
Assessment of Different Spent Mushroom Substrates to Bioremediate Soils Contaminated with Petroleum Hydrocarbons

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spent Mushroom Substrates
2.2. Contaminated Soil
2.3. Mycoremediation Assay
2.4. Statistical Analysis
3. Results
3.1. Soil Characterization
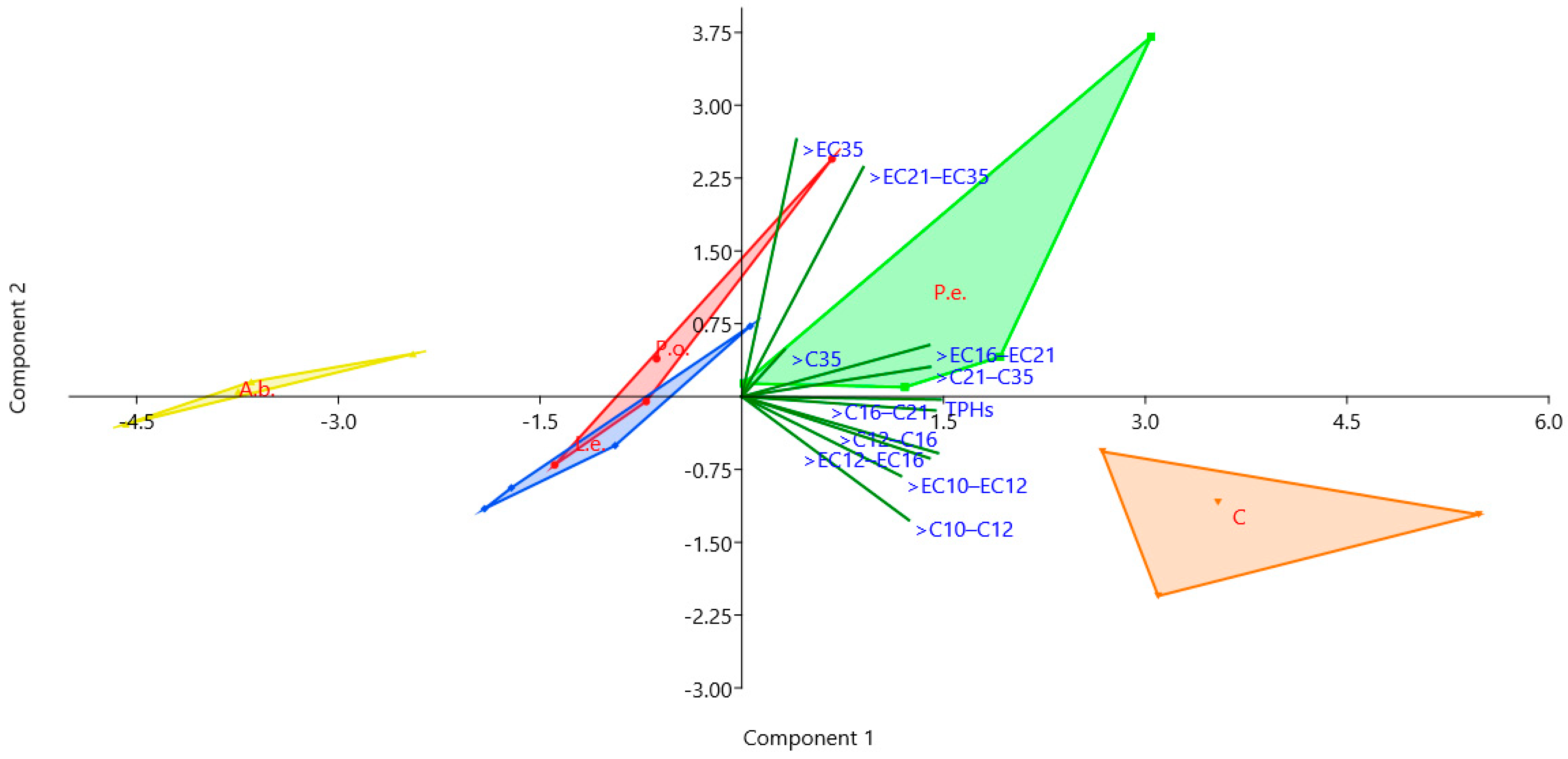
3.2. Mycoremediation Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García-Delgado, C.; Alfaro-Barta, I.; Eymar, E. Combination of biochar amendment and mycoremediation for polycyclic aromatic hydrocarbons immobilization and biodegradation in creosote-contaminated soil. J. Hazard. Mater. 2015, 285, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Aioub, A.A.A.; Li, Y.; Qie, X.; Zhang, X.; Hu, Z. Reduction of soil contamination by cypermethrin residues using phytoremediation with Plantago major and some surfactants. Environ. Sci. Eur. 2019, 31, 26. [Google Scholar] [CrossRef] [Green Version]
- Aioub, A.A.A.; Zuo, Y.; Aioub, A.A.A.; Hu, Z. Biochemical and phytoremediation of Plantago major L. to protect tomato plants from the contamination of cypermethrin pesticide. Environ. Sci. Pollut. Res. 2021, 28, 43992–44001. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, N.M.; Alardhi, S.M.; Mohammed, A.K.; Salih, I.K.; Albayati, T.M. Challenges in the implementation of bioremediation processes in petroleum-contaminated soils: A review. Environ. Nanotechnol. Monit. Manag. 2022, 18, 100694. [Google Scholar] [CrossRef]
- Chukwunonso, I.; Ahmed, A.; Hassan, A.; Shahul, F. Environmental Technology & Innovation Remediation of soil and water contaminated with petroleum hydrocarbon: A review. Environ. Technol. Innov. 2020, 17, 100526. [Google Scholar] [CrossRef]
- Rathankumar, A.K.; Saikia, K.; Kumar, P.S.; Varjani, S.; Kalita, S.; Bharadwaj, N.; George, J.; Vaidyanathan, V.K. Surfactant-aided mycoremediation of soil contaminated with polycyclic aromatic hydrocarbon (PAHs): Progress, limitation, and countermeasures. J. Chem. Technol. Biotechnol. 2022, 97, 391–408. [Google Scholar] [CrossRef]
- D’Annibale, A.; Rosetto, F.; Leonardi, V.; Federici, F.; Petruccioli, M. Role of autochthonous filamentous fungi in bioremediation of a soil historically contaminated with aromatic hydrocarbons. Appl. Environ. Microbiol. 2006, 72, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; Solanas, A.M.; de Lapuente, J.; Borràs, M.; Viñas, M. A diversified approach to evaluate biostimulation and bioaugmentation strategies for heavy-oil-contaminated soil. Sci. Total Environ. 2012, 435–436, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Maletić, S.P.; Dalmacija, B.D.; Rončević, S.D.; Agbaba, J.R.; Perović, S.D.U. Impact of hydrocarbon type, concentration and weathering on its biodegradability in soil. J. Environ. Sci. Heal. Part A Toxic/Hazard. Subst. Environ. Eng. 2011, 46, 1042–1049. [Google Scholar] [CrossRef]
