
Preparation of Human Muscle Precursor Cells for the MyoGravity Project’s Study of Cell Cultures in Experiment Units for Space Flight Purposes

 ,
,  and
and
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Muscle Precursor Cells (huMPCs)
2.2. Cell Culture
2.3. Experimental Hardware (EHs)
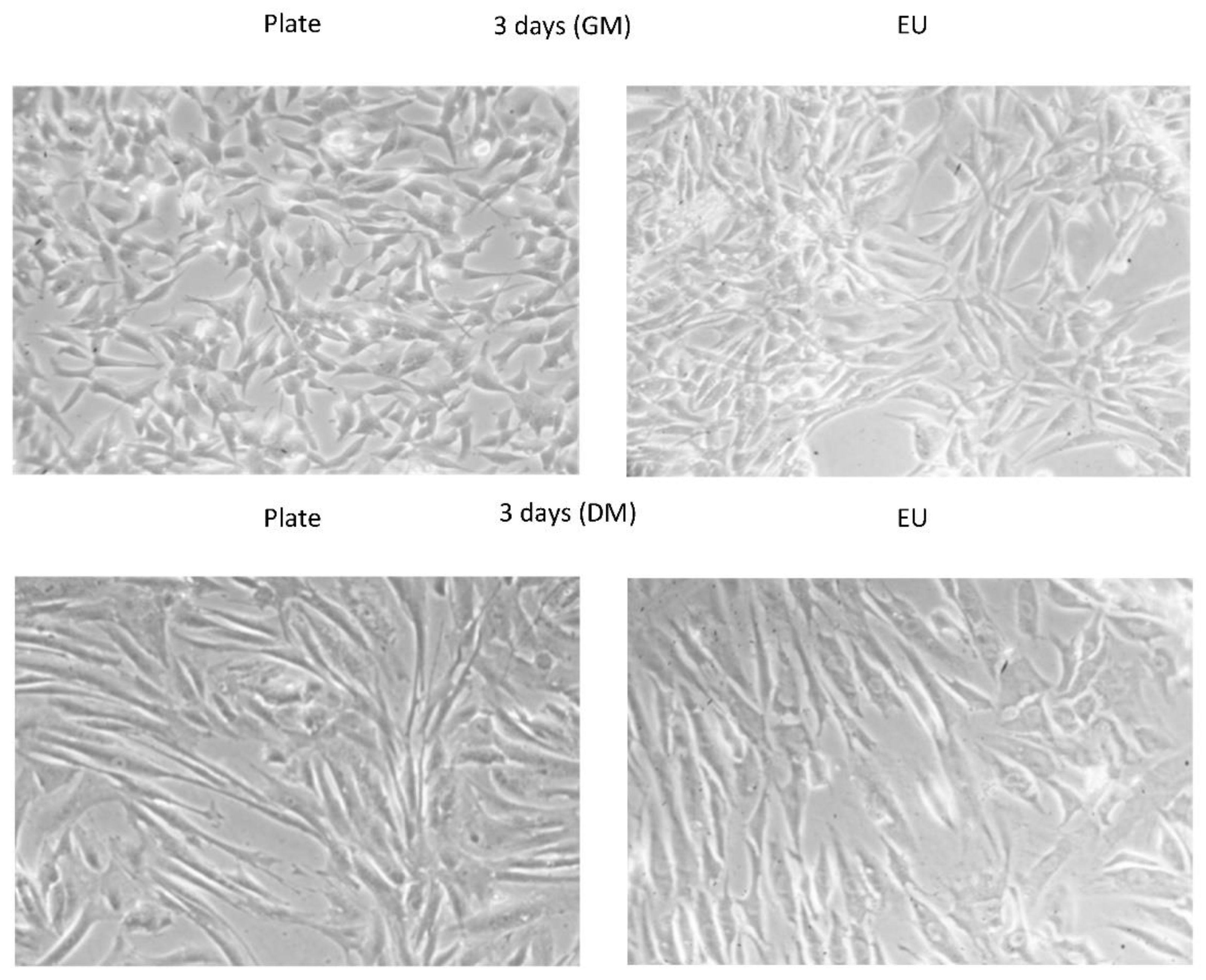
2.4. Cell Growth Inside the Experiment Units
2.5. RNA Extraction and Quantification
2.6. Quantitative Real-Time PCR for Myogenic Regulatory Factor
2.7. MicroRNA Expression Profile
2.8. Statistical Analysis
3. Results
3.1. Cell Culture Optimization into the EUs
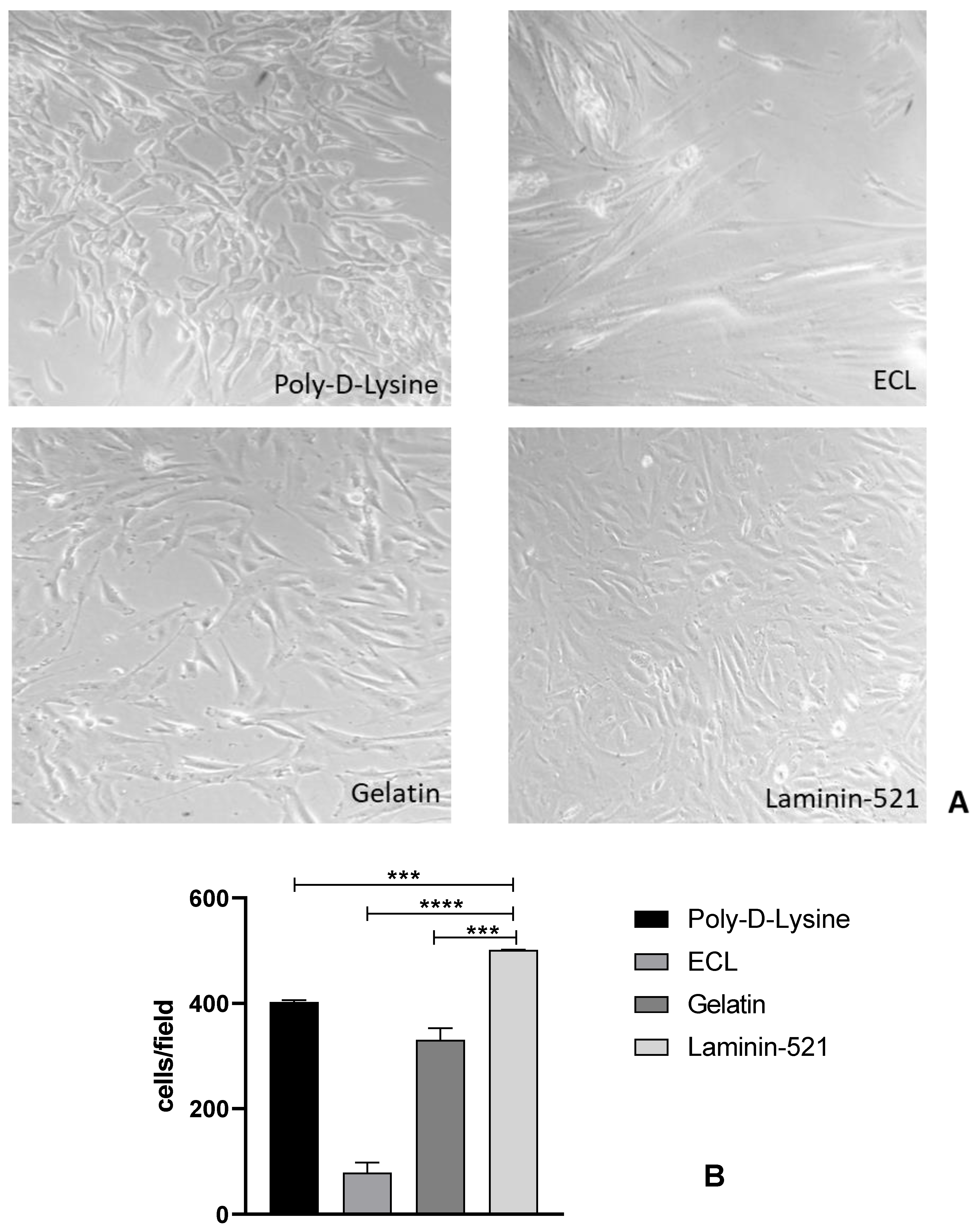
3.1.1. Type of Coating
3.1.2. HEPES Concentration
3.1.3. Cell Resilience to Temperature
3.1.4. Cell Density to Plate
3.1.5. RNA Extraction
3.2. Expression Profile of Muscle-Specific Factors and MicroRNAs in the EUs
3.2.1. Expression Profile of Muscle-Specific Factors
3.2.2. Expression of myomiRNAs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fitts, R.H.; Riley, D.R.; Widrick, J.J. Physiology of a Microgravity Environment Invited Review: Microgravity and Skeletal Muscle. J. Appl. Physiol. 2000, 89, 823–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms Regulating Skeletal Muscle Growth and Atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef] [PubMed]
- Ikemoto, M.; Nikawa, T.; Kano, M.; Hirasaka, K.; Kitano, T.; Watanabe, C.; Tanaka, R.; Yamamoto, T.; Kamada, M.; Kishi, K. Cysteine Supplementation Prevents Unweighting-Induced Ubiquitination in Association with Redox Regulation in Rat Skeletal Muscle. Biol. Chem. 2002, 383, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Hirasaka, K.; Nikawa, T.; Yuge, L.; Ishihara, I.; Higashibata, A.; Ishioka, N.; Okubo, A.; Miyashita, T.; Suzue, N.; Ogawa, T.; et al. Clinorotation Prevents Differentiation of Rat Myoblastic L6 Cells in Association with Reduced NF-ΚB Signaling. Biochim. Biophys. Acta Mol. Cell Res. 2005, 1743, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Battista, N.; Meloni, M.A.; Bari, M.; Mastrangelo, N.; Galleri, G.; Rapino, C.; Dainese, E.; Agrò, A.F.; Pippia, P.; Maccarrone, M. 