Recent Advancements of UF-Based Separation for Selective Enrichment of Proteins and Bioactive Peptides—A Review

 ,
,
Abstract
:Featured Application
Abstract
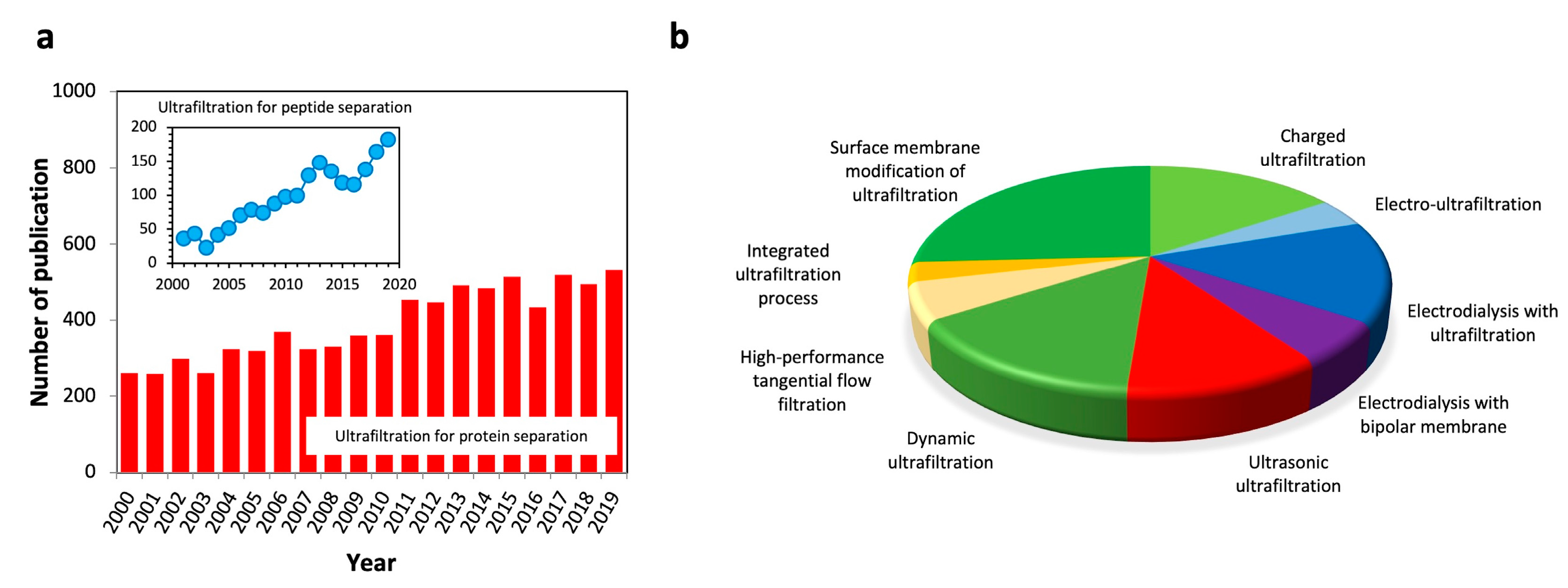
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Hydrolysis Enzyme(s) | Protocols | Bioactivity | Target Compound (s) | Remarks | Ref |
|---|---|---|---|---|---|---|
| Grateloupia turuturu (Macroalgae) | - | Preindustrial UF PCI-MT600 (Polyethersulfone (PES) 30 kDa) from | -Antioxidant | R-phycoerythrin | 100% recovery rate flux: 35.1 L m−2 h−1 | [34] |
| Enteromorpha clathrata (Macroalgae) | Alcalase | QuixStandTM Benchtop System (10 kDa) | -ACE inhibitors | Peptide fragments: PAFG | Yield: 0.82% IC50: 35.9 μM | [35] |
| Pigeon pea | Pepsin–pancreatin, Alcalase, or Pancreatin | Sequential UF by stirred cell Amicon 8400 (1, 3, 5,10 kDa) | -Antioxidant (inhibitor lipid peroxidation) | Peptides < 1 kDa Peptides < 5 kDa | Yield: 36.97% (<1 kDa from pepsin–pancreatin hydrolysate), 28.82% (1–3 kDa, Alcalase), 37.27% (1–3 kDa, pancreatin) | [36] |
| Soy protein | Flavourzyme | Sequential UF (30, 10, 1, 0.3 kDa) | -Lipolysis stimulator | Peptides ~1 kDa | Intracellular triglyceride decrease (2.73 to 2.30 mol mg−1 protein) by 400 ppm peptides | [37] |
