenolic Compounds and Lignin
2.8. Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition of Polysaccharide Fractions
3.2. SEC–MALLS Analysis of Saccharide Fractions
3.3. Monosaccharide Composition of Saccharide Fractions
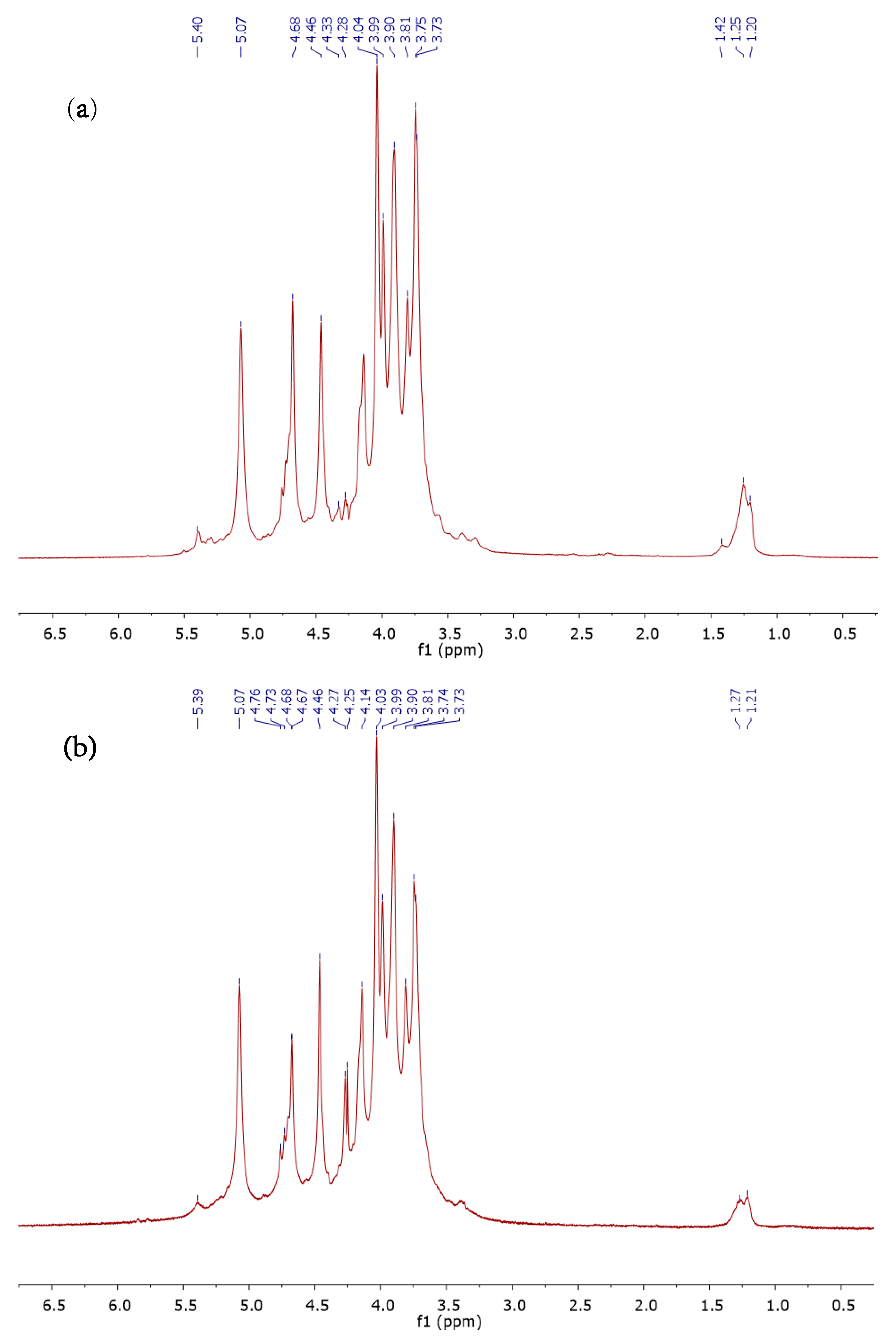
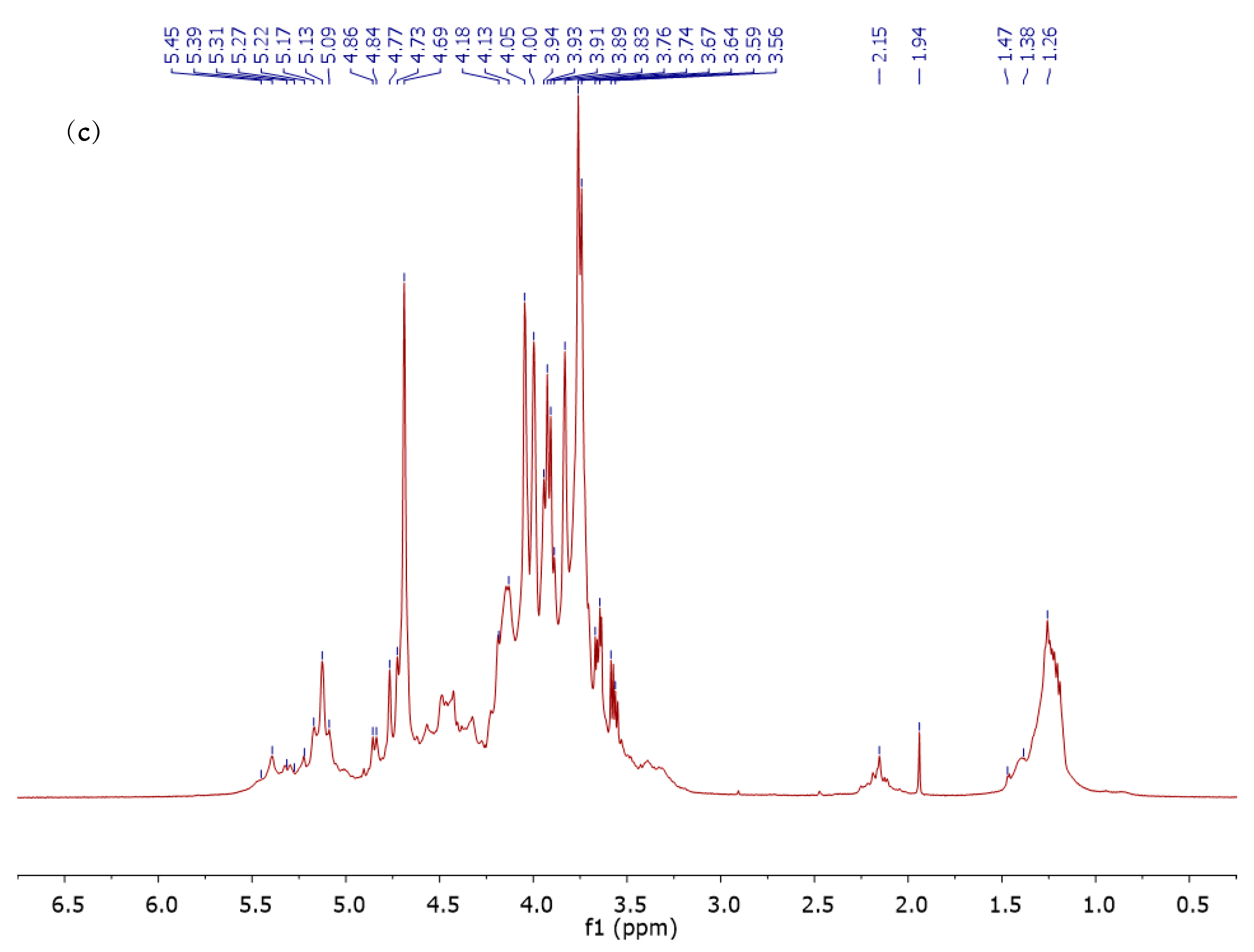
3.4. 1H NMR Analysis of ALCM, ALSM, and FUCCM Fractions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fucose | H-1 | H-2 | H-3 | H-4 | H-5 | H-6 |
|---|---|---|---|---|---|---|
| α-l-Fucp [59] | 5.20 | 3.77 | 3.86 | 3.81 | 4.20 | 1.21 |
| α-l-Fucp (FUCCM 1) | 5.13 (−0.07) | 3.76 (−0.01) | 3.89 (0.03) | 3.83 (0.02) | 4.17 (−0.03) | 1.26 (0.05) |
3.5. Sulfated and Unsulfated Polysaccharides and Oligosaccharides induce Natural Defenses in Olive Tree Leaves
3.5.1. Evaluation of Enzyme Activities and Secondary Metabolism Synthesis in the Leaf Discs of the Olive Tree in Response to Alginates and Their Oligomers
3.5.2. Evaluation of Enzyme Activities and Secondary Metabolism Synthesis in the Leaf Discs of the Olive Tree in Response to Sulfated Polysaccharides AGB, AGP and FUCCM
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2010, 344, 1–50. [Google Scholar] [CrossRef]
- Eurostat. Eurostat Statistics Explained. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Agricultural_production_-_orchards#Olive_trees (accessed on 1 April 2022).