- Treu, R.; Falandysz, J. Mycoremediation of hydrocarbons with basidiomycetes—A review. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2017, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, N.; Mannan, M.A.-u. Mycoremediation: Expunging environmental pollutants. Biotechnol. Rep. 2020, 26, e00452. [Google Scholar] [CrossRef]
- Levasseur, A.; Drula, E.; Lombard, V.; Coutinho, P.M.; Henrissat, B. Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary redox enzymes. Biotechnol. Biofuels 2013, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, K.; Porollo, A.; Lam, Y.W.; Yadav, J.S. A fungal P450 (CYP5136A3) capable of oxidizing polycyclic aromatic hydrocarbons and endocrine disrupting alkylphenols: Role of Trp129 and Leu324. PLoS ONE 2011, 6, e28286. [Google Scholar] [CrossRef] [Green Version]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Cabral, L.; Giovanella, P.; Pellizzer, E.P.; Teramoto, E.H.; Kiang, C.H.; Sette, L.D. Microbial communities in petroleum-contaminated sites: Structure and metabolisms. Chemosphere 2022, 286. [Google Scholar] [CrossRef] [PubMed]
- Gramss, G.; Voigt, K.D.; Kirsche, B. Degradation of polycyclic aromatic hydrocarbons with three to seven aromatic rings by higher fungi in sterile and unsterile soils. Biodegradation 1999, 10, 51–62. [Google Scholar] [CrossRef]
- Zied, D.C.; Sánchez, J.E.; Noble, R.; Pardo-Giménez, A. Use of spent mushroom substrate in new mushroom crops to promote the transition towards a circular economy. Agronomy 2020, 10, 1239. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. Edible Med. Mushrooms 2017, 2010, 5–13. [Google Scholar] [CrossRef]
- Leong, Y.K.; Ma, T.W.; Chang, J.S.; Yang, F.C. Recent advances and future directions on the valorization of spent mushroom substrate (SMS): A review. Bioresour. Technol. 2022, 344, 126157. [Google Scholar] [CrossRef]
- Ghose, A.; Mitra, S. Spent waste from edible mushrooms offers innovative strategies for the remediation of persistent organic micropollutants: A review. Environ. Pollut. 2022, 305, 119285. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Chauhan, P.K. Optimization of eco-friendly amendments as sustainable asset for salt-tolerant plant growth-promoting bacteria mediated maize (Zea mays L.) plant growth, Na uptake reduction and saline soil restoration. Environ. Res. 2022, 211, 113081. [Google Scholar] [CrossRef]
- Frutos, I.; García-Delgado, C.; Gárate, A.; Eymar, E. Biosorption of heavy metals by organic carbon from spent mushroom substrates and their raw materials. Int. J. Environ. Sci. Technol. 2016, 13, 2713–2720. [Google Scholar] [CrossRef]