| Vigna unguiculata (Cowpea) | Alcalase/Flavourzyme/pepsin–pancreatin | Sequential UF high performance UF cell (Model 2000, Millipore) (1, 3, 5, 10 kDa) | -ACE inhibitors -Antioxidants | Peptides < 1 kDa | ACE inhibitors: Alcalase: 24.3–123, Flavourzyme: 0.04–170.6, pancreatin: 44.7–112 μg mL−1 (IC50) Antioxidant: Alcalase: 303.2–1457; Flavourzyme: 357.4–10,211; Pancreatin hydrolysate: 267.1–2830.4 (Trolox equivalent) | [38] |
| Soybean meal fermented by Bacillus subtilis | Thermolysin | Sequential filtration (MF: fractions of > 10 kDa, UF: 1–10, NF: 0.5–1, RO: <0.5 kDa) | Antihypertensive | Peptides < 0.5 kDa, Fragments: QC, PANV, GPANV | Endothelium-independent vasorelaxation action (work similarly with ACE-I inhibitory peptides) | [39] |
| Soy protein | Flavourzyme® or chymotrypsin | Sequential UF (Regenerated cellulose (RC) 50, 10, and 3 kDa) | Antioxidants | Peptides < 3 kDa | Peptides from both hydrolysates decreased liposome oxidation by 83.2% and 84.5%, and showed DPPH activity of 21.3 and 24.4%, respectively | [40] |
| Phaseolus lunatus and Proteus vulgaris | Alcalase | Sequential UF high-performance UF cell (Model 2000, Millipore) (1, 3, 5, 10 kDa) | ACE inhibitors | Peptides < 1 kDa | -Activity determined by higher constituents of hydrophobic amino acids such as valine and proline -IC50 (μg mL−1): Phaseolus lunatus 30.3 and P. vulgaris 63 | [41] |