- Eurostat. Agriculture, Forestry and Fishery Statistics, 2018th ed.; Eurostat: Bietlot Frères, Belgium, 2018; p. 200. [Google Scholar]
- Chliyeh, M.; Ouazzani Touhami, A.; Filali-Maltouf, A.; El Modafar, C.; Moukhli, A.; Oukabli, A.; Benkirane, R.; Douira, A.; Bioscience, A. Effect of a composite endomycorrhizal inoculum on the growth of olive trees under nurseries conditions in morocco. Int. J. Pure Appl. Biosci. 2014, 2, 1–14. [Google Scholar]
- El Mouhtadi, I.; Agouzzal, M.; Guy, F.J.O. L’olivier au maroc. OCL 2014, 21, D203. [Google Scholar] [CrossRef]
- Achbani, E.H.; Benbouazza, A.; Douira, A. First report of olive anthracnose, caused by colletotrichum gloeosporioides, in morocco. Atlas J. Biol. 2013, 2, 172–175. [Google Scholar] [CrossRef]
- Rugini, E.; Baldoni, L.; Muleo, R.; Sebastiani, L. The Olive Tree Genome; Springer: Berlin/Heidelberg, Germany, 2016; Volume 978. [Google Scholar]
- Kour, D.; Bakshi, P.; Wali, V.K.; Sharma, N.; Sharma, A.; Iqbal, M. Alternate bearing in olive—A review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2281–2297. [Google Scholar] [CrossRef]
- Gómez-Lama Cabanás, C.; Ruano-Rosa, D.; Legarda, G.; Pizarro-Tobías, P.; Valverde-Corredor, A.; Triviño, J.; Roca, A.; Mercado-Blanco, J. Bacillales members from the olive rhizosphere are effective biological control agents against the defoliating pathotype of verticillium dahliae. Agriculture 2018, 8, 90. [Google Scholar] [CrossRef]
- Nigro, F.; Antelmi, I.; Labarile, R.; Sion, V.; Pentimone, I. Biological control of olive anthracnose. Acta Hortic. 2018, 1199, 439–444. [Google Scholar] [CrossRef]
- Bueno, A.M.; Jones, O. Alternative methods for controlling the olive fly, bactrocera oleae, involving semiochemicals. IOBC Wprs Bull. 2002, 25, 147–156. [Google Scholar]
- Bizos, G.; Papatheodorou, E.M.; Chatzistathis, T.; Ntalli, N.; Aschonitis, V.G.; Monokrousos, N. The role of microbial inoculants on plant protection, growth stimulation, and crop productivity of the olive tree (Olea europea L.). Plants 2020, 9, 743. [Google Scholar] [CrossRef]
- Dzhavakhiya, V.G.; Shcherbakova, L.A. Control of plant pathogens by biogenic elicitors and possible mechanisms of action. Front. Plant Sci. 2016, 7, 340. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (isr) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Bedoux, G.; Hardouin, K.; Burlot, A.S.; Bourgougnon, N. Bioactive components from seaweeds. In Sea Plants; Academic Press: Cambridge, MA, USA, 2014; pp. 345–378. [Google Scholar]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Seaweed-based compounds and products for sustainable protection against plant pathogens. Mar. Drugs 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.K.; Dangariya, M.; Agarwal, P. Seaweed extracts: Potential biodegradable, environmentally friendly resources for regulating plant defence. Algal Res. 2021, 58, 102363. [Google Scholar] [CrossRef]
- Aitouguinane, M.; Bouissil, S.; Mouhoub, A.; Rchid, H.; Fendri, I.; Abdelkafi, S.; Ould El-Hadj, M.D.; Boual, Z.; Dubessay, P.; Gardarin, C.; et al. Induction of natural defenses in tomato seedlings by using alginate and oligoalginates derivatives extracted from moroccan brown algae. Mar. Drugs 2020, 18, 521. [Google Scholar] [CrossRef]
- Bouissil, S.; Alaoui-Talibi, Z.E.; Pierre, G.; Rchid, H.; Michaud, P.; Delattre, C.; El Modafar, C. Fucoidans of moroccan brown seaweed as elicitors of natural defenses in date palm roots. Mar. Drugs 2020, 18, 596. [Google Scholar] [CrossRef]
- Bouissil, S.; El Alaoui-Talibi, Z.; Pierre, G.; Michaud, P.; El Modafar, C.; Delattre, C. Use of alginate extracted from moroccan brown algae to stimulate natural defense in date palm roots. Molecules 2020, 25, 720. [Google Scholar] [CrossRef]
- El Modafar, C.; Elgadda, M.; El Boutachfaiti, R.; Abouraicha, E.; Zehhar, N.; Petit, E.; El Alaoui-Talibi, Z.; Courtois, B.; Courtois, J. Induction of natural defence accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to bioelicitors isolated from green algae. Sci. Hortic. 2012, 138, 55–63. [Google Scholar] [CrossRef]