5-Lipoxygenase-dependent Apoptosis of Human Lymphocytes in the International Space Station: Data from the ROALD Experiment. FASEB J. 2012, 26, 1791–1798. [Google Scholar] [CrossRef]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.M.; Bradamante, S. The Challenging Environment on Board the International Space Station Affects Endothelial Cell Function by Triggering Oxidative Stress through Thioredoxin Interacting Protein Overexpression: The ESA-SPHINX Experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef]
- Cristofaro, F.; Pani, G.; Pascucci, B.; Mariani, A.; Balsamo, M.; Donati, A.; Mascetti, G.; Rea, G.; Rizzo, A.M.; Visai, L. The NATO Project: Nanoparticle-Based Countermeasures for Microgravity-Induced Osteoporosis. Sci. Rep. 2019, 9, 17141. [Google Scholar] [CrossRef]
- Pietrangelo Tiny Percutaneous Needle Biopsy: An Efficient Method for Studying Cellular and Molecular Aspects of Skeletal Muscle in Humans. Int. J. Mol. Med. 2011, 27, 361–367. [CrossRef] [Green Version]
- Fulle, S.; Didonna, S.; Puglielli, C.; Pietrangelo, T.; Beccafico, S.; Bellomo, R.; Protasi, F.; Fano, G. Age-Dependent Imbalance of the Antioxidative System in Human Satellite Cells. Exp. Gerontol. 2005, 40, 189–197. [Google Scholar] [CrossRef]
- Pietrangelo, T.; Puglielli, C.; Mancinelli, R.; Beccafico, S.; Fanò, G.; Fulle, S. Molecular Basis of the Myogenic Profile of Aged Human Skeletal Muscle Satellite Cells during Differentiation. Exp. Gerontol. 2009, 44, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trimarchi, F.; Favaloro, A.; Fulle, S.; Magaudda, L.; Puglielli, C.; Di Mauro, D. Culture of Human Skeletal Muscle Myoblasts: Timing Appearance and Localization of Dystrophin-Glycoprotein Complex and Vinculin-Talin-Integrin Complex. Cells Tissues Organs 2006, 183, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, A.; Moscheni, C.; Maier, J.A.; Castiglioni, S. Culture of Human Cells in Experimental Units for Spaceflight Impacts on Their Behavior. Exp. Biol. Med. 2017, 242, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulle, S.; Sancilio, S.; Mancinelli, R.; Gatta, V.; Di Pietro, R. Dual Role of the Caspase Enzymes in Satellite Cells from Aged and Young Subjects. Cell Death Dis. 2013, 4, e955. [Google Scholar] [CrossRef] [Green Version]