| Soybean protein | - | Sequential anion-exchange chromatography, UF, reversed-phase chromatography | Anticancer | Lunasin | -Yield: 442 mg kg−1 defatted flour (99% purity) -Histone-binding assays: biological activity is similar to those of the synthetic drugs | [42] |
| Soy protein | Neutrase | Sequential UF by spiral-wound (30, 10, 1 kDa) | Antiadipogenic | Peptides 1.3–2.2 kDa | Decrease of glycerol-3-phosphate dehydrogenase (GPDH) activity decreased (280 to 100 U mg−1) and relative lipid accumulation (RLA) in 3T3-L1 cells | [43] |
| Corn protein | Alcalase | Sequential UF by Prep/Scale Tangential Flow Filtratuin (TFF) system 230V (Millipore) (RC 5, 3, 1 kDa) | Alcohol metabolism stimulator | Peptides < 1 kDa | -Activating hepatic alcohol dehydrogenase (ADH) -Peptides < 5 kDa triggered maximum ability to decrease blood alcohol concentration (BAC) | [44] |
| Corn protein | Validase fungal protease (FP), alkaline protease (AP), and/or neutral protease (NP) | Sequential UF by Millipore 8050 ultra-filtration (RC 10, 3, 1 kDa) | Antioxidants | F1: >10 kDa F2: 3–10 kDa F3: 1–3 kDa F4: <1 kDa | -NP-F3 had the highest activity (ORAC and Trolox equivalents, lipid oxidation inhibition) -AP-F2 and AP-F3 had highest activity (DPPH) | [45] |
