
Population Dynamics and Yeast Diversity in Early Winemaking Stages without Sulfites Revealed by Three Complementary Approaches

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Isolation Procedure
2.2. MALDI-TOF MS
2.2.1. Validation of Yeast Identification by MALDI-TOF MS Biotyper
2.2.2. MALDI-TOF MS Oenological Laboratory Specific Database (OLS-DB)
2.3. Red Wine Vinification Process and Sampling
2.4. Yeast and Fungi Community Analysis
2.4.1. DNA Extraction
2.4.2. Population Dynamics of Targeted Microorganisms by Quantitative PCR
2.4.3. Yeast Biodiversity Analysis
- Meta-barcoding and high-throughput sequencing analysis (HTS)
- Yeast diversity analysis by MALDI-TOF MS
2.5. Statistical Analysis
3. Results
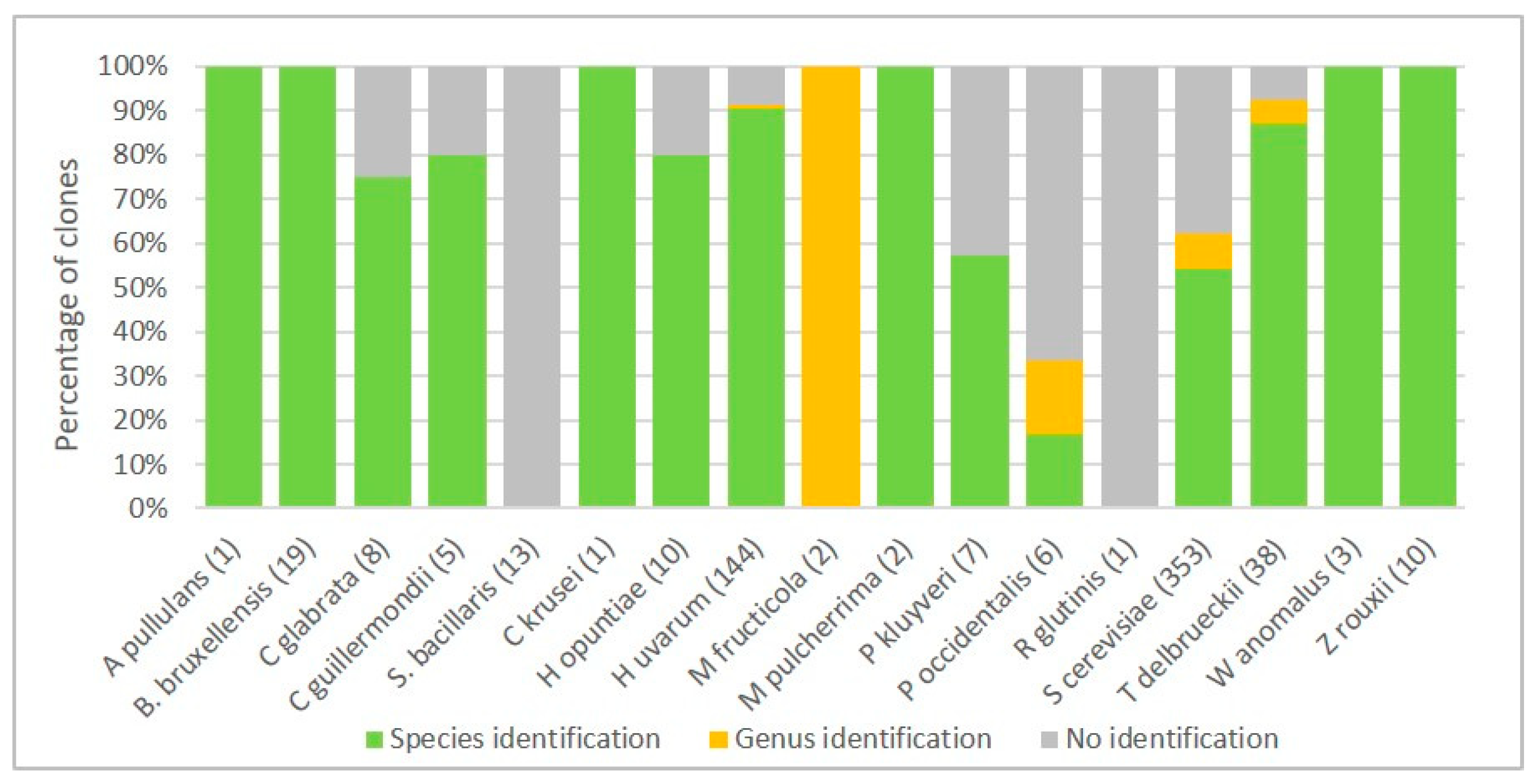
3.1. Optimization of Wine Yeast Identification by MALDI-TOF MS
3.1.1. Comparative Analysis of Species Identification by MALDI-TOF MS Biotyper Database and 26S rDNA Sequencing
3.1.2. Oenological Laboratory Specific Database of MALDI-TOF MS
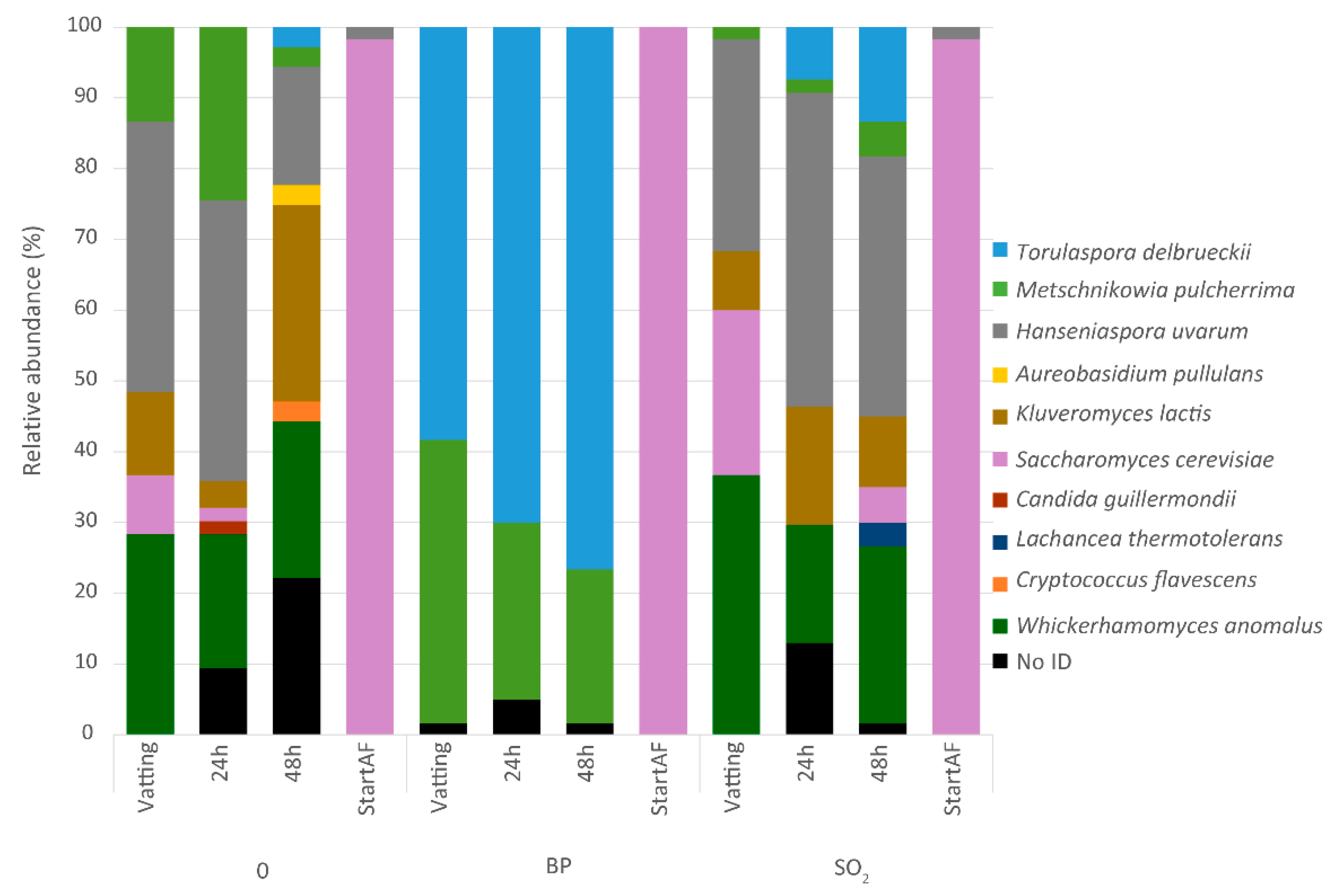
3.2. Yeast and Fungi Community during the Early Stages of Winemaking without Sulfites
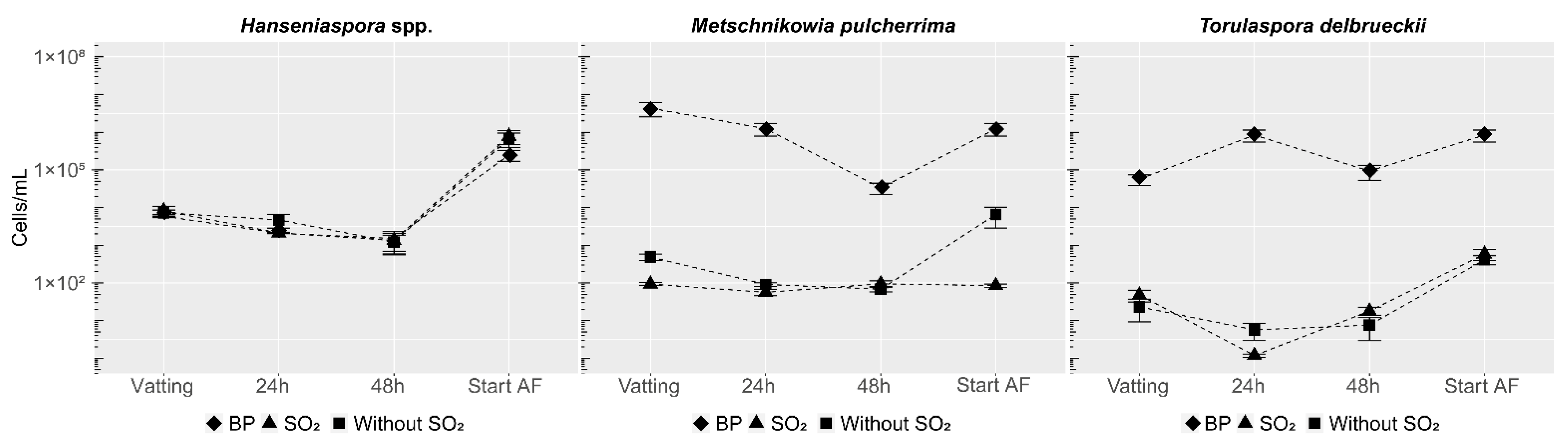
3.2.1. Population Dynamics of Hanseniaspora spp. and Non-Saccharomyces Yeasts Used as Bioprotection
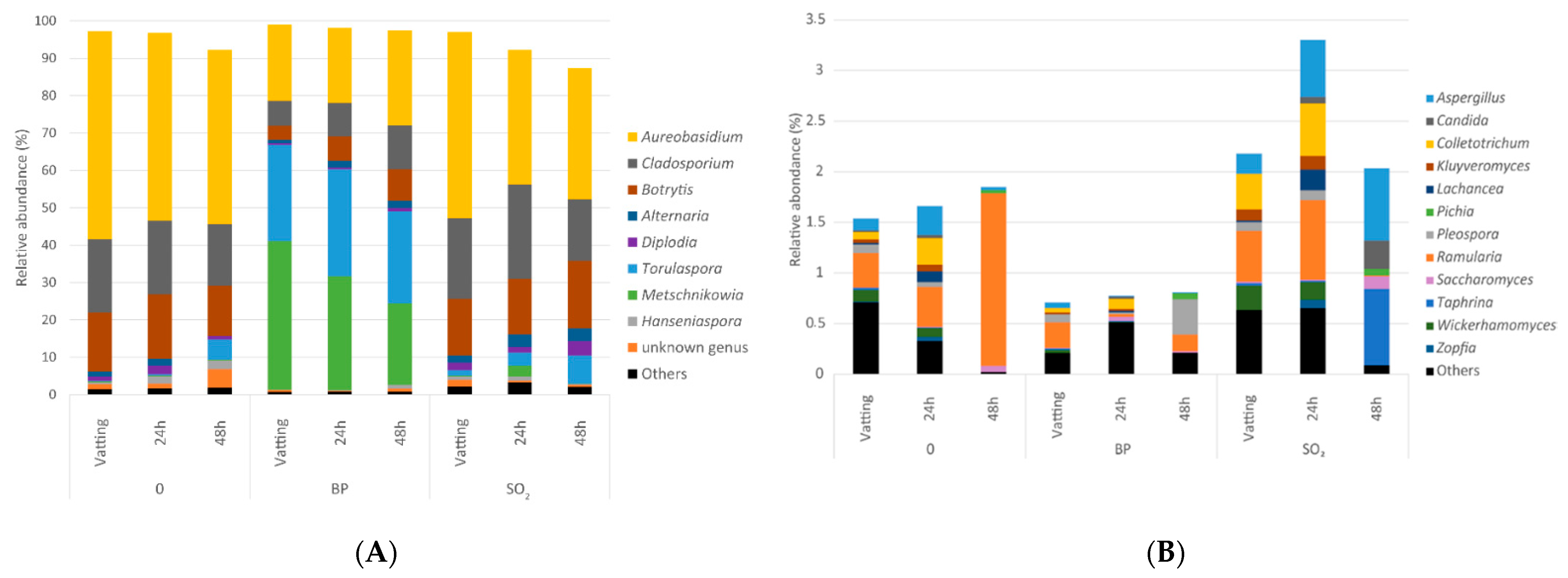
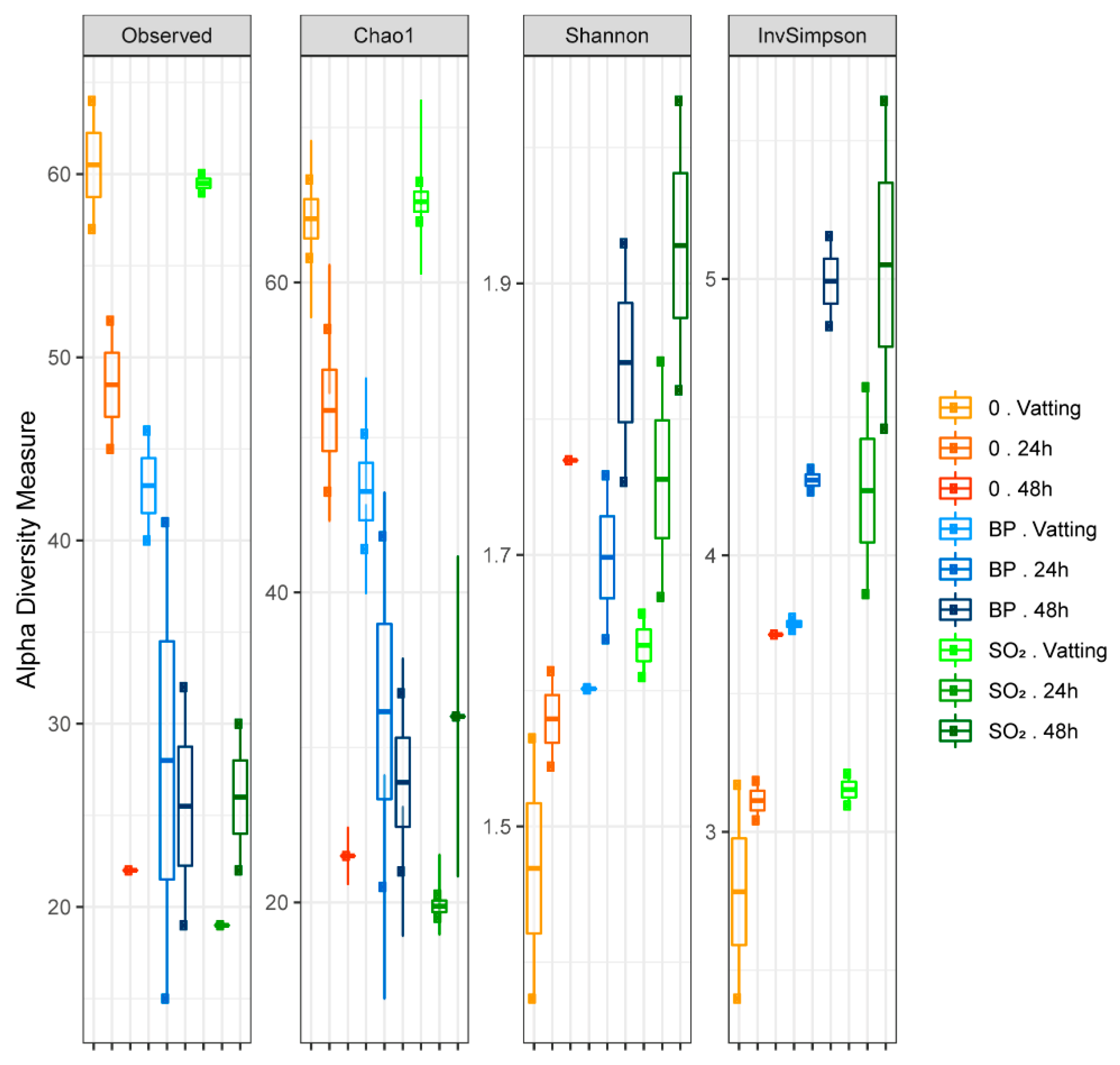
3.2.2. Yeasts and Fungal Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Name CRB | GenBank Accession Number(s) | Species Literature References |
|---|---|---|---|---|
| Brettanomyces | acidodurans | NCAIM Y 2178 | [73] | |
| Zygosaccharomyces | bailii | L0536 | MT950295 | [74] |
| Brettanomyces | bruxellensis | CRBO_L0308 | MT950279 | [46] |
| Brettanomyces | bruxellensis | CRBO_L0417 | MT950285 | |
| Brettanomyces | bruxellensis | CRBO_L0422 | MT950286 | |
| Brettanomyces | bruxellensis | CRBO_L0424 | MT950287 | |
| Brettanomyces | bruxellensis | CRBO_L0463 | MT950293 | |
| Brettanomyces | bruxellensis | CRBO_L0512 | MT950294 | |
| Brettanomyces | bruxellensis | CRBO_L0542 | MT950296 | |
| Brettanomyces | bruxellensis | CRBO_L0611 | MT950299 | |
| Brettanomyces | bruxellensis | CRBO_L14156 | MT950310 | |
| Brettanomyces | bruxellensis | CRBO_L14169 | MT950311 | |
| Brettanomyces | bruxellensis | CRBO_L14173 | MT950312 | |
| Brettanomyces | bruxellensis | CRBO_L14195 | MT950313 | |
| Brettanomyces | bruxellensis | CRBO_L1735 | ||
| Brettanomyces | bruxellensis | CRBO_L1750 | ||
| Brettanomyces | bruxellensis | CRBO_L1774 | ||
| Candida | cantarelli | CRBO_L0404 | MT950283 | |
| Candida | cantarelli | CRBO_L0412 | MT950284 | |
| Saccharomyces | cerevisiae | 522D | [75] | |
| Saccharomyces | cerevisiae | CRBO_L0431 | MT950288 | |
| Saccharomyces | cerevisiae | CRBO_L0439 | MT950289 | |
| Saccharomyces | cerevisiae | CRBO_L0545 | MT950298 | |
| Saccharomyces | cerevisiae | CRBO_L1117 | MT950308 | |
| Torulaspora | delbrueckii | CRBO_L0544 | MT950297 | [76] |
| Torulaspora | delbrueckii | CRBO_L0630 | MT950300 | |
| Pichia | guiliermondii | CRBO_L0652 | MT950302 | [77] |
| Shizosaccharomyces | japonicus | Y13611 | ||
| Pichia | kluyveri | CRBO_L0677 | MT950304 | |
| Pichia | membranifaciens | CRBO_L0709 | MT950305 | |
| Schizosaccharomyces | octosporus | Y-8551 | ||
| Schizosaccharomyces | pombe | CRBO_L0442 | MT950290 | [78] |
| Schizosaccharomyces | pombe | CRBO_L0443 | MT950291 | |
| Schizosaccharomyces | pombe | Y12791 | ||
| Metschnikowia | pulcherima | CRBO_L0313 | MT950282 | [79] |
| Metschnikowia | pulcherima | CRBO_L0640 | MT950301 | |
| Aureobasidium | pullulans | CRBO_L0448 | MT950292 | [80] |
| Aureobasidium | pullulans | CRBO_L11178 | MT950309 | |
| Lanchancea | thermotolerans | CRBO_L0672 | MT950303 | [81] |
| Hanseniaspora | uvarum | CRBO_L0312 | MT950281 | [82] |
| Hanseniaspora | uvarum | CRBO_L0715 | MT950306 | |