- Di Filippo, E.S.; Mancinelli, R.; Pietrangelo, T.; La Rovere, R.M.L.; Quattrocelli, M.; Sampaolesi, M.; Fulle, S. Myomir Dysregulation and Reactive Oxygen Species in Aged Human Satellite Cells. Biochem. Biophys. Res. Commun. 2016, 473, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Iaculli, F.; Di Filippo, E.S.; Piattelli, A.; Mancinelli, R.; Fulle, S. Dental Pulp Stem Cells Grown on Dental Implant Titanium Surfaces: An in Vitro Evaluation of Differentiation and MicroRNAs Expression: MicroRNAS expression during osteoblasts differentiation. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 953–965. [Google Scholar] [CrossRef]
- Balsamo, M.; Barravecchia, I.; Mariotti, S.; Merenda, A.; De Cesari, C.; Vukich, M.; Angeloni, D. Molecular and Cellular Characterization of Space Flight Effects on Microvascular Endothelial Cell Function-PreparatoryWork for the SFEF Project. Microgravity Sci. Technol. 2014, 26, 351–363. [Google Scholar] [CrossRef]
- Lulli, M.; Cialdai, F.; Vignali, L.; Monici, M.; Luzzi, S.; Cicconi, A.; Cacchione, S.; Magi, A.; Di Gesualdo, F.; Balsamo, M.; et al. The Coenzyme Q10 (CoQ10) as Countermeasure for Retinal Damage Onboard the International Space Station: The CORM Project. Microgravity Sci. Technol. 2018, 30, 925–931. [Google Scholar] [CrossRef]
- Cialdai, F.; Bolognini, D.; Vignali, L.; Iannotti, N.; Cacchione, S.; Magi, A.; Balsamo, M.; Vukich, M.; Neri, G.; Donati, A.; et al. Effect of Space Flight on the Behavior of Human Retinal Pigment Epithelial ARPE-19 Cells and Evaluation of Coenzyme Q10 Treatment. Cell. Mol. Life Sci. 2021, 78, 7795–7812. [Google Scholar] [CrossRef]
- Lee, P.H.U.; Chung, M.; Ren, Z.; Mair, D.B.; Kim, D.-H. Factors Mediating Spaceflight-Induced Skeletal Muscle Atrophy. Am. J. Physiol.-Cell Physiol. 2022, 322, C567–C580. [Google Scholar] [CrossRef]