| Broken rice protein | Alkaline protease | UF-coupled hydrolysis with hollow fiber module (Polyvinylidene fluoride (PVDF) 6 kDa) | Antioxidant | Oligopeptides (1.2–0.2 kDa) | -Oligopeptides content increased (60%) compared to batch hydrolysis (40%). 75% DPPH activity for 3-h operation | [46] |
| Source | Hydrolysis Enzyme(s) | Protocols | Bioactivity | Target Compound(s) | Remarks | Ref |
|---|---|---|---|---|---|---|
| Catfish muscle protein | Thermolysin | Sequential UF 10, 5 (Vivaflow 200), 3 kDa (Vivaflow 50) | ACE inhibitors | Peptides: GPPP and IEKPP | -GPPP (IC50: 0.86 μM) -IEKPP (IC50: 1.2 μM) | [47] |
| Yellow croaker | Neutral protease | Sequential UF 10, 5 (Vivaflow 200), 3 kDa (Vivaflow 50) | Antioxidant | Peptides < 3 kDa: SRCHV and PEHW | IC50: 7.67 mg mL−1 (O2- scavenging) IC50: 7.68 mg mL−1 (DPPH) | [48] |
| Round cod muscle protein | Alcalase | Labscale TFF System, Millipore (10 and 5 kDa) | Antioxidant | Peptides < 5 kDa | Activity of sequence HDHPVC (0.7 kDa): IC50 0.38 mM (O2- scavenging), 0.03 mM (DPPH) and HEKVC (0.6 kDa): IC50 0.37 mM (O2- scavenging), 0.07 mM (DPPH) | [49] |