| Starmerella | bacillaris | CRBO_L0311 | MT950280 | [83] |
| Starmerella | bacillaris | CRBO_L0740 | MT950307 |
| Species | Primers | References |
|---|---|---|
| Metschnikowia pulcherrima | MP2-F AGACACTTAACTGGGCCAGC MP2-R GGGGTGGTGTGGAAGTAAGG | [16] |
| Torulaspora delbrueckii | TD-F CAAAGTCATCCAAGCCAGC TD-R TTCTCAAACAATCATGTTTGGTAG | [7] |
| Hanseniaspora spp. | Hauf 2L—CCCTTTGCCTAAGGTACG Hauf 2R—CGCTGTTCTCGCTGTGATG | [7] |
| Chemical Composition in Must | In Wine | |||||
|---|---|---|---|---|---|---|
| Parameters | SO2 | 0 | BP | SO2 | 0 | BP |
| Reducing sugars (g/L) | 245 | 242 | 243 | 1.0 | 3.0 | 1.4 |
| Total acidity (g/L) | 1.63 | 1.71 | 1.67 | |||
| Malic acid (g/L) | 1.1 | 1.0 | 1.1 | |||
| pH | 3.84 | 3.84 | 3.83 | |||
| Yeats assimilable nitrogen (mg/L) | 116 | 124 | 111 | |||
| Total SO2 (mg/L) | 127 | <10 | <10 | |||
| Volatil acidity (acetic acid g/L) | 0.28 | 0.30 | 0.28 | |||
| Stage | 1. Vatting | 2. 24 h of Maceration | 3. 48 h of Maceration | 4. Start of AF | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | 0 | SO2 | BP | 0 | SO2 | BP | 0 | SO2 | BP | 0 | SO2 | BP | |||||||||||||
| Duplicats | a | b | a | b | a | b | a | b | a | b | a | b | a | b | a | b | a | b | a | b | a | b | a | b | |
| HTS metabarcoding (Relative abundance by Genus in %) | Aureobasidium | 63.2 | 48.3 | 49.7 | 50.1 | 20.3 | 20.3 | 49.7 | 50.7 | 41.2 | 30.7 | 13.9 | 26.3 | 46.5 | - | 31.7 | 38.4 | 21.9 | 29.0 | - | - | - | - | - | - |
| Cladosporium | 18.7 | 20.5 | 20.2 | 22.9 | 7.0 | 6.5 | 18.7 | 20.8 | 24.2 | 26.5 | 10.9 | 6.9 | 16.6 | - | 17.0 | 15.9 | 7.1 | 16.3 | - | - | - | - | - | - | |
| Botrytis | 11.1 | 20.4 | 15.8 | 14.5 | 3.6 | 3.9 | 17.7 | 16.8 | 15.1 | 14.5 | 8.7 | 4.3 | 13.4 | - | 16.4 | 19.7 | 9.2 | 7.8 | - | - | - | - | - | - | |
| Alternaria | 0.6 | 1.9 | 1.6 | 2.3 | 0.8 | 1.0 | 1.5 | 2.1 | 5.2 | 1.6 | 1.5 | 2.2 | - | - | 4.3 | 2.6 | 2.6 | 1.3 | - | - | - | - | - | - | |
| Diplodia | 0.9 | 1.6 | 2.0 | 2.0 | 0.6 | 0.4 | 2.0 | 2.7 | 0.1 | 2.9 | 0.1 | 0.9 | 0.9 | - | 1.4 | 6.2 | 1.1 | 0.7 | - | - | - | - | - | - | |
| Torulaspora | 0.3 | 0.2 | 2.7 | 0.4 | 24.8 | 26.6 | 0.5 | 0.2 | 4.0 | 3.0 | 33.4 | 24.0 | 5.5 | - | 7.5 | 7.7 | 24.1 | 25.2 | - | - | - | - | - | - | |
| Metschnikowia | 0.1 | 0.2 | 0.1 | 0.1 | 40.5 | 39.0 | 0.3 | 0.2 | 5.7 | 0.1 | 29.4 | 31.7 | 0.1 | - | 0.0 | 0.1 | 27.3 | 16.2 | - | - | - | - | - | - | |
| Hanseniaspora | 0.4 | 0.5 | 1.2 | 0.7 | 0.1 | 0.1 | 3.8 | 0.1 | 0.2 | 2.0 | 0.1 | 0.1 | 2.4 | - | 0.2 | 0.2 | 0.2 | 1.9 | - | - | - | - | - | - | |
| unknown genus | 1.1 | 1.4 | 1.9 | 1.7 | 0.5 | 0.5 | 1.1 | 1.4 | 1.0 | 0.0 | 0.0 | 0.5 | 4.9 | - | 1.3 | 0.0 | 1.6 | 0.1 | - | - | - | - | - | - | |
| others | 1.7 | 1.4 | 1.9 | 2.5 | 0.8 | 0.6 | 1.7 | 1.7 | 0.8 | 5.8 | 0.2 | 1.4 | 1.9 | - | 3.1 | 1.0 | 0.8 | 0.8 | - | - | - | - | - | - | |
| Q-PCR (Cells/mL) | Torulaspora delbrueckii | <100 | <100 | <100 | <100 | 4.5 × 104 | 6.8 × 104 | <100 | <100 | <100 | <100 | 1.2 × 106 | 4.8 × 105 | <100 | <100 | <100 | <100 | 7.7 × 104 | 1.1 × 105 | 3.7 × 102 | 4.6 × 102 | 8.8 × 102 | 2.9 × 102 | 1.2 × 104 | 4.8 × 105 |
| Metschnikowia pulcherrima | 3.9 × 102 | 5.8 × 102 | <100 | <100 | 4.5 × 106 | 4.3 × 106 | <100 | <100 | <100 | <100 | 1.6 × 106 | 8.4 × 105 | <100 | <100 | 1.1 × 102 | <100 | 3.8 × 104 | 2.9 × 104 | 2.0 × 102 | 1.3 × 104 | <100 | <100 | 1.6 × 106 | 8.4 × 105 | |
| Hanseniaspora sp. | 9.2 × 103 | 5.4 × 103 | 1.2 × 104 | 4.6 × 103 | 5.2 × 103 | 6.7 × 103 | 7.4 × 103 | 2.1 × 103 | 2.3 × 103 | 2.0 × 103 | 2.1 × 103 | 2.0 × 103 | 2.9 × 102 | 2.1 × 103 | 2.8 × 102 | 2.4 × 103 | 3.4 × 102 | 2.7 × 103 | 2.8 × 105 | 1.1 × 106 | 1.2 × 106 | 3.4 × 105 | 3.6 × 105 | 1.4 × 105 | |