- Shenkman, B.S. From Slow to Fast: Hypogravity-Induced Remodeling of Muscle Fiber Myosin Phenotype. Acta Nat. 2016, 8, 47–59. [Google Scholar] [CrossRef]
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human Pathophysiological Adaptations to the Space Environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Downs, M.; Martin, D.S.; Hougland, E.; Sarmiento, L.; Arzeno, N.; Pettit, D.R.; Ploutz-Snyder, R.; Cunningham, D.; Jones, L.W.; et al. Teleguided Self-Ultrasound Scanning for Longitudinal Monitoring of Muscle Mass during Spaceflight. iScience 2021, 24, 102344. [Google Scholar] [CrossRef] [PubMed]
- Rittweger, J.; Albracht, K.; Flück, M.; Ruoss, S.; Brocca, L.; Longa, E.; Moriggi, M.; Seynnes, O.; Di Giulio, I.; Tenori, L.; et al. Sarcolab Pilot Study into Skeletal Muscle’s Adaptation to Long-Term Spaceflight. NPJ Microgravity 2018, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- English, K.L.; Downs, M.; Goetchius, E.; Buxton, R.; Ryder, J.W.; Ploutz-Snyder, R.; Guilliams, M.; Scott, J.M.; Ploutz-Snyder, L.L. High Intensity Training during Spaceflight: Results from the NASA Sprint Study. NPJ Microgravity 2020, 6, 21. [Google Scholar] [CrossRef]
- Trappe, S.; Costill, D.; Gallagher, P.; Creer, A.; Peters, J.R.; Evans, H.; Riley, D.A.; Fitts, R.H. Exercise in Space: Human Skeletal Muscle after 6 Months Aboard the International Space Station. J. Appl. Physiol. 2009, 106, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Vunjak-Novakovic, G.; Searby, N.; De Luis, J.; Freed, L.E. Microgravity Studies of Cells and Tissues. Ann. N. Y. Acad. Sci. 2002, 974, 504–517. [Google Scholar] [CrossRef]
- Morabito, C.; Lanuti, P.; Caprara, G.A.; Marchisio, M.; Bizzarri, M.; Guarnieri, S.; Mariggiò, M.A. Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Conditions. Int. J. Mol. Sci. 2019, 20, 1892. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HEPES (mM) | 20 | 25 | 40 |
|---|---|---|---|
| Cell survival (%) | 98.3 ± 3.2 | 95.1 ± 3.5 | 70.7 ± 2.6 |