- García-Delgado, C.; D’Annibale, A.; Pesciaroli, L.; Yunta, F.; Crognale, S.; Petruccioli, M.; Eymar, E. Implications of polluted soil biostimulation and bioaugmentation with spent mushroom substrate (Agaricus bisporus) on the microbial community and polycyclic aromatic hydrocarbons biodegradation. Sci. Total Environ. 2015, 508, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, G.; Becarelli, S.; Lorenzi, R.; Gentini, A.; Di Gregorio, S. PCB in the environment: Bio-based processes for soil decontamination and management of waste from the industrial production of Pleurotus ostreatus. N. Biotechnol. 2017, 39, 232–239. [Google Scholar] [CrossRef]
- Zhou, J.; Ge, W.; Zhang, X.; Wu, J.; Chen, Q.; Ma, D.; Chai, C. Effects of spent mushroom substrate on the dissipation of polycyclic aromatic hydrocarbons in agricultural soil. Chemosphere 2020, 259, 127462. [Google Scholar] [CrossRef] [PubMed]
- García-Delgado, C.; Alonso-Izquierdo, M.; González-Izquierdo, M.; Yunta, F.; Eymar, E. Purification of polluted water with spent mushroom (Agaricus bisporus) substrate: From agricultural waste to biosorbent of phenanthrene, Cd and Pb. Environ. Technol. 2017, 38, 1792–1799. [Google Scholar] [CrossRef]
- Frutos, I.; García-Delgado, C.; Cala, V.; Gárate, A.; Eymar, E. The use of spent mushroom compost to enhance the ability of Atriplex halimus to phytoremediate contaminated mine soils. Environ. Technol. 2017, 38, 1075–1084. [Google Scholar] [CrossRef]
- Carpio, M.J.; Rodríguez-Cruz, M.S.; García-Delgado, C.; Sánchez-Martín, M.J.; Marín-Benito, J.M. Mobility monitoring of two herbicides in amended soils: A field study for modeling applications. J. Environ. Manag. 2020, 260, 110161. [Google Scholar] [CrossRef]
- Carpio, M.J.; Marín-Benito, J.M.; García-Delgado, C.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Soil microbial community changes in a field treatment with chlorotoluron, flufenacet and diflufenican and two organic amendments. Agronomy 2020, 10, 1166. [Google Scholar] [CrossRef]
- Mayans, B.; Camacho-Arévalo, R.; García-Delgado, C.; Alcántara, C.; Nägele, N.; Antón-Herrero, R.; Escolástico, C.; Eymar, E. Mycoremediation of soils polluted with trichloroethylene: First evidence of pleurotus genus effectiveness. Appl. Sci. 2021, 11, 1354. [Google Scholar] [CrossRef]
- Agencia Estatal Boletín Oficial del Estado, Ministerio de la Presidencia. Real Decreto 9/2005, de 14 de enero; Agencia Estatal Boletín Oficial del Estado, Ministerio de la Presidencia: Madrid, Spain, 2005; Volume 2005, pp. 1833–1843. [Google Scholar]
- García-Delgado, C.; Cala, V.; Eymar, E. Influence of chemical and mineralogical properties of organic amendments on the selection of an adequate analytical procedure for trace elements determination. Talanta 2012, 88, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Dawson, J.J.C.; Godsiffe, E.J.; Thompson, I.P.; Ralebitso-Senior, T.K.; Killham, K.S.; Paton, G.I. Application of biological indicators to assess recovery of hydrocarbon impacted soils. Soil Biol. Biochem. 2007, 39, 164–177. [Google Scholar] [CrossRef]