| Culture | Total Yeasts (UFC/mL) | 1.2 × 103 | 2.3 × 103 | 8.3 × 102 | 1.3 × 103 | 4.8 × 105 | 3.4 × 105 | 2.4 × 103 | 2.5 × 103 | 2.0 × 103 | 1.6 × 103 | 5.9 × 105 | 4.0 × 103 | 1.2 × 103 | 4.0 × 103 | 1.4 × 103 | 9.8 × 102 | 5.5 × 105 | 3.4 × 106 | 2.0 × 107 | 2.1 × 107 | 7.7 × 106 | 1.8 × 107 | 2.6 × 107 | 4.0 × 107 |
| MALDI-TOF MS (%) | TOTAL number of colonies analyzed | 30 | 30 | 30 | 30 | 30 | 30 | 23 | 30 | 24 | 30 | 30 | 30 | 30 | 6 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 | 30 |
| Torulaspora delbruekii | - | - | - | - | 56.7 | 60.0 | - | - | 4.2 | 10.0 | 90.0 | 50.0 | 3.3 | - | 16.7 | 10.0 | 86.7 | 66.7 | - | - | - | - | - | - | |
| Metschnikowia pulcherrima | - | 26.7 | - | 3.3 | 40.0 | 40.0 | 13.0 | 33.3 | - | 3.3 | 10.0 | 40.0 | 3.3 | 16.7 | 3.3 | 6.7 | 10.0 | 33.3 | - | - | - | - | - | - | |
| Hanseniaspora uvarum | 40.0 | 36.7 | 13.3 | 46.7 | - | - | 34.8 | 43.3 | 58.3 | 33.3 | - | - | 16.7 | - | 43.3 | 30.0 | - | - | 3.0 | - | 3.0 | - | - | - | |
| Aureobasidium pullulans | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| Kluveromyces lactis | 13.3 | 10.0 | 3.3 | 13.3 | - | - | 8.7 | - | 12.5 | 20.0 | - | - | 33.3 | - | 10.0 | 10.0 | - | - | - | - | - | - | - | - | |
| Saccharomyces cerevisiae | 13.3 | 3.3 | 36.7 | 10.0 | - | - | 4.3 | - | - | - | - | - | - | - | - | 10.0 | - | - | 97 | 100 | 97 | 100 | 100 | 100 | |
| Candida guillermondii | - | - | - | - | - | - | 4.3 | - | - | - | - | - | - | 16.7 | - | - | - | - | - | - | - | - | - | - | |
| Lachancea thermotolerans | - | - | - | - | - | - | - | - | - | - | - | - | - | 16.7 | 3.3 | 3.3 | - | - | - | - | - | - | - | - | |
| Cryptococcus flavescens | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| Whickerhamomyces anomalus | 33.3 | 23.3 | 46.7 | 26.7 | - | - | 21.7 | 16.7 | 12.5 | 20.0 | - | - | 26.7 | - | 23.3 | 26.7 | - | - | - | - | - | - | - | - | |
| No ID | - | - | - | - | 3.3 | - | 13.0 | 6.7 | 12.5 | 13.3 | - | 10.0 | 16.7 | 50.0 | - | 3.3 | 3.3 | - | - | - | - | - | - | - | |
References
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. PNAS Plus: From the Cover: Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Nat. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [Green Version]
- Albertin, W.; Miot-Sertier, C.; Bely, M.; Marullo, P.; Coulon, J.; Moine, V.; Colonna-Ceccaldi, B.; Masneuf-Pomarede, I. Oenological prefermentation practices strongly impact yeast population dynamics and alcoholic fermentation kinetics in Chardonnay grape must. Int. J. Food Microbiol. 2014, 178, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Chasseriaud, L.; Coulon, J.; Marullo, P.; Albertin, W.; Bely, M. New oenological practice to promote non-Saccharomyces species of interest: Saturating grape juice with carbon dioxide. Appl. Microbiol. Biotechnol. 2018, 102, 3779–3791. [Google Scholar] [CrossRef]
- Zott, K.; Miot-Sertier, C.; Claisse, O.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Dynamics and diversity of non-Saccharomyces yeasts during the early stages in winemaking. Int. J. Food Microbiol. 2008, 125, 197–203. [Google Scholar] [CrossRef]
- Lleixà, J.; Manzano, M.; Mas, A.; Portillo, M.D.C. Saccharomyces and non-Saccharomyces Competition during Microvinification under Different Sugar and Nitrogen Conditions. Front. Microbiol. 2016, 7, 1959. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; García-Fernández, D.; Mas, A.; Esteve-Zarzoso, B. Fungal diversity in grape must and wine fermentation assessed by massive sequencing, quantitative PCR and DGGE. Front. Microbiol. 2015, 6, 1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zott, K.; Claisse, O.; Lucas, P.; Coulon, J.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Characterization of the yeast ecosystem in grape must and wine using real-time PCR. Food Microbiol. 2010, 27, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Swadener, M.; Sakamoto, K.; Mills, D.A.; Bisson, L.F. Sulfur Dioxide Treatment Alters Wine Microbial Diversity and Fermentation Progression in a Dose-Dependent Fashion. Am. J. Enol. Vitic. 2014, 66, 73–79. [Google Scholar] [CrossRef]