- Abouraïcha, E.F.; El Alaoui-Talibi, Z.; Tadlaoui-Ouafi, A.; El Boutachfaiti, R.; Petit, E.; Douira, A.; Courtois, B.; Courtois, J.; El Modafar, C. Glucuronan and oligoglucuronans isolated from green algae activate natural defense responses in apple fruit and reduce postharvest blue and gray mold decay. J. Appl. Phycol. 2016, 29, 471–480. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against penicillium expansum and botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hortic. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Chandia, N.P.; Matsuhiro, B. Characterization of a fucoidan from lessonia vadosa (phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar] [CrossRef]
- Mazumder, A.; Holdt, S.L.; De Francisci, D.; Alvarado-Morales, M.; Mishra, H.N.; Angelidaki, I. Extraction of alginate from sargassum muticum: Process optimization and study of its functional activities. J. Appl. Phycol. 2016, 28, 3625–3634. [Google Scholar] [CrossRef]
- Soukaina, B.; Zainab, E.L.A.-T.; Guillaume, P.; Halima, R.; Philippe, M.; Cherkaoui, E.M.; Cédric, D. Radical depolymerization of alginate extracted from moroccan brown seaweed bifurcaria bifurcata. Appl. Sci. 2020, 10, 4166. [Google Scholar] [CrossRef]
- Aitouguinane, M.; El Alaoui-Talibi, Z.; Rchid, H.; Fendri, I.; Abdelkafi, S.; Ould El-Hadj, M.D.; Boual, Z.; Dubessay, P.; Michaud, P.; Le Cerf, D.; et al. A novel sulfated glycoprotein elicitor extracted from the moroccan green seaweed codium decorticatum induces natural defenses in tomato. Appl. Sci. 2022, 12, 3643. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.; Hamilton, J.; Rebers, P.; Smith, F.J.N. A colorimetric method for the determination of sugars. Nature 1951, 168, 167. [Google Scholar] [CrossRef]
- Monsigny, M.; Petit, C.; Roche, A.-C. Colorimetric determination of neutral sugars by a resorcinol sulfuric acid micromethod. Anal. Biochem. 1988, 175, 525–530. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Montreuil, J.; Spik, G.; Chosson, A.; Segard, E.; Scheppler, N. Methods of study of the structure of glycoproteins. J. Pharm. Belg. 1963, 18, 529–546. [Google Scholar] [PubMed]
- Dodgson, K.; Price, R.J. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; De Mello, J.C.P. Application and analysis of the folin ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef]
- Grasdalen, H. High-field, 1H-nmr spectroscopy of alginate: Sequential structure and linkage conformations. Carbohydr. Res. 1983, 118, 255–260. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Bi, Y.; Luo, Y. Postharvest bth treatment induces resistance of peach (Prunus persica L. Cv. Jiubao) fruit to infection by penicillium expansum and enhances activity of fruit defense mechanisms. Postharvest Biol. Technol. 2005, 35, 263–269. [Google Scholar] [CrossRef]
- Hendrikse, N.M.; Holmberg Larsson, A.; Svensson Gelius, S.; Kuprin, S.; Nordling, E.; Syren, P.O. Exploring the therapeutic potential of modern and ancestral phenylalanine/tyrosine ammonia-lyases as supplementary treatment of hereditary tyrosinemia. Sci. Rep. 2020, 10, 1315. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Belattmania, Z.; Kaidi, S.; El Atouani, S.; Katif, C.; Bentiss, F.; Jama, C.; Reani, A.; Sabour, B.; Vasconcelos, V. Isolation and FTIR-ATR and 1H NMR characterization of alginates from the main alginophyte species of the atlantic coast of Morocco. Molecules 2020, 25, 4335. [Google Scholar] [CrossRef]
- Trica, B.; Delattre, C.; Gros, F.; Ursu, A.V.; Dobre, T.; Djelveh, G.; Michaud, P.; Oancea, F. Extraction and characterization of alginate from an edible brown seaweed (Cystoseira barbata) harvested in the romanian black sea. Mar. Drugs 2019, 17, 405. [Google Scholar] [CrossRef]