- Adam, G.; Duncan, H. Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 2001, 33, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Barajas-Aceves, M.; Hassan, M.; Tinoco, R.; Vazquez-Duhalt, R. Effect of pollutants on the ergosterol content as indicator of fungal biomass. J. Microbiol. Methods 2002, 50, 227–236. [Google Scholar] [CrossRef]
- Covino, S.; Čvančarová, M.; Muzikář, M.; Svobodová, K.; D’annibale, A.; Petruccioli, M.; Federici, F.; Křesinová, Z.; Cajthaml, T. An efficient PAH-degrading Lentinus (Panus) tigrinus strain: Effect of inoculum formulation and pollutant bioavailability in solid matrices. J. Hazard. Mater. 2010, 183, 669–676. [Google Scholar] [CrossRef] [PubMed]
- García-Delgado, C.; Eymar, E.; Camacho-Arévalo, R.; Petruccioli, M.; Crognale, S.; D’Annibale, A. Degradation of tetracyclines and sulfonamides by stevensite- and biochar-immobilized laccase systems and impact on residual antibiotic activity. J. Chem. Technol. Biotechnol. 2018, 93, 3394–3409. [Google Scholar] [CrossRef]
- Ruiz-Dueñas, F.J.; Morales, M.; García, E.; Miki, Y.; Martínez, M.J.; Martínez, A.T. Substrate oxidation sites in versatile peroxidase and other basidiomycete peroxidases. J. Exp. Bot. 2009, 60, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Pindado, O.; Pérez-Pastor, R.M.; Escolano, O. An analytical method for quantifying petroleum hydrocarbon fractions in soils, and its associated uncertainties. Anal. Methods 2014, 6, 5527–5536. [Google Scholar] [CrossRef]
- Carrasco, J.; García-Delgado, C.; Lavega, R.; Tello, M.L.; De Toro, M.; Barba-Vicente, V.; Rodríguez-Cruz, M.S.; Sánchez-Martín, M.J.; Pérez, M.; Preston, G.M. Holistic assessment of the microbiome dynamics in the substrates used for commercial champignon (Agaricus bisporus) cultivation. Microb. Biotechnol. 2020, 13, 1933–1947. [Google Scholar] [CrossRef]
- Chiu, S.W.; Gao, T.; Chan, C.S.S.; Ho, C.K.M. Removal of spilled petroleum in industrial soils by spent compost of mushroom Pleurotus pulmonarius. Chemosphere 2009, 75, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, S.; Becarelli, S.; Siracusa, G.; Ruffini Castiglione, M.; Petroni, G.; Masini, G.; Gentini, A.; de Lima e Silva, M.R.; Lorenzi, R. Pleurotus ostreatus spent mushroom substrate for the degradation of polycyclic aromatic hydrocarbons: The case study of a pilot dynamic biopile for the decontamination of a historically contaminated soil. J. Chem. Technol. Biotechnol. 2016, 91, 1654–1664. [Google Scholar] [CrossRef]
- Calvo, C.; Rodríguez-Calvo, A.; Robledo-Mahón, T.; Manzanera, M.; González-López, J.; Aranda, E.; Silva-Castro, G.A. Biostimulation of crude oil-polluted soils: Influence of initial physicochemical and biological characteristics of soil. Int. J. Environ. Sci. Technol. 2019, 16, 4925–4934. [Google Scholar] [CrossRef]