| Cuttlefish wastewater | Alcalase | Cascaded UF-DF Prep/Scale-TFF cartridges, Millipore (PES 100, 30, 10 kDa, RC 1 kDa) | -Antihypertensive -Antioxidant | Peptides < 10 kDa | Highest antihypertensive fractions from boiling water effluents by UF configurations: 100-30-10kDa (IC50: 100 μg mL−1). Highest antioxidant activities from (1) softening effluents by one UF filtration of 10 kDa PES (β-carotene assay: 9 μM BHT mg−1, DPPH assay: 167 μM BHT mg−1) and (2) from boiling water effluents by configuration 100-10 kDa (β-carotene assay: 18 μM BHT mg−1, DPPH assay: 89 μM BHT mg−1) | [50] |
| Yellowfin tuna’s viscera | Protamex | Sequential UF by Amicon-stirred UF cell (RC 30, 10, and 3 kDa) | -Antibacterial -Antioxidant | Peptides < 3 kDa | -Minimum inhibition: 0.5 mg mL−1, IC50: 1.8 mg mL−1 (DPPH); 1.4 mg mL−1 (ABTS). Toward Gram—(Escherichia coli, 99.7% inhibition; Pseudomonas aeruginosa, 99.76% inhibition). Toward Gram + (Listeria monocytogenes, 99.88% inhibition; Staphylococcus aureus, 99.76% inhibition) | [51] |
| Hoki frame | Pepsin | Sequential UF (5, 3, 1 kDa) | Antioxidant | Peptides 1–3 kDa | 85% DPPH, hydroxyl radical, and alkyl radical scavenging activity in linoleic emulsion | [52] |
| Argentine croaker | Flavourzyme/ α-chymotrypsin | UF cell (Advantec, UHP-76) (RC 0.5 and 1.0 kDa) | Antioxidant | Peptides higher than 1 kDa | Highest activity of lipid oxidation inhibition generated from Argentine croaker by Flavourzyme owing to greater proportion of hydrophobic sulfuric amino acids | [53] |