- Sellimi, S.; Younes, I.; Ayed, H.B.; Maalej, H.; Montero, V.; Rinaudo, M.; Dahia, M.; Mechichi, T.; Hajji, M.; Nasri, M. Structural, physicochemical and antioxidant properties of sodium alginate isolated from a tunisian brown seaweed. Int. J. Biol. Macromol. 2015, 72, 1358–1367. [Google Scholar] [CrossRef]
- Benslima, A.; Sellimi, S.; Hamdi, M.; Nasri, R.; Jridi, M.; Cot, D.; Li, S.; Nasri, M.; Zouari, N. The brown seaweed cystoseira schiffneri as a source of sodium alginate: Chemical and structural characterization, and antioxidant activities. Food Biosci. 2021, 40, 100873. [Google Scholar] [CrossRef]
- Hentati, F.; Delattre, C.; Ursu, A.V.; Desbrieres, J.; Le Cerf, D.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Pierre, G. Structural characterization and antioxidant activity of water-soluble polysaccharides from the tunisian brown seaweed cystoseira compressa. Carbohydr. Polym. 2018, 198, 589–600. [Google Scholar] [CrossRef]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Upgrading the antioxidant properties of fucoidan and alginate from cystoseira trinodis by fungal fermentation or enzymatic pretreatment of the seaweed biomass. Food Chem. 2018, 269, 387–395. [Google Scholar] [CrossRef]
- El Atouani, S.; Bentiss, F.; Reani, A.; Zrid, R.; Belattmania, Z.; Pereira, L.; Mortadi, A.; Cherkaoui, O.; Sabour, B. The invasive brown seaweedsargassum muticumas new resource for alginate in morocco: Spectroscopic and rheological characterization. Phycol. Res. 2016, 64, 185–193. [Google Scholar] [CrossRef]
- Gorham, J.; Lewey, S. Seasonal changes in the chemical composition of sargassum muticum. Mar. Biol. 1984, 80, 103–107. [Google Scholar] [CrossRef]
- Davis, T.A.; Llanes, F.; Volesky, B.; Mucci, A. Metal selectivity of Sargassum spp. and their alginates in relation to their alpha-l-guluronic acid content and conformation. Environ. Sci. Technol. 2003, 37, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Benslima, A.; Sellimi, S.; Hamdi, M.; Nasri, R.; Jridi, M.; Cot, D.; Li, S.; Nasri, M.; Zouari, N. Brown seaweed cystoseira schiffneri as a promising source of sulfated fucans: Seasonal variability of structural, chemical, and antioxidant properties. Food Sci. Nutr. 2021, 9, 1551–1563. [Google Scholar] [CrossRef]
- Sellimi, S.; Kadri, N.; Barragan-Montero, V.; Laouer, H.; Hajji, M.; Nasri, M. Fucans from a tunisian brown seaweed cystoseira barbata: Structural characteristics and antioxidant activity. Int. J. Biol. Macromol. 2014, 66, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Hadj Ammar, H.; Lajili, S.; Ben Said, R.; Le Cerf, D.; Bouraoui, A.; Majdoub, H. Physico-chemical characterization and pharmacological evaluation of sulfated polysaccharides from three species of mediterranean brown algae of the genus Cystoseira. DARU J. Pharm. Sci. 2015, 23, 1. [Google Scholar] [CrossRef]
- Rahelivao, M.P.; Andriamanantoanina, H.; Heyraud, A.; Rinaudo, M. Structure and properties of three alginates from madagascar seacoast algae. Food Hydrocoll. 2013, 32, 143–146. [Google Scholar] [CrossRef]
- Burkus, Z. Determination of the molecular weight of barley β-glucan using intrinsic viscosity measurements. Carbohydr. Polym. 2003, 54, 51–57. [Google Scholar] [CrossRef]
- Khouryieh, H.A.; Herald, T.J.; Aramouni, F.; Alavi, S. Intrinsic viscosity and viscoelastic properties of xanthan/guar mixtures in dilute solutions: Effect of salt concentration on the polymer interactions. Food Res. Int. 2007, 40, 883–893. [Google Scholar] [CrossRef]
- Rochefort, W.E.; Middleman, S. Rheology of xanthan gum: Salt, temperature, and strain effects in oscillatory and steady shear experiments. J. Rheol. 1987, 31, 337–369. [Google Scholar] [CrossRef]
- Moseley, R.; Waddington, R.; Evans, P.; Halliwell, B.; Embery, G. The chemical modification of glycosaminoglycan structure by oxygen-derived species in vitro. Biochim. Biophys. Acta 1995, 1244, 245–252. [Google Scholar] [CrossRef]
- Lim, S.J.; Wan Aida, W.M.; Maskat, M.Y.; Latip, J.; Badri, K.H.; Hassan, O.; Yamin, B.M. Characterisation of fucoidan extracted from malaysian sargassum binderi. Food Chem. 2016, 209, 267–273. [Google Scholar] [CrossRef]