- Morgan, S.C.; McCarthy, G.C.; Watters, B.S.; Tantikachornkiat, M.; Zigg, I.; Cliff, M.A.; Durall, D.M. Effect of sulfite addition and pied de cuve inoculation on the microbial communities and sensory profiles of Chardonnay wines: Dominance of indigenous Saccharomyces uvarum at a commercial winery. FEMS Yeast Res. 2019, 19, 49. [Google Scholar] [CrossRef]
- Morgan, S.C.; Tantikachornkiat, M.; Scholl, C.M.; Benson, N.L.; Cliff, M.A.; Durall, D.M. The effect of sulfur dioxide addition at crush on the fungal and bacterial communities and the sensory attributes of Pinot gris wines. Int. J. Food Microbiol. 2019, 290, 1–14. [Google Scholar] [CrossRef]
- Constantí, M.; Reguant, C.; Poblet, M.; Zamora, F.; Mas, A.; Guillamón, J.M. Molecular Analysis of Yeast Population Dynamics: Effect of Sulphur Dioxide and Inoculum on Must Fermentation. Int. J. Food Microbiol. 1998, 41, 169–175. [Google Scholar] [CrossRef]
- Henick-Kling, T.; Edinger, W.; Daniel, P.; Monk, P. Selective Effects of Sulfur Dioxide and Yeast Starter Culture Addition on Indigenous Yeast Populations and Sensory Characteristics of Wine. J. Appl. Microbiol. 1998, 84, 865–876. [Google Scholar] [CrossRef]
- Morgan, S.C.; Scholl, C.M.; Benson, N.L.; Stone, M.L.; Durall, D.M. Sulfur dioxide addition at crush alters Saccharomyces cerevisiae strain composition in spontaneous fermentations at two Canadian wineries. Int. J. Food Microbiol. 2017, 244, 96–102. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-Protection as an Alternative to Sulphites: Impact on Chemical and Microbial Characteristics of Red Wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Esteve-Zarzoso, B.; Crespo, J.; Cabellos, J.M.; Arroyo, T. Yeast Monitoring of Wine Mixed or Sequential Fermentations Made by Native Strains from D.O. “Vinos de Madrid” Using Real-Time Quantitative PCR. Front. Microbiol. 2017, 8, 2520. [Google Scholar] [CrossRef]
- Andorrà, I.; Landi, S.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of Oenological Practices on Microbial Populations Using Culture-Independent Techniques. Food Microbiol. 2008, 25, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Sirén, K.; Mak, S.S.T.; Fischer, U.; Hansen, L.H.; Gilbert, M.T.P. Multi-omics and potential applications in wine production. Curr. Opin. Biotechnol. 2019, 56, 172–178. [Google Scholar] [CrossRef]
- David, V.; Terrat, S.; Herzine, K.; Claisse, O.; Rousseaux, S.; Tourdot-Maréchal, R.; Masneuf-Pomarede, I.; Ranjard, L.; Alexandre, H. High-throughput sequencing of amplicons for monitoring yeast biodiversity in must and during alcoholic fermentation. J. Ind. Microbiol. Biotechnol. 2014, 41, 811–821. [Google Scholar] [CrossRef]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.P.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [Green Version]
- Heather, J.M.; Chain, B. The sequence of sequencers: The history of sequencing DNA. Genomics 2016, 107, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry: A Fundamental Shift in the Routine Practice of Clinical Microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferroni, A.; Suarez, S.; Beretti, J.-L.; Dauphin, B.; Bille, E.; Meyer, J.; Bougnoux, M.-E.; Alanio, A.; Berche, P.; Nassif, X. Real-Time Identification of Bacteria and Candida Species in Positive Blood Culture Broths by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. J. Clin. Microbiol. 2010, 48, 1542–1548. [Google Scholar] [CrossRef] [Green Version]
- Akimowicz, M.; Bucka-Kolendo, J. MALDI-TOF MS–Application in Food Microbiology. Acta Biochim. Pol. 2020, 67, 327–332. [Google Scholar] [CrossRef]