- Silva-Castro, G.A.; Uad, I.; Rodríguez-Calvo, A.; González-López, J.; Calvo, C. Response of autochthonous microbiota of diesel polluted soils to land-farming treatments. Environ. Res. 2015, 137, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.A.; Peco, J.D.; De Toro, J.A.; Moreno, C.; Amorós, J.A.; Moreno, M.M.; García-Noguero, E.M.; Higueras, P. Approach to the potential usage of two wood ashes waste as soil amendments on the basis of the dehydrogenase activity and soil oxygen consumption. J. Soils Sediments 2018, 18, 2148–2156. [Google Scholar] [CrossRef]
- Krishnan, A.; Convey, P.; Gonzalez, M.; Smykla, J.; Alias, S.A. Effects of temperature on extracellular hydrolase enzymes from soil microfungi. Polar Biol. 2018, 41, 537–551. [Google Scholar] [CrossRef]
- Becarelli, S.; Siracusa, G.; Chicca, I.; Bernabei, G.; Di Gregorio, S. Ascomycetes versus spent mushroom substrate in mycoremediation of dredged sediments contaminated by total petroleum hydrocarbons: The involvement of the bacterial metabolism. Water 2021, 13, 3040. [Google Scholar] [CrossRef]
- Covino, S.; Stella, T.; D’Annibale, A.; Lladó, S.; Baldrian, P.; Čvančarová, M.; Cajthaml, T.; Petruccioli, M. Comparative assessment of fungal augmentation treatments of a fine-textured and historically oil-contaminated soil. Sci. Total Environ. 2016, 566, 250–259. [Google Scholar] [CrossRef]
- Mohammadi-Sichani, M.; Mazaheri Assadi, M.; Farazmand, A.; Kianirad, M.; Ahadi, A.M.; Hadian-Ghahderijani, H. Ability of Agaricus bisporus, Pleurotus ostreatus and Ganoderma lucidum compost in biodegradation of petroleum hydrocarbon-contaminated soil. Int. J. Environ. Sci. Technol. 2019, 16, 2313–2320. [Google Scholar] [CrossRef]
- Li, Q.; Liu, J.; Gadd, G.M. Fungal bioremediation of soil co-contaminated with petroleum hydrocarbons and toxic metals. Appl. Microbiol. Biotechnol. 2020, 104, 8999–9008. [Google Scholar] [CrossRef]
- García-Delgado, C.; Yunta, F.; Eymar, E. Bioremediation of multi-polluted soil by spent mushroom (Agaricus bisporus) substrate: Polycyclic aromatic hydrocarbons degradation and Pb availability. J. Hazard. Mater. 2015, 300, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.; Sanromán, M.Á.; Moldes, D. Recent developments and applications of immobilized laccase. Biotechnol. Adv. 2013, 31, 1808–1825. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.T. Molecular biology and structure-function of lignin-degrading heme peroxidases. Enzyme Microb. Technol. 2002, 30, 425–444. [Google Scholar] [CrossRef]