- Zayed, A.; El-Aasr, M.; Ibrahim, A.S.; Ulber, R. Fucoidan characterization: Determination of purity and physicochemical and chemical properties. Mar. Drugs 2020, 18, 571. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Kim, W.-J.; Kim, S.-M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Il Park, Y. Structure and antitumour activity of fucoidan isolated from sporophyll of korean brown seaweed undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Daniel, R.; Berteau, O.; Chevolot, L.; Varenne, A.; Gareil, P.; Goasdoue, N. Regioselective desulfation of sulfated l-fucopyranoside by a new sulfoesterase from the marine mollusk pecten maximus: Application to the structural study of algal fucoidan (Ascophyllum nodosum). Eur. J. Biochem. 2001, 268, 5617–5626. [Google Scholar] [CrossRef]
- Barros, J.; Dixon, R.A. Plant phenylalanine/tyrosine ammonia-lyases. Trends Plant Sci. 2020, 25, 66–79. [Google Scholar] [CrossRef]
- Golan, K.; Sempruch, C.; Górska-Drabik, E.; Czerniewicz, P.; Łagowska, B.; Kot, I.; Kmieć, K.; Magierowicz, K.; Leszczyński, B. Accumulation of amino acids and phenolic compounds in biochemical plant responses to feeding of two different herbivorous arthropod pests. Arthropod-Plant Interact. 2017, 11, 675–682. [Google Scholar] [CrossRef]
- Jangaard, N.O. The characterization of phenylalanine ammonia-lyase from several plant species. Phytochemistry 1974, 13, 1765–1768. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, H.; Geng, G.; Zhang, Q.; Zhang, S. Changes in defense-related enzymes and phenolics in resistant and susceptible common wheat cultivars under aphid stress. Acta Physiol. Plant. 2021, 43, 36. [Google Scholar] [CrossRef]
- Rösler, J.; Krekel, F.; Amrhein, N.; Schmid, J. Maize phenylalanine ammonia-lyase has tyrosine ammonia-lyase activity. Plant Physiol. 1997, 113, 175–179. [Google Scholar] [CrossRef]
- Siqueira-Soares, R.d.C.; Soares, A.R.; Parizotto, A.V.; Ferrarese, M.d.L.L.; Ferrarese-Filho, O. Root growth and enzymes related to the lignification of maize seedlings exposed to the allelochemical L-DOPA. Sci. World J. 2013, 2013, 134237. [Google Scholar] [CrossRef]
- Montero, T.; Mollá, E.; Martín-Cabrejas, M.A.; López-Andréu, F. Effects of gibberellic acid (ga3) on strawberry pal (phenylalanine ammonia-lyase) and tal (tyrosine ammonia-lyase) enzyme activities. J. Sci. Food Agric. 1998, 77, 230–234. [Google Scholar] [CrossRef]
- Khan, W.; Prithiviraj, B.; Smith, D.L. Chitosan and chitin oligomers increase phenylalanine ammonia-lyase and tyrosine ammonia-lyase activities in soybean leaves. J. Plant Physiol. 2003, 160, 859–863. [Google Scholar] [CrossRef]
- Ganapathy, G.; Keerthi, D.; Nair, R.A.; Pillai, P. Correlation of phenylalanine ammonia lyase (pal) and tyrosine ammonia lyase (tal) activities to phenolics and curcuminoid content in ginger and its wild congener, zingiber zerumbet following pythium myriotylum infection. Eur. J. Plant Pathol. 2016, 145, 777–785. [Google Scholar] [CrossRef]
- Chandía, N.P.; Matsuhiro, B.; Mejías, E.; Moenne, A. Alginic acids in lessonia vadosa: Partial hydrolysis and elicitor properties of the polymannuronic acid fraction. J. Appl. Phycol. 2004, 16, 127–133. [Google Scholar] [CrossRef]
- Sangha, J.S.; Ravichandran, S.; Prithiviraj, K.; Critchley, A.T.; Prithiviraj, B. Sulfated macroalgal polysaccharides λ-carrageenan and ι-carrageenan differentially alter arabidopsis thaliana resistance to sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2010, 75, 38–45. [Google Scholar] [CrossRef]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 2001, 149, 43–51. [Google Scholar] [CrossRef]
- Ghannam, A.; Abbas, A.; Alek, H.; Al-Waari, Z.; Al-Ktaifani, M. Enhancement of local plant immunity against tobacco mosaic virus infection after treatment with sulphated-carrageenan from red alga (Hypnea musciformis). Physiol. Mol. Plant Pathol. 2013, 84, 19–27. [Google Scholar] [CrossRef]