- Turvey, M.E.; Weiland, F.; Meneses, J.; Sterenberg, N.; Hoffmann, P. Identification of beer spoilage microorganisms using the MALDI Biotyper platform. Appl. Microbiol. Biotechnol. 2016, 100, 2761–2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usbeck, J.C.; Kern, C.C.; Vogel, R.F.; Behr, J. Optimization of experimental and modelling parameters for the differentiation of beverage spoiling yeasts by Matrix-Assisted-Laser-Desorption/Ionization–Time-of-Flight Mass Spectrometry (MALDI–TOF MS) in response to varying growth conditions. Food Microbiol. 2013, 36, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Wieme, A.D.; Spitaels, F.; Aerts, M.; De Bruyne, K.; Van Landschoot, A.; Vandamme, P. Identification of beer-spoilage bacteria using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Int. J. Food Microbiol. 2014, 185, 41–50. [Google Scholar] [CrossRef]
- Gutiérrez, C.; Flechoso, M.D.L.; Ángeles, G.; Belda, I.; Ruiz, J.; Kayali, N.; Polo, L.; Santos, A. Wine yeasts identification by MALDI-TOF MS: Optimization of the preanalytical steps and development of an extensible open-source platform for processing and analysis of an in-house MS database. Int. J. Food Microbiol. 2017, 254, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Plowman, J.E.; Tian, B.; Clerens, S.; On, S.L. An improved method for MALDI-TOF analysis of wine-associated yeasts. J. Microbiol. Methods 2020, 172, 105904. [Google Scholar] [CrossRef] [PubMed]
- Blättel, V.; Petri, A.; Rabenstein, A.; Kuever, J.; König, H. Differentiation of species of the genus Saccharomyces using biomolecular fingerprinting methods. Appl. Microbiol. Biotechnol. 2013, 97, 4597–4606. [Google Scholar] [CrossRef] [PubMed]
- Moothoo-Padayachie, A.; Kandappa, H.R.; Krishna, S.B.N.; Maier, T.; Govender, P. Biotyping Saccharomyces cerevisiae strains using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Eur. Food Res. Technol. 2013, 236, 351–364. [Google Scholar] [CrossRef]
- Usbeck, J.C.; Wilde, C.; Bertrand, D.; Behr, J.; Vogel, R.F. Wine yeast typing by MALDI-TOF MS. Appl. Microbiol. Biotechnol. 2014, 98, 3737–3752. [Google Scholar] [CrossRef] [PubMed]
- Kačániová, M.; Kunova, S.; Felsöciová, S.; Ivanišová, E.; Kántor, A.; Puchalski, C.; Terentjeva, M. Microbiota of different wine grape berries. Potravin. Slovak J. Food Sci. 2019, 13, 174–181. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Hoff, J.; Hart, R.; Ndimba, B.; Jolly, N. Characterisation of Non-Saccharomyces Yeasts Using Different Methodologies and Evaluation of their Compatibility with Malolactic Fermentation. S. Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Lücke, F.-K. Utilization of microbes to process and preserve meat. Meat Sci. 2000, 56, 105–115. [Google Scholar] [CrossRef]
- Stiles, M.E. Biopreservation by lactic acid bacteria. Antonie Leeuwenhoek 1996, 70, 331–345. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Identification of clinically important ascomycetous yeasts based on nucleotide divergence in the 5’ end of the large-subunit (26S) ribosomal DNA gene. J. Clin. Microbiol. 1997, 35, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Pavlovic, M.; Mewes, A.; Maggipinto, M.; Schmidt, W.; Messelhäußer, U.; Balsliemke, J.; Hörmansdorfer, S.; Busch, U.; Huber, I. MALDI-TOF MS Based Identification of Food-Borne Yeast Isolates. J. Microbiol. Methods 2014, 106, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Bilecen, K.; Yaman, G.; Ciftci, U.; Laleli, Y.R. Performances and Reliability of Bruker Microflex LT and VITEK MS MALDI-TOF Mass Spectrometry Systems for the Identification of Clinical Microorganisms. BioMed Res. Int. 2015, 2015, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Condina, M.R.; Dilmetz, B.A.; Bazaz, S.R.; Meneses, J.; Warkiani, M.E.; Hoffmann, P. Rapid separation and identification of beer spoilage