
Use Electroencephalogram Entropy as an Indicator to Detect Stress-Induced Sleep Alteration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Surgery
2.3. Apparatus
2.4. Experimental Procedure
2.5. Data Analysis
2.5.1. Analysis of the Vigilance States
2.5.2. EEG Parameter Investigation
Spectrum
Instantaneous Amplitude, Frequency, and Phase
Shannon Entropy
2.6. Statistics
3. Results
3.1. Alterations of Sleep Duration and Spectrogram after Footshock
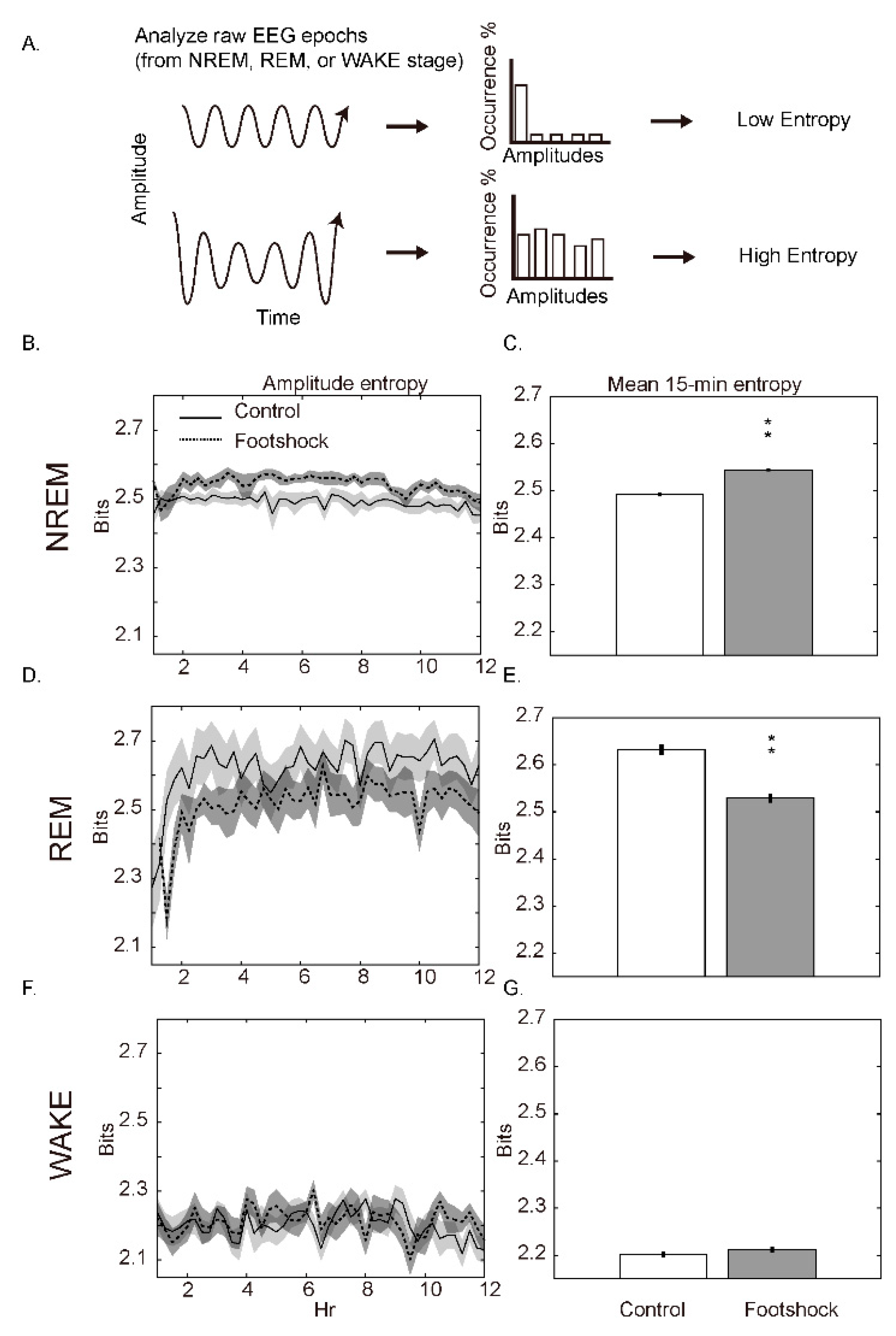
3.2. Amplitude Entropy
3.3. Frequency Entropy
3.4. Phase Entropy
4. Discussion
4.1. Footshock Reduces REM Sleep Duration
4.2. The EEG Spectrum after Footshock
4.3. Amplitude Entropy
4.4. Frequency Entropy
4.5. Phase Entropy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASD | Acute stress disorder |
| EEG | Electroencephalography |
| NREM | Non-rapid-eye movement |
| PTSD | Post-traumatic stress disorder |
| REM | Rapid-eye movement |
References
- Buzsaki, G. Rhythms of the Brain; Oxford University Press: Oxford, UK; New York, NY, USA, 2006. [Google Scholar]
- Jouvet, M. Neurophysiology of the states of sleep. Physiol. Rev. 1967, 47, 117–177. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyden, W.M.; Poe, G.R.; Liberzon, I. Trauma exposure and sleep: Using a rodent model to understand sleep function in PTSD. Exp. Brain Res. 2014, 232, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Vanderheyden, W.M.; George, S.A.; Urpa, L.; Kehoe, M.; Liberzon, I.; Poe, G.R. Sleep alterations following exposure to stress predict fear-associated memory impairments in a rodent model of PTSD. Exp. Brain Res. 2015, 233, 2335–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Dimsdale, J.E. The effect of psychosocial stress on sleep: A review of polysomnographic evidence. Behav. Sleep Med. 2007, 5, 256–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibodeau, R.; Jorgensen, R.S.; Kim, S. Depression, anxiety, and resting frontal EEG asymmetry: A meta-analytic review. J. Abnorm. Psychol. 2006, 115, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Vanderwolf, C.H. Hippocampal electrical activity and voluntary movement in the rat. Electroencephalogr. Clin. Neurophysiol. 1969, 26, 407–418. [Google Scholar] [CrossRef]
- Hsiao, Y.T.; Yi, P.L.; Cheng, C.H.; Chang, F.C. Disruption of footshock-induced theta rhythms by stimulating median raphe nucleus reduces anxiety in rats. Behav. Brain Res. 2013, 247, 193–200. [Google Scholar] [CrossRef]
- Seidenbecher, T.; Laxmi, T.R.; Stork, O.; Pape, H.C. Amygdalar and hippocampal theta rhythm synchronization during fear memory retrieval. Science 2003, 301, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Lever, C.; Kaplan, R.; Burgess, N. The Function of Oscillations in the Hippocampal Formation. In Space, Time and Memory in the Hippocampal Formation; Derdikman, D., Knierim, J.J., Eds.; Springer Vienna: Vienna, Austria, 2014; pp. 303–350. [Google Scholar]
