
The Effect of UV-C Irradiation on the Mechanical and Physiological Properties of Potato Tuber and Different Products


Abstract
:1. Introduction
2. Influence of UV-C on the Physical and Mechanical Properties of Different Crops
2.1. Physical Properties
2.2. Mechanical Properties
3. Effect of UV-C on the Mechanical Properties of Potato
4. Stimulation Process of UV-C
4.1. Effect of UV-C on Cell Physiology and Mechanical Properties
4.2. Low UV-C Dose Stimulation
4.3. Moderate and High UV-C Dose Stimulation
5. Factors Affecting UV-C Process in Stimulation
6. Innovative Technologies Used in Combination with UV-C
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soares, I.G.; Silva, E.B.; Amaral, A.J.; Machado, E.C.; Silva, J.M. Physico-chemical and sensory evaluation of potato (Solanum tuberosum L.) after irradiation. An. Acad. Bras. Ciências 2016, 88, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawara, P.; Gliniak, M.; Popardowski, E.; Szczuka, M.; Trzyniec, K. Control system of a prototype measurement system for the identification of ultra-low photonic emission of organic materials. In Proceedings of the 2018 Progress in Applied Electrical Engineering, Koscielisko, Poland, 18–22 June 2018. [Google Scholar] [CrossRef]
- Jakubowski, T. Transfer of microwave irradiation effects of seed potatoes (Solanum tuberosum L.) to the plants of next generations. Bulg. J. Agric. Sci. 2015, 21, 1185–1193. [Google Scholar]
- Kielbasa, P.; Drozdz, T.; Nawara, P.; Drozdz, M.; Trzyniec, K. Assessment of the potential of using photon emission to identify selected qualitative features of organic matter. In Proceedings of the 2018 Applications of Electromagnetics in Modern Techniques and Medicine, Racławice, Poland, 9–12 September 2018; pp. 117–120. [Google Scholar] [CrossRef]
- Nawara, P.; Trzyniec, K.; Dróżdż, T.; Popardowski, E.; Juliszewski, T.; Zagórda, M.; Miernik, A. Analysis of the possibility of identifying the quality parameters of the oil using ultra-weak secondary luminescence. Prz. Elektrotech. 2020, 96, 117–120. (In Polish) [Google Scholar] [CrossRef]
- Jakubowski, T. The influence of selected physical methods on the content of starch and simple sugars in stored potato tubers. In Proceedings of the 2019 Applications of Electromagnetics in Modern Engineering and Medicine, Janow Podlaski, Poland, 9–12 June 2019; pp. 63–66. [Google Scholar] [CrossRef]
- Jakubowski, T. The reaction of garden cress (Lepidium sativum L. to microwave radiation. In Proceedings of the 2018 Applications of Electromagnetics in Modern Techniques and Medicine, Racławice, Poland, 9–12 September 2018; pp. 81–84. [Google Scholar] [CrossRef]
- Fauster, T.; Schlossnikl, D.; Rath, F.; Ostermeier, R.; Teufel, F.; Toepfl, S.; Jaeger, H. Impact of pulsed electric field (PEF) pretreatment on process performance of industrial French fries production. J. Food Eng. 2018, 235, 16–22. [Google Scholar] [CrossRef]
- Jadhav, H.B.; Annapure, U.S.; Deshmukh, R.R. Non-thermal technologies for food processing. Front. Nutr. 2021, 8, 657090. [Google Scholar] [CrossRef]
- Huang, H.W.; Wu, S.J.; Lu, J.K.; Shyu, Y.T.; Wang, C.Y. Current status and future trends of high-pressure processing in food industry. Food Control 2017, 72, 1–8. [Google Scholar] [CrossRef]
- Jakubowski, T. The effect of stimulation of seed potatoes (Solanum tuberosum L.) in the magnetic field on selected vegetation parameters of potato plants. Prz. Elektrotech. 2020, 96, 166–169. [Google Scholar]
- Jakubowski, T. The influence of microwave radiation at the frequency 2.45 GHz on the germination. Prz. Elektrotech. 2018, 94, 254–325. [Google Scholar] [CrossRef]
- Jakubowski, T. Effect of microwave radiation on the germination of Solanum tuberosum L. tubers. Bangladesh J. Bot. 2016, 45, 1255–1257. [Google Scholar]
- Mahto, R.; Das, M. Effect of gamma irradiation on the physico-mechanical and chemical properties of potato (Solanum tuberosum L.), cv. ‘Kufri Sindhuri’, in non-refrigerated storage conditions. Postharvest Biol. Technol. 2014, 92, 37–45. [Google Scholar] [CrossRef]
- Jamieson, L.E.; Meier, X.; Page, B.; Zulhendri, F.; Page-Weir, N.; Brash, D.; McDonald, R.M.; Stanley, J.; Woolf, A.B. A review of postharvest disinfestation technologies for selected fruits and vegetables. Plant Food Res. Client Rep. 2009, 19, 36072. [Google Scholar]
- Lu, C.; Ding, J.; Park, H.K.; Feng, H. High intensity ultrasound as a physical elicitor affects secondary metabolites and antioxidant capacity of tomato fruits. Food Control 2020, 113, 107176. [Google Scholar] [CrossRef]
- Hockberger, P.E. A History of Ultraviolet Photobiology for Humans, Animals and Microorganisms. Photochem. Photobiol. 2002, 76, 561–579. [Google Scholar] [CrossRef]
- Bintsis, T.; Litopoulou-Tzanetaki, E.; Robinson, R.K. Existing and potential applications of ultraviolet light in the food industry–a critical review. J. Sci. Food Agric. 2000, 80, 637–645. [Google Scholar] [CrossRef]
- Darras, A.I.; Tsikaloudakis, G.; Lycoskoufis, I.; Dimitriadis, C.; Karamousantas, D. Low doses of UV-C irradiation affects growth, fruit yield and photosynthetic activity of tomato plants. Sci. Hortic. 2020, 267, 109357. [Google Scholar] [CrossRef]
- Vanhaelewyn, L.; Van Der Straeten, D.; De Coninck, B.; Vandenbussche, F. Ultraviolet radiation from a plant perspective: The plant-microorganism context. Front. Plant Sci. 2020, 11, 597642. [Google Scholar] [CrossRef]
- Rameš, J.; Chaloupecký, V.; Sojková, N.; Bencko, V. An attempt to demonstrate the increased resistance of selected bacterial strains during repeated exposure to UV radiation at 254 nm. Cent. Eur. J. Public Health 1997, 5, 30–31. [Google Scholar]
- United States Food and Drug Administration—FDA. Ultraviolet radiation for the processing and treatment of food. In Code of Federal Regulations; 21 Part, Section 179.39; FDA: Silver Spring, MD, USA, 2002. [Google Scholar]
- National Archives and Records Administration, Code of Federal Regulations. 21 CFR 179.39—Ultraviolet Radiation for the Processing and Treatment of Food. Available online: https://www.govinfo.gov/app/details/CFR-2011-title21-vol3/CFR-2011-title21-vol3-sec179-39/context (accessed on 4 May 2022).