bacteria by inertial microfluidics and MALDI-TOF mass spectrometry. Lab Chip 2019, 19, 1961–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; De Vargas, C.; Dunthorn, M. Swarm v2: Highly-scalable and high-resolution amplicon clustering. PeerJ 2015, 3, e1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avramova, M.; Cibrario, A.; Peltier, E.; Coton, M.; Coton, E.; Schacherer, J.; Spano, G.; Capozzi, V.; Blaiotta, G.; Salin, F.; et al. Brettanomyces bruxellensis population survey reveals a diploid-triploid complex structured according to substrate of isolation and geographical distribution. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef]
- Martins, G.M. Communautés Microbiennes de La Baie de Raisin: Incidence Des Facteurs Biotiques et Abiotiques: Incidence Des Facteurs Biotiques et Abiotiques. Communautés Microbiennes de la Baie de Raisin: Incidence des Facteurs Biotiques et Abiotiques. Ph.D. Thesis, Université de Bordeaux Ségalen, Bordeaux, France, 2012. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Evol. Microbiol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Granchi, L.; Bosco, M.; Messini, A.; Vincenzini, M. Rapid detection and quantification of yeast species during spontaneous wine fermentation by PCR-RFLP analysis of the rDNA ITS region. J. Appl. Microbiol. 1999, 87, 949–956. [Google Scholar] [CrossRef]
- Prakitchaiwattana, C.J.; Fleet, G.H.; Heard, G.M. Application and evaluation of denaturing gradient gel electrophoresis to analyse the yeast ecology of wine grapes. FEMS Yeast Res. 2004, 4, 865–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agustini, B.C.; Silva, L.P.; Bloch, C.; Bonfim, T.M.B.; Da Silva, G.A. Evaluation of MALDI-TOF mass spectrometry for identification of environmental yeasts and development of supplementary database. Appl. Microbiol. Biotechnol. 2014, 98, 5645–5654. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.-A.; Bálint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Leray, M.; Knowlton, N. Random sampling causes the low reproducibility of rare eukaryotic OTUs in Illumina COI metabarcoding. PeerJ 2017, 5, e3006. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, B.; Bauer, F.F.; Cardinali, G.; Setati, M.E. Ecological interactions are a primary driver of population dynamics in wine yeast microbiota during fermentation. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Colabella, C.; Corte, L.; Roscini, L.; Bassetti, M.; Tascini, C.; Mellor, J.C.; Meyer, W.; Robert, V.; Vu, D.; Cardinali, G. NGS barcode sequencing in taxonomy and diagnostics, an application in “Candida” pathogenic yeasts with a metagenomic perspective. IMA Fungus 2018, 9, 91–105. [Google Scholar] [CrossRef]
- Stefanini, I.; Cavalieri, D. Metagenomic Approaches to Investigate the Contribution of the Vineyard Environment to the Quality of Wine Fermentation: Potentials and Difficulties. Front. Microbiol. 2018, 9, 991. [Google Scholar] [CrossRef] [PubMed]
- Işçi, B.; Yıldırım, H.K.; Altindişli, A. Evaluation of methods for DNA extraction from must and wine. J. Inst. Brew. 2014, 120, 238–243. [Google Scholar] [CrossRef]
- Belda, I.; Zarraonaindia, I.; Perisin, M.; Palacios, A.; Acedo, A. From Vineyard Soil to Wine Fermentation: Microbiome Approximations to Explain the “Terroir” Concept. Front. Microbiol. 2017, 8, 821. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.M.; Portillo, M.C.; Belda-Ferre, P.; Mira, A. Amplification by PCR Artificially Reduces the Proportion of the Rare Biosphere in Microbial Communities. PLoS ONE 2012, 7, e29973. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among Wine Grape Microbiome, Metabolome, and Fermentation Behavior Suggest Microbial Contribution to Regional Wine Characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [Green Version]