| Tuna dark muscle protein | Alcalase | Tangential UF-NF UF: tubular ceramic membrane 8 kDa NF: flat PES 1 kDa | Antioxidant | Peptides < 1 kDa Peptides 1–4 kDa | Peptides < 1 kDa: highest scavenging activity of free radicals (75%) and hydroxyl radicals (65%). Peptides 1–4 kDa: stronger iron chelating ability (40%) and highest superoxide radical (65%) | [54] |
| Source | Hydrolysis Enzyme(s) | Protocols | Bioactivity | Target Compound(s) | Remarks | Ref |
|---|---|---|---|---|---|---|
| Egg white protein (US-pretreated at 40 kHz, 15 min) | Alcalase | Sequential UF by Millipore UF stirred cell unit 8050 (RC 30, 10, and 1 kDa) | Antioxidant | Peptides 1–10 kDa | Activity determined by DPPH: 28.10% ± 1.38% and ABTS: 79.44% ± 2.31% | [55] |
| Dephosphorylated egg-yolk protein | Alcalase, protease N, trypsin VI | Sequential UF (RC 5 and 1 kDa) | Antioxidant | Peptides < 1 kDa (amino acids H, M, L, F) | A 3-fold activity increase from nonhydrolysed dephosphorylated egg-yolk protein, confirmed by ORAC | [56] |
| Dilapidated egg yolk protein concentrates | - | UF-DF: (Spiral-wound PES 10 and 30 kDa) | Antioxidant | Phosvitin | Protein increased 6.25-fold by 10 kDa membrane (5.92-fold by 30 kDa). 10 and 30 kDaMWCO yields were similar (84%) | [57] |