- Frohnmeyer, H.; Staiger, D. Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant Physiol. 2003, 133, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Bassham, J.A.; Calvin, M. The path of carbon in photosynthesis. In Die CO2-Assimilation. The Assimilation of Carbon Dioxide. Handbuch der Pflanzenphysiologie; Pirson, A., Ed.; Encyclopedia of Plant Physiology; Springer: Berlin/Heidelberg, Germany, 1960; p. 5. [Google Scholar] [CrossRef] [Green Version]
- Konstankiewicz, K.; Pawlak, K.; Zdunek, A. Influence_of_structural_parameters. Int. Agrophys. 2000, 15, 243–246. [Google Scholar]
- Pombo, M.A.; Dotto, M.C.; Martínez, G.A.; Civello, P.M. UV-C irradiation delays strawberry fruit softening and modifies the expression of genes involved in cell wall degradation. Postharvest Biol. Technol. 2009, 51, 141–148. [Google Scholar] [CrossRef]
- Jackman, R.L.; Stanley, D.W. Perspectives in the textural evaluation of plant foods. Trends Food Sci. Technol. 1995, 6, 187–194. [Google Scholar] [CrossRef]
- Waldron, K.W.; Smith, A.C.; Parr, A.J.; Ng, A.; Parker, M.L. New approaches to understanding and controlling cell separation in relation to fruit and vegetable texture. Trends Food Sci. Technol. 1997, 8, 213–221. [Google Scholar] [CrossRef]
- Ichiki, H.; Van, N.N.; Yoshinaga, K. Stone-clod Separation and Its Application to Potato Cultivation in Hokkaido. Eng. Agric. Environ. Food 2013, 6, 77–85. [Google Scholar] [CrossRef]
- Zhang, D.Q.; Mu, T.H.; Sun, H.N.; Chen, J.W.; Zhang, M. Comparative study of potato protein concentrates extracted using ammonium sulfate and isoelectric precipitation. Int. J. Food Prop. 2017, 20, 2113–2127. [Google Scholar] [CrossRef] [Green Version]
- Ezekiel, R.; Singh, N.; Sharma, S.; Kaur, A. Beneficial phytochemicals in potato—A review. Food Res. Int. 2013, 50, 487–496. [Google Scholar] [CrossRef]
- Flegg, P.B.; Spencer, D.M.; Wood, D.A. The Biology and Technology of the Cultivated Mushroom; John Wiley & Sons Ltd.: Chichester, UK, 1985; pp. i–xii+347. [Google Scholar]
- Miernik, A.; Jakubowski, T. Selected methods for starch content determination in plant materials. J. Phys. Conf. Ser. 2021, 1782, 012019. [Google Scholar] [CrossRef]
- Nawara, P.; Jakubowski, T.; Sobol, Z. Application of the CIE L∗a∗b∗ method for the evaluation of the colour of fried products from potato tubers exposed to C band ultraviolet light. E3S Web Conf. 2019, 132, 02004. [Google Scholar] [CrossRef] [Green Version]
- Błaszczak, W.; Sadowska, J.; Fornal, J.; Vacek, J.; Flis, B.; Zagórski-Ostoja, W. Influence of cooking and microwave heating on microstructure and mechanical properties of transgenic potatoes. Food/Nahrung 2004, 48, 169–176. [Google Scholar] [CrossRef]
- Jakubowski, T. A system for the control and recording of physical parameters inside a chamber for UV-C irradiating of biological material. E3S Web Conf. 2019, 132, 01006. [Google Scholar] [CrossRef] [Green Version]
- Gliniak, M.; Tomasik, M.; Popardowski, E.; Knaga, J.; Lis, A.; Gliniak, M. Application of natural luminescence for analysis of the radionuclide migration path during hard coal combustion. In Proceedings of the 2018 Applications of Electromagnetics in Modern Techniques and Medicine, Racławice, Poland, 9–12 September 2018; pp. 61–64. [Google Scholar] [CrossRef]
- Kharchenko, S.; Borshch, Y.; Kovalyshyn, S.; Piven, M.; Abduev, M.; Miernik, A.; Popardowski, E.; Kiełbasa, P. Modeling of aerodynamic separation of preliminarily stratified grain mixture in vertical pneumatic separation duct. Appl. Sci. 2021, 11, 4383. [Google Scholar] [CrossRef]
- Jakubowski, T.; Syrotyuk, S.; Yankovska, K. The use of microwave radiation with a frequency of 2.45 GHz as a factor reducing the storage losses of potato tubers. J. Phys. Conf. Ser. 2021, 1782, 012011. [Google Scholar] [CrossRef]
- Gliniak, M.; Tomasik, M.; Popardowski, E.; Knaga, J.; Lis, A.; Gliniak, M. Application of natural luminescence for assessment of hard coal quality. In Proceedings of the 2018 Applications of Electromagnetics in Modern Techniques and Medicine, Racławice, Poland, 9–12 September 2018; pp. 73–76. [Google Scholar] [CrossRef]
- Sobol, Z.; Jakubowski, T.; Nawara, P. The effect of UV-C stimulation of potato tubers and soaking of potato strips in water on color and analyzed color by CIE l*a*b*. Sustainability 2020, 12, 3487, Correction in Sustainability 2020, 12, 7473. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Fan, D.; Huang, L.; Gao, Y.; Lian, H.; Zhao, J.; Zhang, H. Effects of microwaves on molecular arrangements in potato starch. RSC Adv. 2017, 7, 14348–14353. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Yan, M.; Yuan, S.; Sun, S.; Huo, Q. Effect of microwave treatment on the physicochemical properties of potato starch granules. Chem. Cent. J. 2013, 7, 113. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, B.S.; Inamura, P.Y.; Mastro, N.L. The influence of gamma irradiation on texture, color and viscosity properties of potato starch. In Proceedings of the 2015 International Nuclear Atlantic Conference—INAC 2015, São Paulo, Brazil, 4–9 October 2015. [Google Scholar]
- Wang, J.; Chao, Y. Effect of gamma irradiation on quality of dried potato. Radiat. Phys. Chem. 2003, 66, 293–297. [Google Scholar] [CrossRef]