- Knop, D.; Levinson, D.; Makovitzki, A.; Agami, A.; Lerer, E.; Mimran, A.; Yarden, O.; Hadar, Y. Limits of versatility of versatile peroxidase. Appl. Environ. Microbiol. 2016, 82, 4070–4080. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.K.; Das, N. Phyto-mycoremediation of benzo[a]pyrene in soil by combining the role of yeast consortium and sunflower plant. J. Environ. Biol. 2018, 39, 261–268. [Google Scholar] [CrossRef]
- Guirado, M.; Garrido-Sanz, D.; Pindado, O.; Rodríguez-Rastrero, M.; Merino-Martín, L.; Sierra, M.J.; Escolano, O.; Rivilla, R.; Millán, R. Effectiveness of biochar application and bioaugmentation techniques for the remediation of freshly and aged diesel-polluted soils. Int. Biodeterior. Biodegrad. 2021, 163. [Google Scholar] [CrossRef]
- Wu, M.; Xu, Y.; Ding, W.; Li, Y.; Xu, H. Mycoremediation of manganese and phenanthrene by Pleurotus eryngii mycelium enhanced by Tween 80 and saponin. Appl. Microbiol. Biotechnol. 2016, 100, 7249–7261. [Google Scholar] [CrossRef] [PubMed]
- Giubilei, M.A.; Leonardi, V.; Federici, E.; Covino, S.; Šašek, V.; Novotny, C.; Federici, F.; D’Annibale, A.; Petruccioli, M. Effect of mobilizing agents on mycoremediation and impact on the indigenous microbiota. J. Chem. Technol. Biotechnol. 2009, 84, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, C.J. Mycoremediation (bioremediation with fungi)-growing mushrooms to clean the earth. Chem. Speciat. Bioavailab. 2014, 26, 196–198. [Google Scholar] [CrossRef]
- Dickson, U.J.; Coffey, M.; Mortimer, R.J.G.; Di Bonito, M.; Ray, N. Mycoremediation of petroleum contaminated soils: Progress, prospects and perspectives. Environ. Sci. Process. Impacts 2019, 21, 1446–1458. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, E.J.; Rene, E.R.; Gerlach, R. Potential use of fungal-bacterial co-cultures for the removal of organic pollutants. Crit. Rev. Biotechnol. 2021, 42, 361–383. [Google Scholar] [CrossRef] [PubMed]
- Chun, S.C.; Muthu, M.; Hasan, N.; Tasneem, S.; Gopal, J. Mycoremediation of PCBs by pleurotus ostreatus: Possibilities and prospects. Appl. Sci. 2019, 9, 4185. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Choi, I.G. Genomic and transcriptomic perspectives on mycoremediation of polycyclic aromatic hydrocarbons. Appl. Microbiol. Biotechnol. 2020, 104, 6919–6928. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| MHWC (%) | 13 ± 4 | |
| pH (H2O) | 8.66 ± 0.05 | |
| pH (KCl) | 7.42 ± 0.04 | |
| Electric Conductivity (dS/cm) | 0.7 ± 0.2 | |
| Total Limestone | n.d. | |
| Sand (%) | 91.20 ± 0.01 | |
| Silt (%) | 2.5 ± 0.3 | |
| Clay (%) | 6.3 ± 0.3 | |
| Texture | Sandy | |
| Organic Matter (g/kg) | 23 ± 4 | |
| Assimilable Phosphorus (g/kg) | 0.011 ± 0.001 | |
| Nitrogen (g/kg) | 0.2 ± 0.0 | |
| Trace elements (mg/kg) | Legislated values (mg/kg) * | |
| Sb | 4 ± 2 | 80 |
| As | 16 ± 4 | 40 |
| Cd | 0.3 ± 0.0 | 300 |
| Cu | 315 ± 92 | 8000 |
| Co | 4.0 ± 0.2 | 1500 |
| Cr | n.d. | 2300 |
| Sn | 6.75 ± 0.07 | 100,000 |