| Cell proliferation (%) | 97.5 ± 3.9 | 83.4 ± 4.2 | 25.2 ± 3.7 |
| Temperature (°C) | 25 | 27 | 30 |
|---|---|---|---|
| Cell survival (% of control) | 30.5 ± 4.2 | 98.2 ± 6.2 | 98.6 ± 6.5 |
| Conditions | Plate | EU |
|---|---|---|
| PDL | 4.0 ± 0.2 | 3.8 ± 0.1 |
| FI (%) | 55.2 ± 5.1 | 57.0 ± 2.3 |
| RNA Isolation Kit | Yield on Plates ng/µL (260/280 Ratio) | Yield inside the EUs ng/µL (260/280 Ratio) |
|---|---|---|
| mirVana miRNA | total RNA 18.6 (2.05) | total RNA 29.5 (1.54) |
| PureLink miRNA PureLink RNA Mini Kit | microRNAs 3.0 (1.96) total RNA 16.8 (2.06) | microRNAs 3.2 (1.90) total RNA 18.9 (2.13) |
| miRNeasy Micro Kit | total RNA 72.5 (1.98) | total RNA 78.1 (1.90) |
| Coating of Thermanox™ | Human Recombinant Laminin-521 (BioLamina) |
| HEPES concentration | 20 mM |
| Lowest temperature tolerated | 27 °C |
| Plating densities for proliferation purposes | 2.0 × 104 cells per coverslip |
| Plating densities for differentiation purposes | 4.0 × 104 cells per coverslip |
| RNA extraction | miRNeasy Micro Kit (Qiagen) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Filippo, E.S.; Chiappalupi, S.; Balsamo, M.; Vukich, M.; Sorci, G.; Fulle, S. Preparation of Human Muscle Precursor Cells for the MyoGravity Project’s Study of Cell Cultures in Experiment Units for Space Flight Purposes. Appl. Sci. 2022, 12, 7013. https://doi.org/10.3390/app12147013
Di Filippo ES, Chiappalupi S, Balsamo M, Vukich M, Sorci G, Fulle S. Preparation of Human Muscle Precursor Cells for the MyoGravity Project’s Study of Cell Cultures in Experiment Units for Space Flight Purposes. Applied Sciences. 2022; 12(14):7013. https://doi.org/10.3390/app12147013
Chicago/Turabian StyleDi Filippo, Ester Sara, Sara Chiappalupi, Michele Balsamo, Marco Vukich, Guglielmo Sorci, and Stefania Fulle. 2022. "Preparation of Human Muscle Precursor Cells for the MyoGravity Project’s Study of Cell Cultures in Experiment Units for Space Flight Purposes" Applied Sciences 12, no. 14: 7013. https://doi.org/10.3390/app12147013