- Chung, H.J.; Liu, Q. Molecular structure and physicochemical properties of potato and bean starches as affected by gamma-irradiation. Int. J. Biol. Macromol. 2010, 47, 214–222. [Google Scholar] [CrossRef]
- Rezaee, M.; Almassi, M.; Minaei, S.; Paknejad, F. Impact of post-harvest radiation treatment timing on shelf life and quality characteristics of potatoes. J. Food Sci. Technol. 2013, 50, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Li, L.; Chen, L.; Li, X. Study on supramolecular structural changes of ultrasonic treated potato starch granules. Food Hydrocoll. 2012, 29, 116–122. [Google Scholar] [CrossRef]
- Chung, K.M.; Moon, T.W.; Kim, H.; Chun, J.K. Physicochemical properties of sonicated mung bean, potato, and rice starches. Cereal Chem. 2002, 79, 631–633. [Google Scholar] [CrossRef]
- Liu, C.; Grimi, N.; Lebovka, N.; Vorobiev, E. Effects of preliminary treatment by pulsed electric fields and convective air-drying on characteristics of fried potato. Innov. Food Sci. Emerg. Technol. 2018, 47, 454–460. [Google Scholar] [CrossRef]
- Gormley, T.R. Texture studies on mushrooms. Int. J. Food Sci. Technol. 1969, 4, 161–169. [Google Scholar] [CrossRef]
- Sobol, Z.; Jakubowski, T.; Surma, M. Effect of Potato Tuber Exposure to UV-C Radiation and Semi-Product Soaking in Water on Acrylamide Content in French Fries Dry Matter. Sustainability 2020, 12, 3426. [Google Scholar] [CrossRef] [Green Version]
- Sobol, Z.; Jakubowski, T. The effect of storage duration and UV-C stimulation of potato tubers, and soaking of potato strips in water on the density of intermediates of French fries production. Prz. Elektrotech. 2020, 96, 242–245. [Google Scholar] [CrossRef]
- Sobol, Z.; Jakubowski, T.; Wrona, P. The effect of UV-C stimulation of potato tubers and soaking of potato strips in water on density differences of intermediates for French-fry production. In Proceedings of the Contemporary Research Trends in Agricultural Engineering, Proceedings of the BIO Web Conferences, Kraków, Poland, 25–27 September 2017; EDP Sciences: Les Ulis, France, 2018; p. 02031. [Google Scholar] [CrossRef]
- Pelai, Z.; Pedisi, S.; Repaji, M.; Zori, Z.; Levaj, B. Effect of UV-C Irradiation, Storage and Subsequent Cooking on Chemical Constituents of Fresh-Cut Potatoes. Foods 2021, 10, 1698. [Google Scholar] [CrossRef] [PubMed]
- Peleg, M. On fundamental issues in texture evaluation and texturization—A view. Food Hydrocoll. 2006, 20, 405–414. [Google Scholar] [CrossRef]
- Ma, L.; Wang, Q.; Li, L.; Grierson, D.; Yuan, S.; Zheng, S.; Wang, Y.; Wang, B.; Bai, C.; Fu, A.; et al. UV-C irradiation delays the physiological changes of bell pepper fruit during storage. Postharvest Biol. Technol. 2021, 180, 111506. [Google Scholar] [CrossRef]
- Huyskens-Keil, S.; Hassenberg, K.; Herppich, W.B. Impact of postharvest UV-C and ozone treatment on textural properties of white asparagus (Asparagus officinalis L.). J. Appl. Bot. Food Qual. 2012, 84, 229–234. [Google Scholar]
- Poubol, J.; Lichanporn, I.; Puthmee, T.; Kanlayanarat, S. Effect of ultraviolet-C irradiation on quality and natural microflora of asparagus spears. Acta Hortic. 2010, 875, 257–262. [Google Scholar] [CrossRef]
- Manzocco, L.; da Pieve, S.; Maifreni, M. Impact of UV-C light on safety and quality of fresh-cut melon. Innov. Food Sci. Emerg. Technol. 2011, 12, 13–17. [Google Scholar] [CrossRef]
- Manzocco, L.; Da Pieve, S.; Bertolini, A.; Bartolomeoli, I.; Maifreni, M.; Vianello, A.; Nicoli, M.C. Surface decontamination of fresh-cut apple by UV-C light exposure: Effects on structure, colour and sensory properties. Postharvest Biol. Technol. 2011, 61, 165–171. [Google Scholar] [CrossRef]
- Gómez, P.L.; Alzamora, S.A.; Castro, M.A.; Salvatori, D.M. Effect of ultraviolet-C light dose on quality of cut-apple: Microorganism, color and compression behavior. J. Food Eng. 2010, 98, 60–70. [Google Scholar] [CrossRef]
- Gogo, E.O.; Opiyo, A.M.; Hassenberg, K.; Ulrichs, C.; Huyskens-Keil, S. Postharvest UV-C treatment for extending shelf life and improving nutritional quality of African indigenous leafy vegetables. Postharvest Biol. Technol. 2017, 129, 107–117. [Google Scholar] [CrossRef]
- Vunnam, R.; Hussain, A.; Nair, G.; Bandla, R.; Gariepy, Y.; Donnelly, D.J.; Kubow, S.; Raghavan, G.S. Physico-chemical changes in tomato with modified atmosphere storage and UV treatment. J. Food Sci. Technol. 2014, 51, 2106–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cote, S.; Rodoni, L.; Miceli, E.; Concellón, A.; Civello, P.M.; Vicente, A.R. Effect of radiation intensity on the outcome of postharvest UV-C treatments. Postharvest Biol. Technol. 2013, 83, 83–89. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Borland, A.; Singleton, I.; Barnes, J. Impact of atmospheric ozone-enrichment on quality-related attributes of tomato fruit. Postharvest Biol. Technol. 2007, 45, 317–325. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Howard, L. Blueberry fruit response to postharvest application of ultraviolet radiation. Postharvest Biol. Technol. 2008, 47, 280–285. [Google Scholar] [CrossRef]