- Boynton, P.J.; Greig, D. Species richness influences wine ecosystem function through a dominant species. Fungal Ecol. 2016, 22, 61–72. [Google Scholar] [CrossRef] [Green Version]
- De Souza, R.S.C.; Okura, V.K.; Armanhi, J.S.L.; Jorrín, B.; Lozano, N.; Da Silva, M.J.; González-Guerrero, M.; De Araújo, L.M.; Verza, N.C.; Bagheri, H.C.; et al. Unlocking the bacterial and fungal communities assemblages of sugarcane microbiome. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, N.; Schneider, D.; Sahner, J.; Ballauff, J.; Edy, N.; Barus, H.; Irawan, B.; Budi, S.W.; Qaim, M.; Daniel, R.; et al. Intensive tropical land use massively shifts soil fungal communities. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Minervini, F.; Lattanzi, A.; De Angelis, M.; Celano, G.; Gobbetti, M. House microbiotas as sources of lactic acid bacteria and yeasts in traditional Italian sourdoughs. Food Microbiol. 2015, 52, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Sirén, K.; Mak, S.S.T.; Melkonian, C.; Carøe, C.; Swiegers, J.H.; Molenaar, D.; Fischer, U.; Gilbert, M.T.P. Taxonomic and Functional Characterization of the Microbial Community During Spontaneous in vitro Fermentation of Riesling Must. Front. Microbiol. 2019, 10, 697. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Khelaifia, S.; Alou, M.T.; Ndongo, S.; Dione, N.; Hugon, P.; Caputo, A.; Cadoret, F.; Traore, S.I.; Seck, E.H.; et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat. Microbiol. 2016, 1, 1–8. [Google Scholar] [CrossRef]
- Sarhan, M.S.; Hamza, M.A.; Youssef, H.H.; Patz, S.; Becker, M.; ElSawey, H.; Nemr, R.; Daanaa, H.-S.A.; Mourad, E.F.; Morsi, A.T.; et al. Culturomics of the plant prokaryotic microbiome and the dawn of plant-based culture media—A review. J. Adv. Res. 2019, 19, 15–27. [Google Scholar] [CrossRef]
- Mourad, E.F.; Sarhan, M.; Daanaa, H.-S.A.; Abdou, M.; Morsi, A.T.; Abdelfadeel, M.R.; Elsawey, H.; Nemr, R.; El-Tahan, M.; Hamza, M.A.; et al. Plant Materials are Sustainable Substrates Supporting New Technologies of Plant-Only-Based Culture Media for in vitro Culturing of the Plant Microbiota. Microbes Environ. 2018, 33, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Picciotti, G. The growth kinetics and fermentation behaviour of some non-Saccharomyces yeasts associated with wine-making. Biotechnol. Lett. 1995, 17, 1247–1250. [Google Scholar] [CrossRef]
- Comi, G.; Romano, P.; Cocolin, L.; Fiore, C. Characterization of Kloeckera apiculata strains from the Friuli regionin Northern Italy. World J. Microbiol. Biotechnol. 2001, 17, 391–394. [Google Scholar] [CrossRef]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of Yeast Species and Strains in Wine Flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Péter, G.; Dlauchy, D.; Tóbiás, A.; Fülöp, L.; Podgoršek, M.; Čadež, N. Brettanomyces Acidodurans Sp. Nov., a New Acetic Acid Producing Yeast Species from Olive Oil. Antonie Leeuwenhoek 2017, 110, 657–664. [Google Scholar] [CrossRef]
- Schuller, D.; Côrte-Real, M.; Leão, C. A Differential Medium for the Enumeration of the Spoilage Yeast Zygosaccharomyces Bailii in Wine. J. Food Prot. 2000, 63, 1570–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.-L.; Masneuf-Pomarede, I. Cellar-Associated Saccharomyces Cerevisiae Population Structure Revealed High-Level Diversity and Perennial Persistence at Sauternes Wine Estates. Appl. Environ. Microbiol. 2016, 82, 2909–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertin, W.; Chasseriaud, L.; Comte, G.; Panfili, A.; Delcamp, A.; Salin, F.; Marullo, P.; Bely, M. Winemaking and Bioprocesses Strongly Shaped the Genetic Diversity of the Ubiquitous Yeast Torulaspora Delbrueckii. PLoS ONE 2014, 9, e94246. [Google Scholar] [CrossRef]