- Sangha, J.S.; Khan, W.; Ji, X.; Zhang, J.; Mills, A.A.; Critchley, A.T.; Prithiviraj, B. Carrageenans, sulphated polysaccharides of red seaweeds, differentially affect arabidopsis thaliana resistance to trichoplusia ni (Cabbage looper). PLoS ONE 2011, 6, e26834. [Google Scholar] [CrossRef]
- Sangha, J.S.; Kandasamy, S.; Khan, W.; Bahia, N.S.; Singh, R.P.; Critchley, A.T.; Prithiviraj, B. Lambda-carrageenan suppresses tomato chlorotic dwarf viroid (tcdvd) replication and symptom expression in tomatoes. Mar. Drugs 2015, 13, 2875–2889. [Google Scholar] [CrossRef]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Carrageenans from red seaweeds as promoters of growth and elicitors of defense response in plants. Front. Mar. Sci. 2016, 3, 81. [Google Scholar] [CrossRef]
- de Borba, M.C.; Velho, A.C.; Maia-Grondard, A.; Baltenweck, R.; Magnin-Robert, M.; Randoux, B.; Holvoet, M.; Hilbert, J.L.; Flahaut, C.; Reignault, P.; et al. The algal polysaccharide ulvan induces resistance in wheat against zymoseptoria tritici without major alteration of leaf metabolome. Front. Plant Sci. 2021, 12, 703712. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, M.B.; Ferreira, L.G.; Hawerroth, C.; Duarte, M.E.; Noseda, M.D.; Stadnik, M.J. Ulvans induce resistance against plant pathogenic fungi independently of their sulfation degree. Carbohydr. Polym. 2015, 133, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Jaulneau, V.; Lafitte, C.; Jacquet, C.; Fournier, S.; Salamagne, S.; Briand, X.; Esquerre-Tugaye, M.T.; Dumas, B. Ulvan, a sulfated polysaccharide from green algae, activates plant immunity through the jasmonic acid signaling pathway. J. Biomed. Biotechnol. 2010, 2010, 525291. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.-F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. An ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Zheng, F.; Chen, L.; Zhang, P.; Zhou, J.; Lu, X.; Tian, W. Carbohydrate polymers exhibit great potential as effective elicitors in organic agriculture: A review. Carbohydr. Polym. 2020, 230, 115637. [Google Scholar] [CrossRef]
- Reunov, A.; Lapshina, L.; Nagorskaya, V.; Zvyagintseva, T.; Shevchenko, N. Effect of fucoidan from the brown alga fucus evanescens on the development of infection induced by potato virus x in Datura stramonium L. Leaves. J. Plant Dis. Prot. 2009, 116, 49–54. [Google Scholar] [CrossRef]
- Lapshina, L.A.; Reunov, A.V.; Nagorskaya, V.P.; Zvyagintseva, T.N.; Shevchenko, N.M. Inhibitory effect of fucoidan from brown alga fucus evanescens on the spread of infection induced by tobacco mosaic virus in tobacco leaves of two cultivars. Russ. J. Plant Physiol. 2006, 53, 246–251. [Google Scholar] [CrossRef]
- Wei, X.; Cai, L.; Liu, H.; Tu, H.; Xu, X.; Zhou, F.; Zhang, L. Chain conformation and biological activities of hyperbranched fucoidan derived from brown algae and its desulfated derivative. Carbohydr. Polym. 2019, 208, 86–96. [Google Scholar] [CrossRef]
- Seto, S.P.; Miller, T.; Temenoff, J.S. Effect of selective heparin desulfation on preservation of bone morphogenetic protein-2 bioactivity after thermal stress. Bioconjugate Chem. 2015, 26, 286–293. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Rasin, A.B.; Zueva, A.O.; Kusaykin, M.I.; Zvyagintseva, T.N.; Rubtsov, N.K.; Ermakova, S.P. Discovery of a fucoidan endo-4o-sulfatase: Regioselective 4o-desulfation of fucoidans and its effect on anticancer activity in vitro. Carbohydr. Polym. 2021, 271, 118449. [Google Scholar] [CrossRef] [PubMed]
- Takada, T.; Katagiri, T.; Ifuku, M.; Morimura, N.; Kobayashi, M.; Hasegawa, K.; Ogamo, A.; Kamijo, R. Sulfated polysaccharides enhance the biological activities of bone morphogenetic proteins. J. Biol. Chem. 2003, 278, 43229–43235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tziros, G.T.; Samaras, A.; Karaoglanidis, G.S. Laminarin induces defense responses and efficiently controls olive leaf spot disease in olive. Molecules 2021, 26, 1043. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah, I.; Aghrouss, S.; Douira, A.; Aissam, S.; El Alaoui-Talibi, Z.; Filali-Maltouf, A.; El Modafar, C. Seaweed polysaccharides as bio-elicitors of natural defenses in olive trees against verticillium wilt of olive. J. Plant Interact. 2018, 13, 248–255. [Google Scholar] [CrossRef]