- Stevens, C.; Liu, J.; Khan, V.A.; Lu, J.Y.; Kabwe, M.K.; Wilson, C.L.; Igwegbe, E.C.; Chalutz, E.; Droby, S. The effects of low-dose ultraviolet light-C treatment on polygalacturonase activity, delay ripening and Rhizopus soft rot development of tomatoes. Crop Prot. 2004, 23, 551–554. [Google Scholar] [CrossRef]
- Tiecher, A.; de Paula, L.A.; Chaves, F.C.; Rombaldi, C.V. UV-C effect on ethylene, polyamines and the regulation of tomato fruit ripening. Postharvest Biol. Technol. 2013, 86, 230–239. [Google Scholar] [CrossRef]
- Obande, M.A.; Tucker, G.A.; Shama, G. Effect of preharvest UV-C treatment of tomatoes (Solanum lycopersicon Mill.) on ripening and pathogen resistance. Postharvest Biol. Technol. 2011, 62, 188–192. [Google Scholar] [CrossRef] [Green Version]
- Bu, J.; Yu, Y.; Aisikaer, G.; Ying, T. Postharvest UV-C irradiation inhibits the production of ethylene and the activity of cell wall-degrading enzymes during softening of tomato (Lycopersicon esculentum L.) fruit. Postharvest Biol. Technol. 2013, 86, 337–345. [Google Scholar] [CrossRef]
- Pan, Y.G.; Zu, H. Effect of UV-C radiation on the quality of fresh-cut pineapples. Procedia Eng. 2012, 37, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Allende, A.; McEvoy, J.L.; Luo, Y.; Artes, F.; Wang, C.Y. Effectiveness of two-sided UV-C treatments in inhibiting natural microflora and extending the shelf-life of minimally processed ‘Red Oak Leaf’ lettuce. Food Microbiol. 2006, 23, 241–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artés, F.; Gómez, P.; Aguayo, E.; Escalona, V.; Artés-Hernández, F. Sustainable sanitation techniques for keeping quality and safety of fresh-cut plant commodities. Postharvest Biol. Technol. 2009, 51, 287–296. [Google Scholar] [CrossRef]
- Edward, T.L.; Kirui, M.S.K.; Omolo, J.O.; Ngumbu, R.G.; Odhiambo, P.M. Effect of Ultraviolet-A (UV-A) and Ultraviolet-C (UV-C) Light on Mechanical Properties of Oyster Mushrooms during Growth. J. Biophys. 2014, 2014, 687028. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, M.; Zhan, Z.; Adhikari, B. Changes in Quality Characteristics of Fresh-cut Cucumbers as Affected by Pressurized Argon Treatment. Food Bioprocess Technol. 2014, 7, 693–701. [Google Scholar] [CrossRef]
- Tabilo-Munizaga, G.; Barbosa-Cánovas, G.V. Rheology for the food industry. J. Food Eng. 2005, 67, 147–156. [Google Scholar] [CrossRef]
- Ohwovoriole, E.N.; Oboli, S.; Mgbeke, A.C. Studies and preliminary design for a cassava tuber peeling machine. Trans. ASAE 1988, 31, 380–0385. [Google Scholar] [CrossRef]
- Araque, L.C.O.; Ortiz, C.M.; Darré, M.; Rodoni, L.M.; Civello, P.M.; Vicente, A.R. Role of UV-C irradiation scheme on cell wall disassembly and surface mechanical properties in strawberry fruit. Postharvest Biol. Technol. 2019, 150, 122–128. [Google Scholar] [CrossRef]
- Jaramillo Sánchez, G.; Contigiani, E.V.; Coronel, M.B.; Alzamora, S.M.; García-Loredo, A.; Nieto, A.B. Study of UV-C treatments on postharvest life of blueberries ‘O’Neal’ and correlation between structure and quality parameters. Heliyon 2021, 7, e07190. [Google Scholar] [CrossRef]
- Mohammed, H.E.S.H.; Suliman, A.E.R.E.; Ahmed, A.E.R.; Ebrahim, M.A. Ultraviolet effect on faba bean seed quality during storage. Asian J. Plant Sci. 2020, 19, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Chudhangkura, A.; Teangpook, C.; Sikkhamondhol, C.; Jariyavattanavijit, C. Effects of ultraviolet C, controlled atmosphere, and ultrasound pretreatment on free ferulic acid in canned sweet corn kernels. J. Food Sci. Technol. 2018, 55, 4167–4173. [Google Scholar] [CrossRef] [PubMed]
- Kasim, M.U.; Kasim, R.; Erkal, S. UV-C treatments on fresh-cut green onions enhanced antioxidant activity, maintained green color and controlled ‘telescoping’. J. Food Agric. Environ. 2008, 6, 63–67. [Google Scholar]
- Ait Barka, E.; Kalantari, S.; Makhlouf, J.; Arul, J. Impact of UV-C irradiation on the cell wall-degrading enzymes during ripening of tomato (Lycopersicon esculentum L.) fruit. J. Agric. Food Chem. 2000, 48, 667–671. [Google Scholar] [CrossRef]
- Zambrano-Zaragoza, M.L.; Quintanar-Guerrero, D.; González-Reza, R.M.; Cornejo-Villegas, M.A.; Leyva-Gómez, G.; Urbán-Morlán, Z. Effects of uv-c and edible nano-coating as a combined strategy to preserve fresh-cut cucumber. Polymer 2021, 13, 3705. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Kim, J.G. Low-dose UV-C irradiation reduces the microbial population and preserves antioxidant levels in peeled garlic (Allium sativum L.) during storage. Postharvest Biol. Technol. 2015, 100, 109–112. [Google Scholar] [CrossRef]
- Castronuovo, D.; Sofo, A.; Lovelli, S.; Candido, V.; Scopa, A. Effects of UV-C radiation on common dandelion and purple coneflower: First results. Int. J. Plant Biol. 2017, 8, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Forges, M.; Bardin, M.; Urban, L.; Aarrouf, J.; Charles, F. Impact of UV-C radiation applied during plant growth on pre-and postharvest disease sensitivity and fruit quality of strawberry. Plant Dis. 2020, 104, 3239–3247. [Google Scholar] [CrossRef]