- Jensen, S.L.; Umiker, N.L.; Arneborg, N.; Edwards, C.G. Identification and Characterization of Dekkera Bruxellensis, Candida Pararugosa, and Pichia Guilliermondii Isolated from Commercial Red Wines. Food Microbiol. 2009, 26, 915–921. [Google Scholar] [CrossRef] [PubMed]
- Benito, Á.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.-Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces Pombe Strains Have Characteristics That Are Beneficial for Winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef] [PubMed]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial Activity of M Etschnikowia Pulcherrima on Wine Yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Onetto, C.A.; Borneman, A.R.; Schmidt, S.A. Investigating the Effects of Aureobasidium Pullulans on Grape Juice Composition and Fermentation. Food Microbiol. 2020, 90, 103451. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Schmidtke, L.; Boss, P.K.; Grbin, P.R.; Masneuf-Pomarede, I.; Bely, M.; Albertin, W.; Jiranek, V. Oenological Traits of Lachancea Thermotolerans Show Signs of Domestication and Allopatric Differentiation. Sci. Rep. 2018, 8, 14812. [Google Scholar] [CrossRef]
- Albertin, W.; Setati, M.E.; Miot-Sertier, C.; Mostert, T.T.; Colonna-Ceccaldi, B.; Coulon, J.; Girard, P.; Moine, V.; Pillet, M.; Salin, F.; et al. Hanseniaspora Uvarum from Winemaking Environments Show Spatial and Temporal Genetic Clustering. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella Bacillaris in Winemaking: Opportunities and Risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]





| Genus | Species | Number of Strains in Biotyper DB | Number of Strains in OLS DB |
|---|---|---|---|
| Aureobasidium | pullulans | 3 | 2 |
| Brettanomyces | acidodurans | 0 | 1 |
| Brettanomyces | bruxellensis | 5 | 15 |
| Candida | cantarelli | 0 | 2 |
| Starmerella | bacillaris | 0 | 2 |
| Hanseniaspora | uvarum | 8 | 2 |
| Lanchancea | thermotolerans | 3 | 1 |
| Metschnikowia | pulcherima | 4 | 2 |
| Pichia (Candida) | guiliermondii | 14 | 1 |
| Pichia | kluyveri | 1 | 1 |
| Pichia (Candida) | membranifaciens | 2 | 1 |
| Saccharomyces | cerevisiae | 13 | 5 |
| Shizosaccharomyces | japonicus | 0 | 1 |
| Shizosaccharomyces | octosporus | 0 | 1 |
| Shizosaccharomyces | pombe | 4 | 3 |
| Torulaspora | delbrueckii | 5 | 2 |
| Zygosaccharomyces | bailii | 3 | 1 |
| Observed | Chao1 | Shannon | InvSimpson | |
|---|---|---|---|---|
| Stage | 52.70% ** | 53.29% ** | 44.90% * | 53.96% ** |
| Treatment | 19.13% * | 11.25% | 13.77% | 22.79% * |
| Treatment*Stage | 14.48% | 19.67% | 4.90% | 6.19% |
| Residuals | 13.64% | 15.78% | 36.33% | 17.05% |
| Stage | Treatment | Shannon |
|---|---|---|
| Vatting | 0 | 1.34 |
| SO2 | 1.21 | |
| BP | 0.68 | |
| 24 h of maceration | 0 | 1.24 |
| SO2 | 1.16 | |
| BP | 0.52 | |
| 48 h of maceration | 0 | 1.24 |
| SO2 | 1.58 | |
| BP | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Windholtz, S.; Dutilh, L.; Lucas, M.; Maupeu, J.; Vallet-Courbin, A.; Farris, L.; Coulon, J.; Masneuf-Pomarède, I. Population Dynamics and Yeast Diversity in Early Winemaking Stages without Sulfites Revealed by Three Complementary Approaches. Appl. Sci. 2021, 11, 2494. https://doi.org/10.3390/app11062494
Windholtz S, Dutilh L, Lucas M, Maupeu J, Vallet-Courbin A, Farris L, Coulon J, Masneuf-Pomarède I. Population Dynamics and Yeast Diversity in Early Winemaking Stages without Sulfites Revealed by Three Complementary Approaches. Applied Sciences. 2021; 11(6):2494. https://doi.org/10.3390/app11062494
Chicago/Turabian StyleWindholtz, Sara, Lucie Dutilh, Marine Lucas, Julie Maupeu, Amélie Vallet-Courbin, Laura Farris, Joana Coulon, and Isabelle Masneuf-Pomarède. 2021. "Population Dynamics and Yeast Diversity in Early Winemaking Stages without Sulfites Revealed by Three Complementary Approaches" Applied Sciences 11, no. 6: 2494. https://doi.org/10.3390/app11062494