| Fractions | Source | Yield (% w/w) | Total Sugar (% w/w) | Neutral Sugar (% w/w) | Uronic Acid (% w/w) | Sulfate (% w/w) | Protein (% w/w) | Polyphenol (% w/w) |
|---|---|---|---|---|---|---|---|---|
| ALSM 1 | S. muticum | 23 | 44.46 ± 1.67 | 2.42 ±1.96 | 63.27 ± 0.17 | Nd 6 | Trace | Nd |
| ALCM 2 | C. myriophylloides | 31 | 57.48 ± 0.35 | 30.27 ± 0.84 | 44.99 ± 3.70 | 7.29 ±0.4 | Nd | Nd |
| FUCCM 3 | C. myriophylloides | 0.87 | 52.11 ± 3.07 | 29.06 ± 0.52 | 23.19 ± 1.56 | 12.83 ± 0.06 | 2.93 ±0.17 | Nd |
| AGB 4 | C. decorticatum | 11 | 37.67 ± 1.10 | 37.56 ± 1.73 | 3.05 ± 0.29 | 22.68 ± 2.08 | 20.03 ± 0.29 | Trace 7 |
| AGP 5 | C. decorticatum | 5 | 48.38 ± 1.44 | 45.68 ± 0.58 | 4.04 ± 0.35 | 13.21 ± 0.23 | 11.89 ± 2.48 | Trace |
| Fractions | Source | Mn 8 (g/mol) | Mw 9 (g/mol) | Ð 10 | Intrinsic Viscosity (mL/g) | DP 11 |
|---|---|---|---|---|---|---|
| ALCM 1 | C. myriophylloides | 59,000 (±1.3%) | 76,000 (±0.9%) | 1.3 (±1.6%) | 224.2 (±1.5%) | 352 |
| ALSM 2 | S. muticum | 173,000 (±1.7%) | 202,000 (±1.7%) | 1.2 (±2.3%) | 738.6 (±1.0%) | 935 |
| OACM 3 | C. myriophylloides | 11,000 (±4.9%) | 19,000 (±5.6%) | 1.7 (±7.4%) | 35.8 (±6.0%) | 88 |
| OASM 4 | S. muticum | 2000 (±9.8%) | 3000 (±10.8%) | 1.04 (±14.5%) | 8.4 (±3.1%) | 14 |
| FUCCM 5 | C. myriophylloides | 37,000 (±3.0%) | 148,000 (±1.0%) | 4.0 (±3.1%) | 84.4 (±3.7%) | Nd 12 |
| AGP 6 | C. decorticatum | 1,295,000 (±0.9%) | 2,042,000 (±0.7%) | 1.6 (±1.2%) | 560.6 (±4.%) | Nd |
| AGB 7 | C. decorticatum | 711,000 (±1.4%) | 2,173,000 (±0.4%) | 3.1 (±1.4%) | 266.7 (±3.0%) | Nd |
| Monosaccharides (%Molar Ratio) | AGB 1 | AGP 2 | FUCCM 3 | ALCM 4 | ALSM 5 |
|---|---|---|---|---|---|
| Galactose (Gal) | 46.07 ± 1.46 | 51.55 ± 2.67 | Nd | Nd | Nd |
| Glucose (Glc) | 38.78 ± 0.24 | 24.56 ± 1.05 | 0.91 ± 0.10 | 0.55 ± 0.09 | 1.13 ± 0.57 |
| Arabinose (Ara) | 12.08 ± 1 | 17.32 ± 1.33 | Nd | Nd | Nd |
| Xylose (Xyl) | 1.70 ± 0.23 | 3.93 ± 0.61 | Nd | Nd | Nd |
| Rhamnose (Rha) | 1.37 ± 0.13 | 2.63 ± 0.15 | Nd | Nd | Nd |
| Fucose (Fuc) | Nd 6 | Nd | 53.39 ± 4.57 | 24.84 ± 4.74 | 11.28 ± 1.23 |
| Mannuronic acid (ManA) | Nd | Nd | 45.70 ± 4.52 | 74.61 ± 4.81 | 87.59 ± 0.85 |
| Alginates | Source | Frequencies of Structural Blocks | % M | % G | η | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| FG | FM | FGG | FMM | FMG | FGM | M/G | |||||
| ALSM 1 | S. muticum | 0.54 | 0.46 | 0.47 | 0.40 | 0.063 | 0.063 | 0.87 | 46 | 54 | 0.25 |
| ALCM 2 | C. myriophylloides | 0.47 | 0.53 | 0.43 | 0.49 | 0.038 | 0.038 | 1.12 | 53 | 47 | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitouguinane, M.; Alaoui-Talibi, Z.E.; Rchid, H.; Fendri, I.; Abdelkafi, S.; El-Hadj, M.D.O.; Boual, Z.; Dubessay, P.; Michaud, P.; Traïkia, M.; et al. Polysaccharides from Moroccan Green and Brown Seaweed and Their Derivatives Stimulate Natural Defenses in Olive Tree Leaves. Appl. Sci. 2022, 12, 8842. https://doi.org/10.3390/app12178842
Aitouguinane M, Alaoui-Talibi ZE, Rchid H, Fendri I, Abdelkafi S, El-Hadj MDO, Boual Z, Dubessay P, Michaud P, Traïkia M, et al. Polysaccharides from Moroccan Green and Brown Seaweed and Their Derivatives Stimulate Natural Defenses in Olive Tree Leaves. Applied Sciences. 2022; 12(17):8842. https://doi.org/10.3390/app12178842
Chicago/Turabian StyleAitouguinane, Meriem, Zainab El Alaoui-Talibi, Halima Rchid, Imen Fendri, Slim Abdelkafi, Mohamed Didi Ould El-Hadj, Zakaria Boual, Pascal Dubessay, Philippe Michaud, Mounir Traïkia, and et al. 2022. "Polysaccharides from Moroccan Green and Brown Seaweed and Their Derivatives Stimulate Natural Defenses in Olive Tree Leaves" Applied Sciences 12, no. 17: 8842. https://doi.org/10.3390/app12178842