- Pristijono, P.; Golding, J.B.; Bowyer, M.C. Postharvest UV-C treatment, followed by storage in a continuous low-level ethylene atmosphere, maintains the quality of ‘Kensington pride’ mango fruit stored at 20 °C. Horticulturae 2019, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Pristijono, P.; Bowyer, M.C.; Papoutsis, K.; Scarlett, C.J.; Vuong, Q.V.; Stathopoulos, C.E.; Golding, J.B. Improving the storage quality of Tahitian limes (Citrus latifolia) by pre-storage UV-C irradiation. J. Food Sci. Technol. 2019, 56, 1438–1444. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Wang, C.Y.; Buta, J.G.; Krizek, D.T. Use of UV-C irradiation to prevent decay and maintain postharvest quality of ripe ‘Tommy Atkins’ mangoes. Int. J. Food Sci. Technol. 2008, 36, 767–773. [Google Scholar] [CrossRef]
- Lin, Q.; Xie, Y.; Liu, W.; Zhang, J.; Cheng, S.; Xie, X.; Guan, W.; Wang, Z. UV-C treatment on physiological response of potato (Solanum tuberosum L.) during low temperature storage. J. Food Sci. Technol. 2017, 54, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubowski, T.; Królczyk, J.B. Method for the reduction of natural losses of potato tubers during their long-term storage. Sustainability 2020, 12, 1048. [Google Scholar] [CrossRef] [Green Version]
- Jakubowski, T. Use of UV-C radiation for reducing storage losses of potato tubers. Bangladesh J. Bot. 2018, 47, 533–537. [Google Scholar] [CrossRef] [Green Version]
- Rocha, A.B.O.; Honório, S.L.; Messias, C.L.; Otón, M.; Gómez, P.A. Effect of UV-C radiation and fluorescent light to control postharvest soft rot in potato seed tubers. Sci. Hortic. 2015, 181, 174–181. [Google Scholar] [CrossRef]
- Cools, K.; del Carmen Alamar, M.; Terry, L.A. Controlling sprouting in potato tubers using ultraviolet-C irradiance. Postharvest Biol. Technol. 2014, 98, 106–114. [Google Scholar] [CrossRef]
- Xie, Y.; Lin, Q.; Guan, W.; Cheng, S.; Wang, Z.; Sun, C. Comparison of Sodium Acid Sulfate and UV-C Treatment on Browning and Storage Quality of Fresh-Cut Potatoes. J. Food Qual. 2017, 2017, 5980964. [Google Scholar] [CrossRef] [Green Version]
- Pristijono, P.; Bowyer, M.C.; Scarlett, C.J.; Vuong, Q.V.; Stathopoulos, C.E.; Golding, J.B. Effect of UV-C irradiation on sprouting of potatoes in storage. In Proceedings of the VIII International Postharvest Symposium: Enhancing Supply Chain and Consumer Benefits-Ethical and Technological Issues 1194, Cartagena, Spain, 21–24 June 2016; pp. 475–478. [Google Scholar] [CrossRef]
- Jakubowski, T. Impact of UV-C Irradiation of Potato Seed Tubers on the Defects in Potato Plant Crops. Agric. Eng. 2019, 23, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.; Khan, V.A.; Lu, J.Y.; Wilson, C.L.; Chalutz, E.; Droby, S.; Kabwe, M.K.; Haung, Z.; Adeyeye, O.; Pusey, L.P.; et al. Induced resistance of sweetpotato to Fusarium root rot by UV-C hormesis. Crop Prot. 1999, 18, 463–470. [Google Scholar] [CrossRef]
- Stevens, C.; Khan, V.A.; Lu, J.Y.; Wilson, C.L.; Pusey, P.L.; Igwegbe, E.C.; Kabwe, K.; Mafolo, Y.; Liu, J.; Chalutz, E.; et al. Integration of ultraviolet (UV-C) light with yeast treatment for control of postharvest storage rots of fruits and vegetables. Biol. Control 1997, 10, 98–103. [Google Scholar] [CrossRef]
- Mercier, J. Role of phytoalexins and other antimicrobial compounds from fruits and vegetables in postharvest disease resistance. In Proceedings-Phytochemical Society of Europe; Oxford University Press Inc.: Oxford, UK, 1996; Volume 41, pp. 221–242. [Google Scholar]
- Gonzalez-Aguilar, G.A.; Villa-Rodriguez, J.A.; Ayala-Zavala, J.F.; Yahia, E.M. Improvement of the antioxidant status of tropical fruits as a secondary response to some postharvest treatments. Trends Food Sci. Technol. 2010, 219100, 475–482. [Google Scholar] [CrossRef]
- Shama, G. Process challenges in applying low doses of ultraviolet light to fresh produce for eliciting beneficial hormetic responses. Postharvest Biol. Technol. 2007, 44, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Vicente, A.R.; Martínez, G.A.; Chaves, A.R.; Civello, P.M. Combined use of UV-C irradiation and heat treatment to improve postharvest life of strawberry fruit. J. Sci. Food Agric. 2004, 84, 1831–1838. [Google Scholar] [CrossRef]
- Ortiz Araque, L.C.; Rodoni, L.M.; Darré, M.; Ortiz, C.M.; Civello, P.M.; Vicente, A.R. Cyclic low dose UV-C treatments retain strawberry fruit quality more effectively than conventional pre-storage single high fluence applications. LWT-Food Sci. Technol. 2018, 92, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Hassan, H.; Abd El-Rahman, A.; Liela, A. Sprouting suppression and quality attributes of potato tubers as affected by post-harvest UV-C treatment under cold storage. Int. J. Adv. Res 2016, 4, 241–253. [Google Scholar] [CrossRef] [Green Version]
- Shama, G.; Alderson, P. UV hormesis in fruits: A concept ripe for commercialization. Trends Food Sci. Technol. 2005, 16, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Terry, L.A.; Joyce, D.C. Elicitors of induced disease resistance in postharvest horticultural crops: A brief review. Postharvest Biol. Technol. 2004, 32, 1–13. [Google Scholar] [CrossRef]