| Hg | 0.285 ± 0.007 | 15 |
| Mo | 8 ± 1 | 1500 |
| Ni | 8.0 ± 0.9 | 15,600 |
| Pb | 80.5 ± 22 | 2700 |
| Zn | 290 ± 42 | 100,000 |
| Hydrolase (µmol·h−1·g−1) | Dehydrogenase (nmol·h−1·g−1) | Ergosterol (mg/kg) | Lacasse (U/kg) | Versatile Peroxidase (U/kg) | Mn-Peroxidase (U/kg) | |
|---|---|---|---|---|---|---|
| C | 22.5 ± 0.7 b | 0.80 ± 0.08 b | n.d. | n.d. | 70 ± 14 a | 4 ± 4 a |
| Pe | 71 ± 44 b | 2 ± 1 ab | 1.0 ± 0.5 a | 10 ± 7 b | 99 ± 30 a | 17 ± 12 a |
| Le | 76 ± 56 b | 1.0 ± 0.4 b | 0.7 ± 0.5 a | n.d. | 63 ± 50 a | 9 ± 8 a |
| Po | 26 ± 7 b | 1.3 ± 0.5 ab | 0.6 ± 0.7 a | n.d. | 74 ± 57 a | 10 ± 10 a |
| Ab | 378 ± 243 a | 3 ± 2 a | 0.7 ± 0.4 a | 54 ± 38 a | 48 ± 39 a | 10 ± 6 a |
| mg/kg | C | Pe | Le | Po | Ab |
|---|---|---|---|---|---|
| Aliphatic | |||||
| >C10−C12 | 359 ± 44 a | 141 ± 51 b | 190 ± 31 b | 166 ± 12 b | 73 ± 35 c |
| >C12−C16 | 4629 ± 269 a | 3599 ± 283 b | 2902 ± 206 c | 2960 ± 114 c | 2071 ± 343 d |
| >C16−C21 | 5854 ± 245 a | 5180 ± 250 b | 3951 ± 253 c | 4100 ± 248 c | 3144 ± 503 d |
| >C21−C35 | 3204 ± 158 a | 2916 ± 212 ab | 2334 ± 130 cd | 2611 ± 392 bc | 2093 ± 278 d |
| >C35 | 30 ± 9 ab | 22 ± 13 ab | 15 ± 5 b | 39 ± 16 a | 27 ± 10 ab |
| Σ Aliphatic | 14,077 ± 656 a | 11,857 ± 617 b | 9392 ± 599 c | 9875 ± 728 c | 7407 ± 1151 d |
| Aromatic | |||||
| >EC10−EC12 | 18 ± 7 a | 11 ± 2 b | 12 ± 3 ab | 8 ± 2 bc | 4 ± 1 c |
| >EC12−EC16 | 638 ± 104 a | 451 ± 65 b | 294 ± 98 c | 345 ± 36 bc | 127 ± 26 d |
| >EC16−EC21 | 2667 ± 264 a | 2765 ± 298 a | 1811 ± 187 b | 1990 ± 98 b | 1255 ± 111 c |
| >EC21−EC35 | 516 ± 44 ab | 614 ± 180 a | 412 ± 49 b | 522 ± 118 ab | 413 ± 31 b |
| >EC35 | 56 ± 11 a | 85 ± 42 a | 72 ± 39 a | 71 ± 25 a | 58 ± 10 a |
| Σ Aromatic | 3895 ± 410 a | 3926 ± 540 a | 2601 ± 249 b | 2935 ± 223 b | 1857 ± 155 c |
| ΣTPHs | 17,971 ± 1061 a | 15,784 ± 1061 b | 11,993 ± 836 c | 12,811 ± 948 c | 9264 ± 1230 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antón-Herrero, R.; García-Delgado, C.; Baena, N.; Mayans, B.; Delgado-Moreno, L.; Eymar, E. Assessment of Different Spent Mushroom Substrates to Bioremediate Soils Contaminated with Petroleum Hydrocarbons. Appl. Sci. 2022, 12, 7720. https://doi.org/10.3390/app12157720
Antón-Herrero R, García-Delgado C, Baena N, Mayans B, Delgado-Moreno L, Eymar E. Assessment of Different Spent Mushroom Substrates to Bioremediate Soils Contaminated with Petroleum Hydrocarbons. Applied Sciences. 2022; 12(15):7720. https://doi.org/10.3390/app12157720
Chicago/Turabian StyleAntón-Herrero, Rafael, Carlos García-Delgado, Natalia Baena, Begoña Mayans, Laura Delgado-Moreno, and Enrique Eymar. 2022. "Assessment of Different Spent Mushroom Substrates to Bioremediate Soils Contaminated with Petroleum Hydrocarbons" Applied Sciences 12, no. 15: 7720. https://doi.org/10.3390/app12157720