- Wells, C.E.; Amos, D.P.; Jeewajee, A.; Douchamps, V.; Rodgers, J.; O’Keefe, J.; Burgess, N.; Lever, C. Novelty and anxiolytic drugs dissociate two components of hippocampal theta in behaving rats. J. Neurosci. 2013, 33, 8650–8667. [Google Scholar] [CrossRef] [Green Version]
- Winson, J. Loss of hippocampal theta rhythm results in spatial memory deficit in the rat. Science 1978, 201, 160–163. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Perez-Garci, E.; Del Rio-Portilla, Y.; Ugalde, E.; Guevara, M.A. EEG bands during wakefulness, slow-wave, and paradoxical sleep as a result of principal component analysis in the rat. Sleep 2001, 24, 374–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Psychiatric Association. DSM-V: Diagnostic and Statistical Manual of Mental Disorders: DSM-5; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Bastien, C.H.; Vallieres, A.; Morin, C.M. Precipitating factors of insomnia. Behav. Sleep Med. 2004, 2, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Benca, R.M.; Obermeyer, W.H.; Larson, C.L.; Yun, B.; Dolski, I.; Kleist, K.D.; Weber, S.M.; Davidson, R.J. EEG alpha power and alpha power asymmetry in sleep and wakefulness. Psychophysiology 1999, 36, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Horne, J.A. Prefrontal cortex: Links between low frequency delta EEG in sleep and neuropsychological performance in healthy, older people. Psychophysiology 2003, 40, 349–357. [Google Scholar] [CrossRef]
- Franken, P.; Dijk, D.J.; Tobler, I.; Borbely, A.A. Sleep deprivation in rats: Effects on EEG power spectra, vigilance states, and cortical temperature. Am. J. Physiol. 1991, 261, R198–R208. [Google Scholar] [CrossRef]
- Borbely, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar]
- Tort, A.B.; Kramer, M.A.; Thorn, C.; Gibson, D.J.; Kubota, Y.; Graybiel, A.M.; Kopell, N.J. Dynamic cross-frequency couplings of local field potential oscillations in rat striatum and hippocampus during performance of a T-maze task. Proc. Natl. Acad. Sci. USA 2008, 105, 20517–20522. [Google Scholar] [CrossRef] [Green Version]
- Tort, A.B.; Komorowski, R.W.; Manns, J.R.; Kopell, N.J.; Eichenbaum, H. Theta-gamma coupling increases during the learning of item-context associations. Proc. Natl. Acad. Sci. USA 2009, 106, 20942–20947. [Google Scholar] [CrossRef] [Green Version]
- Hinman, J.R.; Penley, S.C.; Long, L.L.; Escabi, M.A.; Chrobak, J.J. Septotemporal variation in dynamics of theta: Speed and habituation. J. Neurophysiol. 2011, 105, 2675–2686. [Google Scholar] [CrossRef]
- Ahmed, O.J.; Mehta, M.R. Running speed alters the frequency of hippocampal gamma oscillations. J. Neurosci. 2012, 32, 7373–7383. [Google Scholar] [CrossRef]