- Xu, Y.; Charles, M.T.; Luo, Z.; Mimee, B.; Tong, Z.; Véronneau, P.Y.; Rolland, D.; Roussel, D. Preharvest ultraviolet C treatment affected senescence of stored strawberry fruit with a potential role of microRNAs in the activation of the antioxidant system. J. Agric. Food Chem. 2018, 66, 12188–12197. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, W. UV treatment improved the quality of postharvest fruits and vegetables by inducing resistance. Trends Food Sci. Technol. 2019, 92, 71–80. [Google Scholar] [CrossRef]
- Xu, Y.; Charles, M.T.; Luo, Z.; Mimee, B.; Tong, Z.; Roussel, D.; Rolland, D.; Véronneau, P.Y. Preharvest UV-C treatment affected postharvest senescence and phytochemicals alternation of strawberry fruit with the possible involvement of abscisic acid regulation. Food Chem. 2019, 299, 125138. [Google Scholar] [CrossRef]
- Vàsquez, H.; Ouhibi, C.; Lizzi, Y.; Azzouz, N.; Forges, M.; Bardin, M.; Nicot, P.; Urban, L.; Aarrouf, J. Pre-harvest hormetic doses of UV-C radiation can decrease susceptibility of lettuce leaves (Lactuca sativa L.) to Botrytis cinerea L. Sci. Hortic. 2017, 222, 32–39. [Google Scholar] [CrossRef]
- Ouhibi, C.; Attia, H.; Nicot, P.; Lecompte, F.; Vidal, V.; Lachaâl, M.; Urban, L.; Aarrouf, J. Effects of nitrogen supply and of UV-C irradiation on the susceptibility of Lactuca sativa L. to Botrytis cinerea and Sclerotinia minor. Plant Soil 2015, 393, 35–46. [Google Scholar] [CrossRef]
- Yang, Z.; Cao, S.; Su, X.; Jiang, Y. Respiratory activity and mitochondrial membrane associated with fruit senescence in postharvest peaches in response to UV-C treatment. Food Chem. 2014, 161, 16–21. [Google Scholar] [CrossRef]
- Pombo, M.A.; Rosli, H.G.; Martínez, G.A.; Civello, P.M. UV-C treatment affects the expression and activity of defense genes in strawberry fruit (Fragaria × ananassa, Duch.). Postharvest Biol. Technol. 2011, 59, 94–102. [Google Scholar] [CrossRef]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants: Growth, antioxidant activity and phenolic compounds. Plant Physiol. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef]
- Charles, M.T.; Tano, K.; Asselin, A.; Arul, J. Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit. V. Constitutive defence enzymes and inducible pathogenesis-related proteins. Postharvest Biol. Technol. 2009, 51, 414–424. [Google Scholar] [CrossRef]
- Urban, L.; Charles, F.; de Miranda, M.R.A.; Aarrouf, J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol. Biochem. 2016, 105, 1–11. [Google Scholar] [CrossRef]
- Urban, L.; Chabane Sari, D.; Orsal, B.; Lopes, M.; Miranda, R.; Aarrouf, J. UV-C light and pulsed light as alternatives to chemical and biological elicitors for stimulating plant natural defenses against fungal diseases. Sci. Hortic. 2018, 235, 452–459. [Google Scholar] [CrossRef]
- Liu, C.; Cai, L.; Han, X.; Ying Tiejin, T. Temporary effect of postharvest UV-C irradiation on gene expression profile in tomato fruit. Gene 2011, 486, 56–64. [Google Scholar] [CrossRef]
- Duarte-Sierra, A.; Nadeau, F.; Angers, P.; Michaud, D.; Arul, J. UV-C hormesis in broccoli florets: Preservation, phyto-compounds and gene expression. Postharvest Biol. Technol. 2019, 157, 110965. [Google Scholar] [CrossRef]
- Li, D.; Luo, Z.; Mou, W.; Wang, Y.; Ying, T.; Mao, L. ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Lamikanra, O.; Kueneman, D.; Ukuku, D.; Bett-Garber, K.L. Effect of Processing Under Ultraviolet Light on the Shelf Life of Fresh-Cut Cantaloupe Melon. J. Food Sci. 2005, 70, C534–C539. [Google Scholar] [CrossRef]
- Prasanna, V.; Prabha, T.N.; Tharanathan, R.N. Fruit ripening phenomena—An overview. Crit. Rev. Food Sci. Nutr. 2007, 47, 1–19. [Google Scholar] [CrossRef]
- Artés-Hernández, F.; Escalona, V.H.; Robles, P.A.; Martínez-Hernández, G.B.; Artés, F. Effect of UV-C radiation on quality of minimally processed spinach leaves. J. Sci. Food Agric. 2009, 89, 414–421. [Google Scholar] [CrossRef]
- Vicente, A.R.; Pineda, C.; Lemoine, L.; Civello, P.M.; Martinez, G.A.; Chaves, A.R. UV-C treatments reduce decay, retain quality and alleviate chilling injury in pepper. Postharvest Biol. Technol. 2005, 35, 69–78. [Google Scholar] [CrossRef]
- Phonyiam, O.; Ohara, H.; Kondo, S.; Naradisorn, M.; Setha, S. Postharvest UV-C Irradiation Influenced Cellular Structure, Jasmonic Acid Accumulation, and Resistance against Green Mold Decay in Satsuma Mandarin Fruit (Citrus unshiu). Front. Sustain. Food Syst. 2021, 5, 336. [Google Scholar] [CrossRef]
- Sripong, K.; Jitareerat, P.; Uthairatanakij, A. UV irradiation induces resistance against fruit rot disease and improves the quality of harvested mangosteen. Postharvest Biol. Technol. 2019, 149, 187–194. [Google Scholar] [CrossRef]
- Imaizumi, T.; Yamauchi, M.; Sekiya, M.; Shimonishi, Y.; Tanaka, F. Responses of phytonutrients and tissue condition in persimmon and cucumber to postharvest UV-C irradiation. Postharvest Biol. Technol. 2018, 145, 33–40. [Google Scholar] [CrossRef]
- Wang, D.; Chen, L.; Ma, Y.; Zhang, M.; Zhao, Y.; Zhao, X. Effect of UV-C treatment on the quality of fresh-cut lotus (Nelumbo nucifera Gaertn.) root. Food Chem. 2019, 278, 659–664. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of postharvest UV-C irradiation on phenolic acids, flavonoids, and key phenylpropanoid pathway genes in tomato fruit. Sci. Hortic. 2018, 241, 107–114. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Han, C.; Ji, N.; Jin, P.; Zheng, Y. UV-C treatment maintains quality and enhances antioxidant capacity of fresh-cut strawberries. Postharvest Biol. Technol. 2019, 156, 110945. [Google Scholar] [CrossRef]
- Prajapati, U.; Asrey, R.; Varghese, E.; Singh, A.K.; Pal Singh, M. Effects of postharvest ultraviolet-C treatment on shelf-life and quality of bitter gourd fruit during storage. Food Packag. Shelf Life 2021, 28, 100665. [Google Scholar] [CrossRef]
- Charles, M.T.; Benhamou, N.; Arul, J. Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit: III. Ultrastructural modifications and their impact on fungal colonization. Postharvest Biol. Technol. 2008, 47, 27–40. [Google Scholar] [CrossRef]
- Quintero-Cerón, J.P.; Bohorquez-Pérez, Y.; Valenzuela-Rea, C.; Solanilla-Duque, J.F. Advancements in the application of short-wave ultraviolet light (UVC) in whole and fresh-cut fruit and vegetables: A review. Rev. Tumbaga 2013, 8, 29–60. [Google Scholar]
- Rivera-Pastrana, D.M.; Gardea Béjar, A.A.; Martínez-Téllez, M.A.; Rivera-Domínguez, M.; González-Aguilar, G.A. Postharvest biochemical effects of UV-C irradiation on fruit and vegetables. Artículo Revisión Rev. Fitotec. Mex 2007, 30, 361–372. [Google Scholar]
- Nigro, F.; Ippolito, A.; Lima, G. Use of UV-C light to reduce Botrytis storage rot of table grapes. Postharvest Biol. Technol. 1998, 13, 171–181. [Google Scholar] [CrossRef]
- Wang, C.Y.; Chen, C.T.; Wang, S.Y. Changes of flavonoid content and antioxidant capacity in blueberries after illumination with UV-C. Food Chem. 2009, 117, 426–431. [Google Scholar] [CrossRef]
- Sun, T.; Ouyang, H.; Sun, P.; Zhang, W.; Wang, Y.; Cheng, S.; Chen, G. Postharvest UV-C irradiation inhibits blackhead disease by inducing disease resistance and reducing mycotoxin production in ‘Korla’fragrant pear (Pyrus sinkiangensis). Int. J. Food Microbiol. 2022, 362, 109485. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, S.; Qiao, L.; Chen, J.; Liu, D.; Ye, X. Effect of UV-C treatments on phenolic compounds and antioxidant capacity of minimally processed Satsuma mandarin during refrigerated storage. Postharvest Biol. Technol. 2013, 76, 50–57. [Google Scholar] [CrossRef]
- Gutiérrez, D.R.; Rodríguez, S.D.C. Combined Effect of UV-C and Ozone on Bioactive Compounds and Microbiological Quality of Fresh-Cut Rocket Leaves. Am. J. Food Sci. Technol. 2019, 7, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Liu, W.; Yuan, L.; Guan, W.Q.; Brennan, C.S.; Zhang, Y.Y.; Zhang, J.; Wang, Z.D. The influence of postharvest UV-C treatment on anthocyanin biosynthesis in fresh-cut red cabbage. Sci. Rep. 2017, 7, 5232. [Google Scholar] [CrossRef]
- González-Aguilar, G.A.; Zavaleta-Gatica, R.; Tiznado-Hernández, M.E. Improving postharvest quality of mango ‘Haden’ by UV-C treatment. Postharvest Biol. Technol. 2007, 45, 108–116. [Google Scholar] [CrossRef]
- Watada, A.E.; Ko, N.P.; Minott, D.A. Factors affecting quality of fresh-cut horticultural products. Postharvest Biol. Technol. 1996, 9, 115–125. [Google Scholar] [CrossRef]
- Brecht, J.K. Physiology of lightly processed fruits and vegetables. HortScience 1995, 30, 18–22. [Google Scholar] [CrossRef]
- Abe, F.; Saito, K.; Miura, K.; Toriyama, K. A single nucleotide polymorphism in the alternative oxidase gene among rice varieties differing in low temperature tolerance. FEBS Lett. 2002, 527, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, C.; Canada, J.; Alvarenga, B. Prospects of UV radiation for application in postharvest technology. Emir. J. Food Agric. 2012, 24, 586–597. [Google Scholar] [CrossRef]
- Reed, D.W.; Tukey, H.B.J. Light intensity and temperature effects on epicuticular wax morphology and internal cuticle ultrastructure of carnation and Brussels sprouts leaf cuticles. J. Am. Soc. Hortic. Sci. 1982, 107, 417–420. [Google Scholar]
- Delorme, M.M.; Guimarães, J.T.; Coutinho, N.M.; Balthazar, C.F.; Rocha, R.S.; Silva, R.; Margalho, L.P.; Pimentel, T.C.; Silva, M.C.; Freitas, M.Q.; et al. Ultraviolet radiation: An interesting technology to preserve quality and safety of milk and dairy foods. Trends Food Sci. Technol. 2020, 102, 146–154. [Google Scholar] [CrossRef]
- Abdipour, M.; Sadat Malekhossini, P.; Hosseinifarahi, M.; Radi, M. Integration of UV irradiation and chitosan coating: A powerful treatment for maintaining the postharvest quality of sweet cherry fruit. Sci. Hortic. 2020, 264, 109197. [Google Scholar] [CrossRef]
- Park, S.H.; Kang, J.W.; Kang, D.H. Inactivation of foodborne pathogens on fresh produce by combined treatment with UV-C radiation and chlorine dioxide gas, and mechanisms of synergistic inactivation. Food Control 2018, 92, 331–340. [Google Scholar] [CrossRef]
- Choi, D.S.; Park, S.H.; Choi, S.R.; Kim, J.S.; Chun, H.H. The combined effects of ultraviolet-C irradiation and modified atmosphere packaging for inactivating Salmonella enterica serovar Typhimurium and extending the shelf life of cherry tomatoes during cold storage. Food Packag. Shelf Life 2015, 3, 19–30. [Google Scholar] [CrossRef]
- Jemni, M.; Gómez, P.A.; Souza, M.; Chaira, N.; Ferchichi, A.; Otón, M.; Artés, F. Combined effect of UV-C, ozone and electrolyzed water for keeping overall quality of date palm. LWT-Food Sci. Technol. 2014, 59, 649–655. [Google Scholar] [CrossRef]