- Zheng, C.; Bieri, K.W.; Trettel, S.G.; Colgin, L.L. The relationship between gamma frequency and running speed differs for slow and fast gamma rhythms in freely behaving rats. Hippocampus 2015, 25, 924–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Bokil, H.; Andrews, P.; Kulkarni, J.E.; Mehta, S.; Mitra, P.P. Chronux: A platform for analyzing neural signals. J. Neurosci. Meth. 2010, 192, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Sigurdsson, T.; Topiwala, M.A.; Gordon, J.A. Cross-correlation of instantaneous amplitudes of field potential oscillations: A straightforward method to estimate the directionality and lag between brain areas. J. Neurosci. Methods 2010, 191, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Freedman, D.; Diaconis, P. On the histogram as a density estimator: L2 theory. Z. Wahrscheinlichkeitstheorie Verwandte Geb. 1981, 57, 453–476. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.K.; Brennan, F.X.; Pawlyk, A.C.; Ross, R.J.; Morrison, A.R. REM sleep: A sensitive index of fear conditioning in rats. Eur. J. Neurosci. 2005, 21, 1077–1080. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, R.; Sanford, L.D.; Yang, L.; Zhou, J.; Zhang, J.; Wing, Y.K.; Shi, J.; Lu, L.; Tang, X. Sleep in posttraumatic stress disorder: A systematic review and meta-analysis of polysomnographic findings. Sleep Med. Rev. 2019, 48, 101210. [Google Scholar] [CrossRef]
- Sainsbury, R.S.; Heynen, A.; Montoya, C.P. Behavioral correlates of hippocampal type 2 theta in the rat. Physiol. Behav. 1987, 39, 513–519. [Google Scholar] [CrossRef]
- O’Keefe, J. Hippocampus, theta, and spatial memory. Curr. Opin. Neurobiol. 1993, 3, 917–924. [Google Scholar] [CrossRef]
- Bieri, K.W.; Bobbitt, K.N.; Colgin, L.L. Slow and fast gamma rhythms coordinate different spatial coding modes in hippocampal place cells. Neuron 2014, 82, 670–681. [Google Scholar] [CrossRef] [Green Version]
- Cowdin, N.; Kobayashi, I.; Mellman, T.A. Theta frequency activity during rapid eye movement (REM) sleep is greater in people with resilience versus PTSD. Exp. Brain Res. 2014, 232, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.L.; Denninger, T.; Fyhn, M.; Hafting, T.; Bonnevie, T.; Jensen, O.; Moser, M.B.; Moser, E.I. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009, 462, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Pearl, P.L.; LaFleur, B.J.; Reigle, S.C.; Rich, A.S.; Freeman, A.A.; McCutchen, C.; Sato, S. Sawtooth wave density analysis during REM sleep in normal volunteers. Sleep Med. 2002, 3, 255–258. [Google Scholar] [CrossRef]
- Poe, G.R. Sleep Is for Forgetting. J. Neurosci. 2017, 37, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, Y.; Hsiao, Y.-T.; Chang, F.-C. Use Electroencephalogram Entropy as an Indicator to Detect Stress-Induced Sleep Alteration. Appl. Sci. 2022, 12, 4812. https://doi.org/10.3390/app12104812
Lo Y, Hsiao Y-T, Chang F-C. Use Electroencephalogram Entropy as an Indicator to Detect Stress-Induced Sleep Alteration. Applied Sciences. 2022; 12(10):4812. https://doi.org/10.3390/app12104812
Chicago/Turabian StyleLo, Yun, Yi-Tse Hsiao, and Fang-Chia Chang. 2022. "Use Electroencephalogram Entropy as an Indicator to Detect Stress-Induced Sleep Alteration" Applied Sciences 12, no. 10: 4812. https://doi.org/10.3390/app12104812