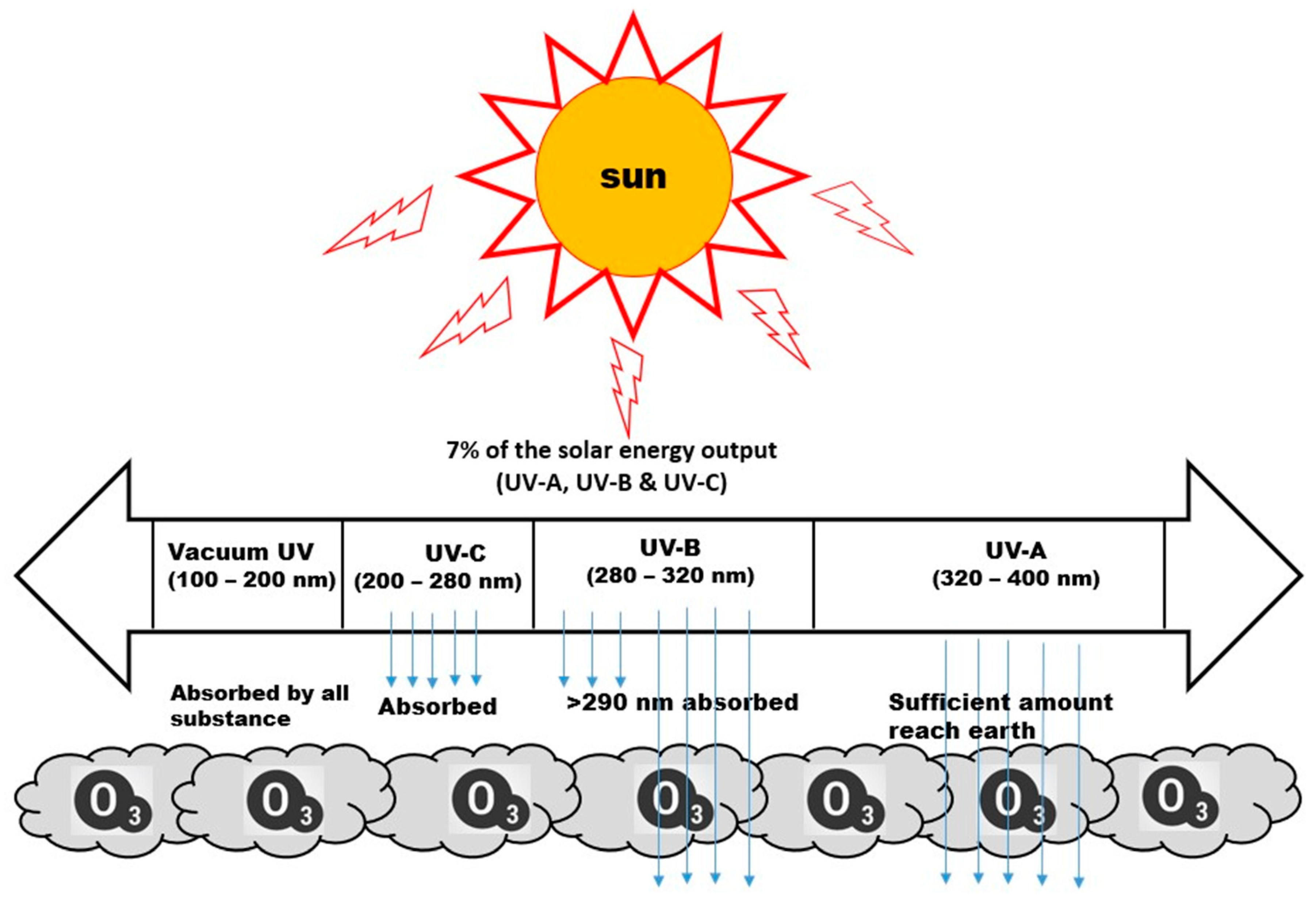{kind=link}
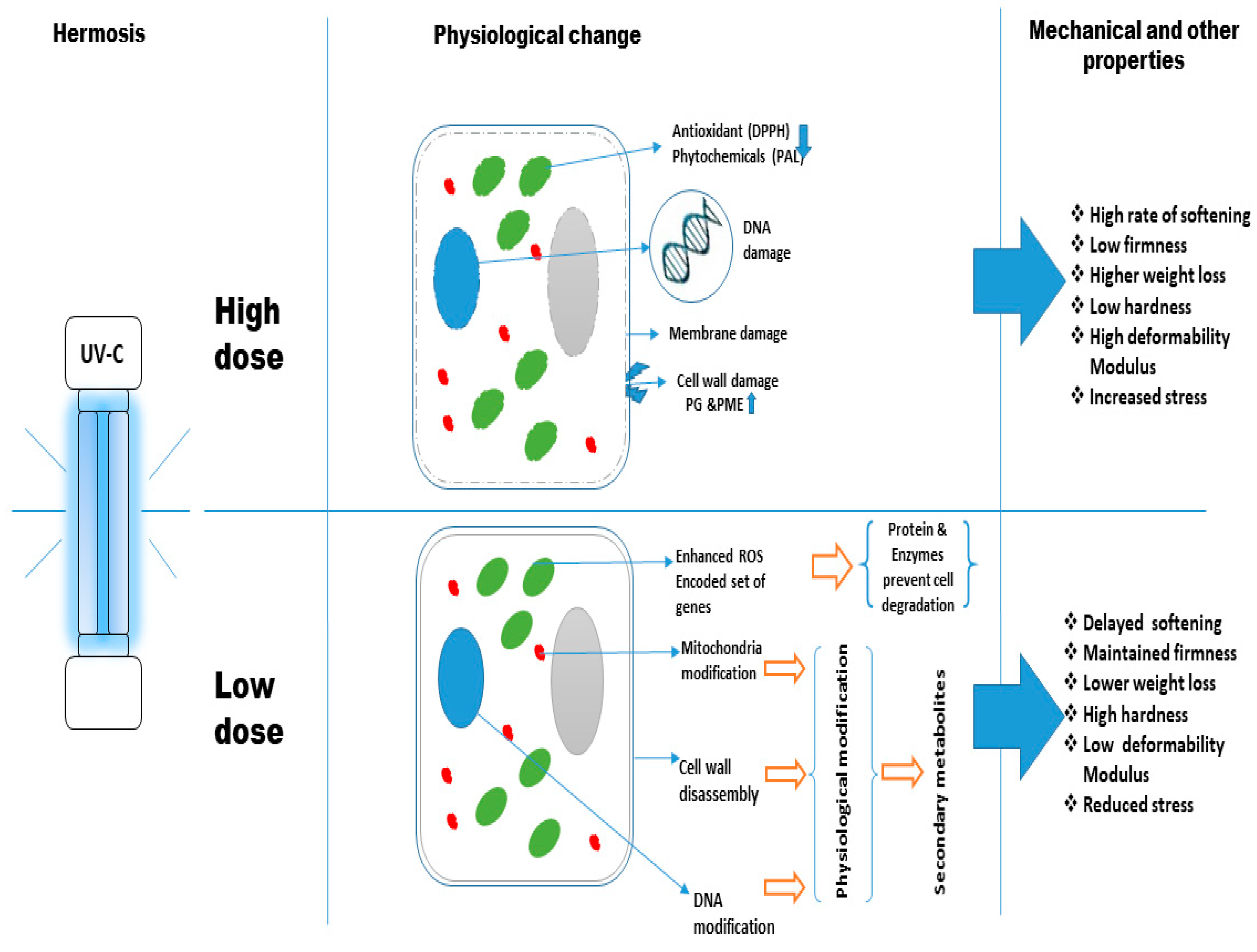{kind=link}
| Commodity | Operational Condition | Key Finding | Reference |
|---|---|---|---|
| Sweet corn kernels | UV-C dose at 0. 1.94, and 4.01 kJ∙m−2, controlled atmosphere of with %oxygen: %carbondioxide: %nitrogen ratios of 21:0.03:78, 3:10:87, and 3:15:82 at 6 °C for 20 h. | Hardness remains unchanged | [83] |
| Fresh-cut green onion | UV-C exposure time at 3, 5, 10, and 15 min and storage days of 5, 10, 15 days and storage temperature of 5 °C. | Higher UV-C exposure results in higher weight loss (%). | [84] |
| Tomato (Lycopersicon esculentum L.) | UV-C dose 3.7 kJ∙m−2 from 0 to 25 days of storage duration time at 16 °C and relative humidity of 95%. | Firmness (Newton) decreased with storage duration. Higher resistance penetration compared to the control sample. | [85] |
| Cucumber (Cucumis sativus L.) | UV-C dose of 4.5 kJ∙m−2 stored for 15 days at 4 °C, a combination of UV-C with Nano-coating Nanocapsules. | The UV-C control sample brought better firmness, as the loss was delayed to day 9 of storage. | [86] |
| Peeled garlic (Allium sativum L.) | UV-C dose of 2 kJ∙m−2 stored for 15 days at room temperature. | High firmness value with the UV-C treated sample. | [87] |
| Cherry tomato | UV-C dose of 3.7 kJ∙m−2 | UV-C treated and control sample both mass loss and firmness were unaffected. | [65] |
| Tomato (Lycopersicon esculentum L.) | UV-C dose of 4.2 kJ∙m−2 for 8 min | The firmness decreased gradually during storage in both the control and UV-C treated tomatoes. | [72] |
| Common dandelion and purple coneflower | UV-C dose of 3.8 J∙m−2, 1 m of distance from light source, 10 to 120 exposure time, and 21 days of storage period. | Fresh and dry weight loss recorded for both dandelion and purple coneflower is higher than the control sample. | [88] |
| Strawberry | UV-C dose of 1.70 kJ∙m−2 for 4.8 min and Storage duration of 0, 2, and 4 days at 21 °C. | No difference in firmness between fruit from control and UV-C-treated samples. | [89] |
| Mango (Kensington Pride) | UV-C dose at 4.0, 8.3, and 11.7 kJ∙m−2), room temperature (20 ± 1 °C), relative humidity at 80%, and ethylene storage duration from 3 to 12 days. | At a higher UV-C dose, the firmness is significantly higher than untreated fruits after 6 days of storage. No significant difference in weight loss with the control sample. | [90] |
| Tahitian lime (Citrus latifolia) | The doses were 3.4, 7.2, and 10.5 kJ∙m−2. Fruits were located 20 cm from the UV-C light source. | Higher dose (10.5 kJ∙m−2) reduced weight loss. | [91] |
| Mango (Tommy Atkins) | UV-C dose of 8220 mW∙m−2 and exposure time of 10 and 20 min. and storage temperature 5 and 20 °C | Lower weight loss and high firmness for Samples exposed to 10 min and 5 °C. | [92] |
| Commodity | Operating Condition | Key Finding | Reference |
|---|---|---|---|
| Tomato (Solanum lycopersicum) | UV-C dose of 4 kJ∙m−2, exposure time of 6 min. | Increase in phenolic compounds content. | [133] |
| Blueberries | UV-C dose of 0.43, 2.15, 4.30 and 6.45 kJ∙m−2. | Higher antioxidant capacity in fruit treated with 2.15, 4.30, and 6.45 kJ∙m−2 compared to the control fruit. Increased phenolic components in a lesser amount at 0.43 kJ∙m−2. | [140] |
| Fragrant pear (Korla) | UV-C irradiation of 0.12, 0.24, 0.36, 0.48, 0.72 and 1.08 kJ∙m−2. | low-dose UV-C irradiation (0.36 kJ∙m−2) enhanced the phenolic compound. | [141] |
| Mandarin (Satsuma) | UV-C dose of 0.75, 1.5 and 3.0 kJ∙m−2. | Phenolic acids and antioxidant capacity were not significantly affected by UV-C treatments, while 1.5 and 3.0 kJ∙m−2 significantly increased flavonoids and total phenolics. 0.75 kJ∙m−2 was infective to induce any change. | [142] |
| Garlic (Danyang) | UV-C of 25 kJ∙m−2, exposure time of 380 s. | The UV-C treatment reduces microorganisms present and no significant differences in quality attributes, phenolic compounds, and antioxidants. | [143] |
| Fresh-cut Rocket Leaves | UV-C dose at 1, 3 and 5 kJ∙m−2. | The optimum dose of UV-C for enhancing total anthocyanin content was 3.0 kJ∙m−2. | [144] |
| Mango (Haden) | UV-C dose of 2.46 and 4.93 kJ∙m−2. | the highest accumulation of phytochemicals in mangoes exposed to 4.93 kJ∙m−2. | [145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemessa, A.; Popardowski, E.; Hebda, T.; Jakubowski, T. The Effect of UV-C Irradiation on the Mechanical and Physiological Properties of Potato Tuber and Different Products. Appl. Sci. 2022, 12, 5907. https://doi.org/10.3390/app12125907
Lemessa A, Popardowski E, Hebda T, Jakubowski T. The Effect of UV-C Irradiation on the Mechanical and Physiological Properties of Potato Tuber and Different Products. Applied Sciences. 2022; 12(12):5907. https://doi.org/10.3390/app12125907
Chicago/Turabian StyleLemessa, Addis, Ernest Popardowski, Tomasz Hebda, and Tomasz Jakubowski. 2022. "The Effect of UV-C Irradiation on the Mechanical and Physiological Properties of Potato Tuber and Different Products" Applied Sciences 12, no. 12: 5907. https://doi.org/10.3390/app12125907