| Goat milk protein hydrolysates | Subtilisin and trypsin | (1) casein and whey separated using 0.14 μm ceramic membrane; (2) retentate containing casein is hydrolysed; (3) hydrolysates then filtered by 50 kDa ceramic membrane | ACE inhibitors | Peptides < 50 kDa | Permeate of 50 kDa UF showed 30% enhancement of ACE-inhibitory activities (IC50: 218.50 μg mL−1) | [58] |
2. Advancements in Ultrafiltration for Protein and Peptide Separation
2.1. Charged-Membrane
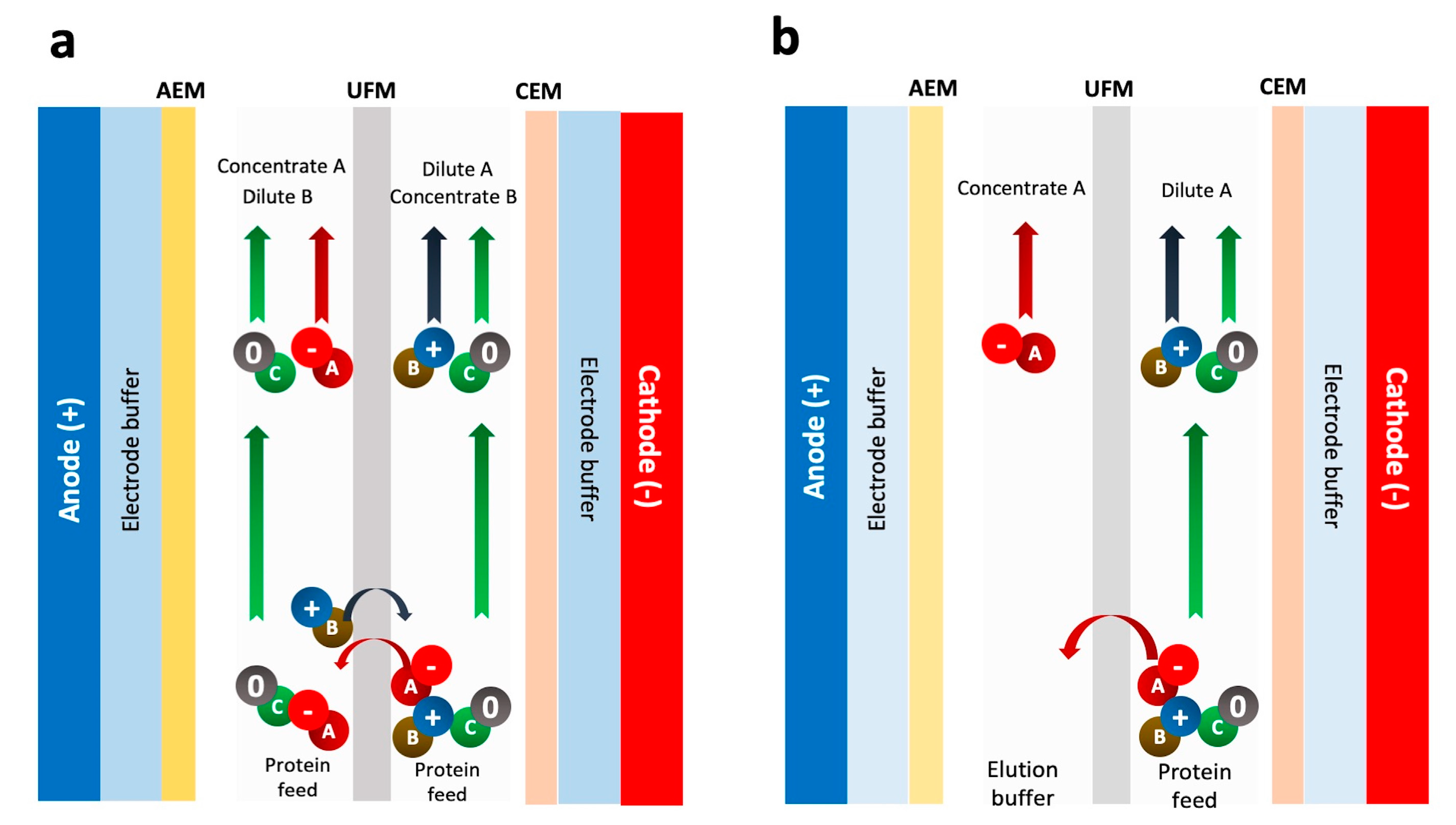
2.2. Electroultrafiltration (EUF) and Electrodialysis with UF Membrane (EDUF)
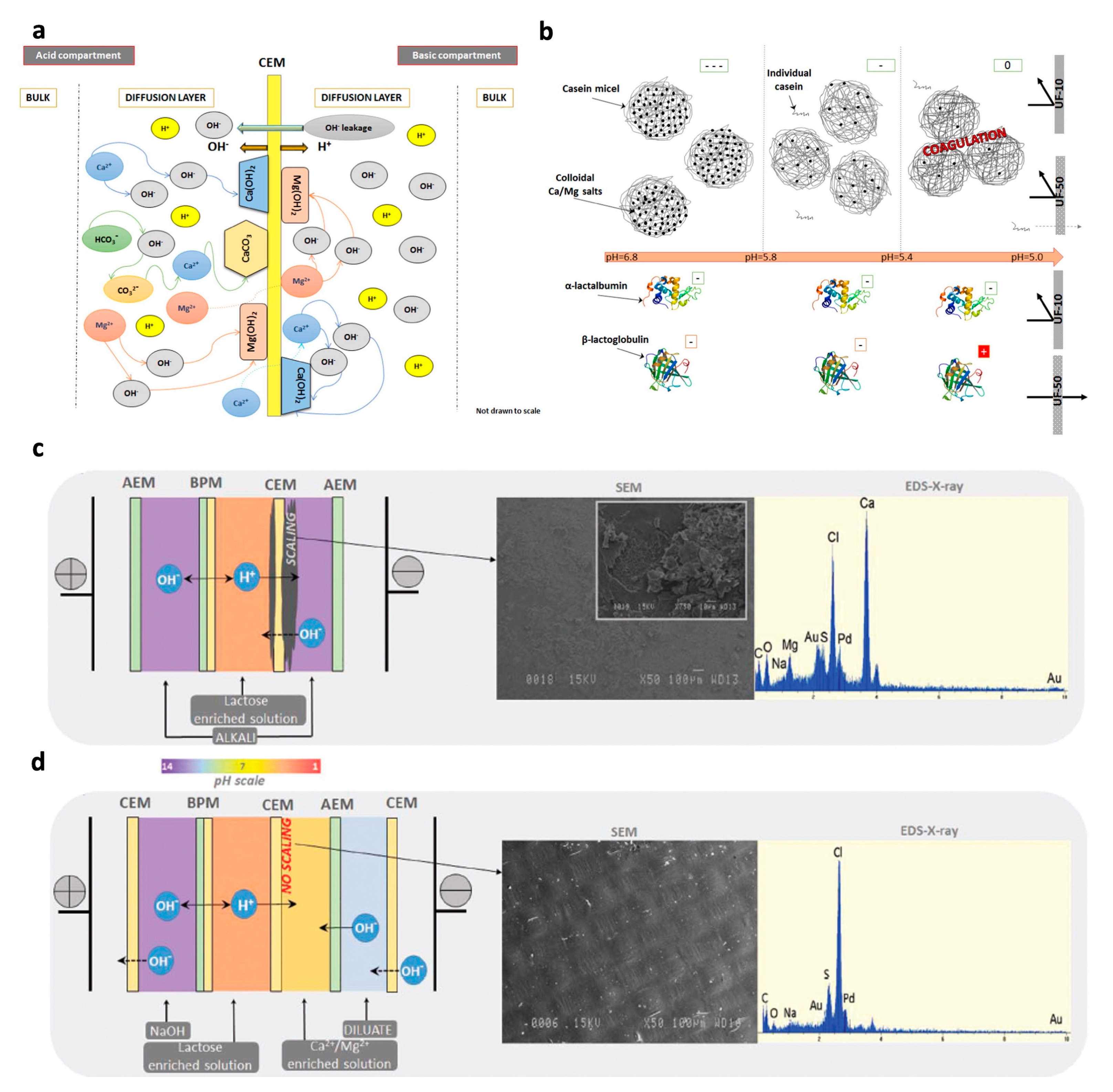
2.3. Combining Electrodialysis with Bipolar Membrane (EDBM) and UF
2.4. Ultrasonic-Assisted UF
2.5. Dynamic UF
2.6. High-Performance Tangential Flow Filtration (HPTFF)
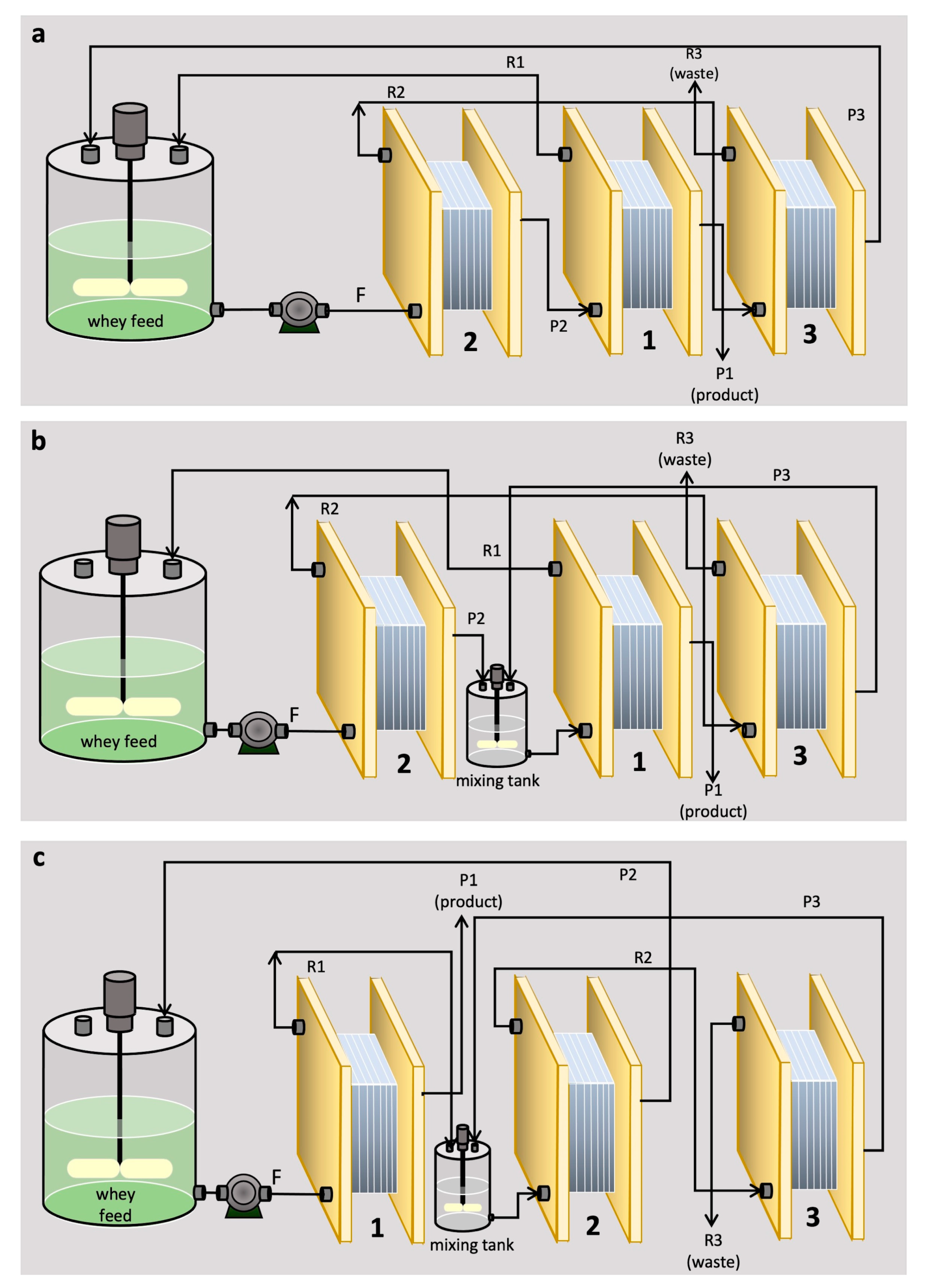
2.7. Integration of UF with Other Processes
3. Strategic Development of Ultrafiltration Membrane
3.1. Membrane Modules and Fiber Diameter
3.2. Membrane Materials and Molecular Weight Cutoff
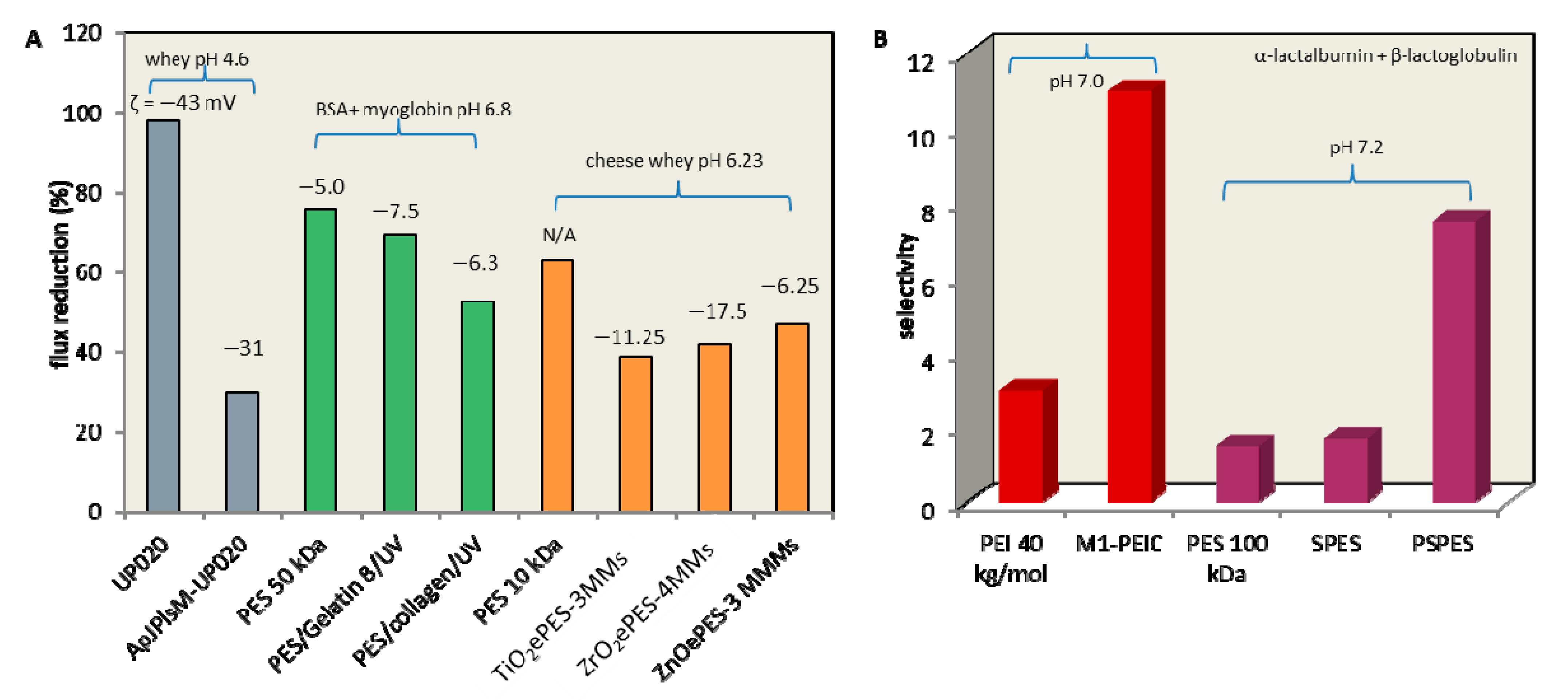
3.3. Surface Modifications
4. Ultrafiltration Performance and Selectivity Enhancements: Controls of Fouling and Concentration Polarization from Operating Conditions Perspectives
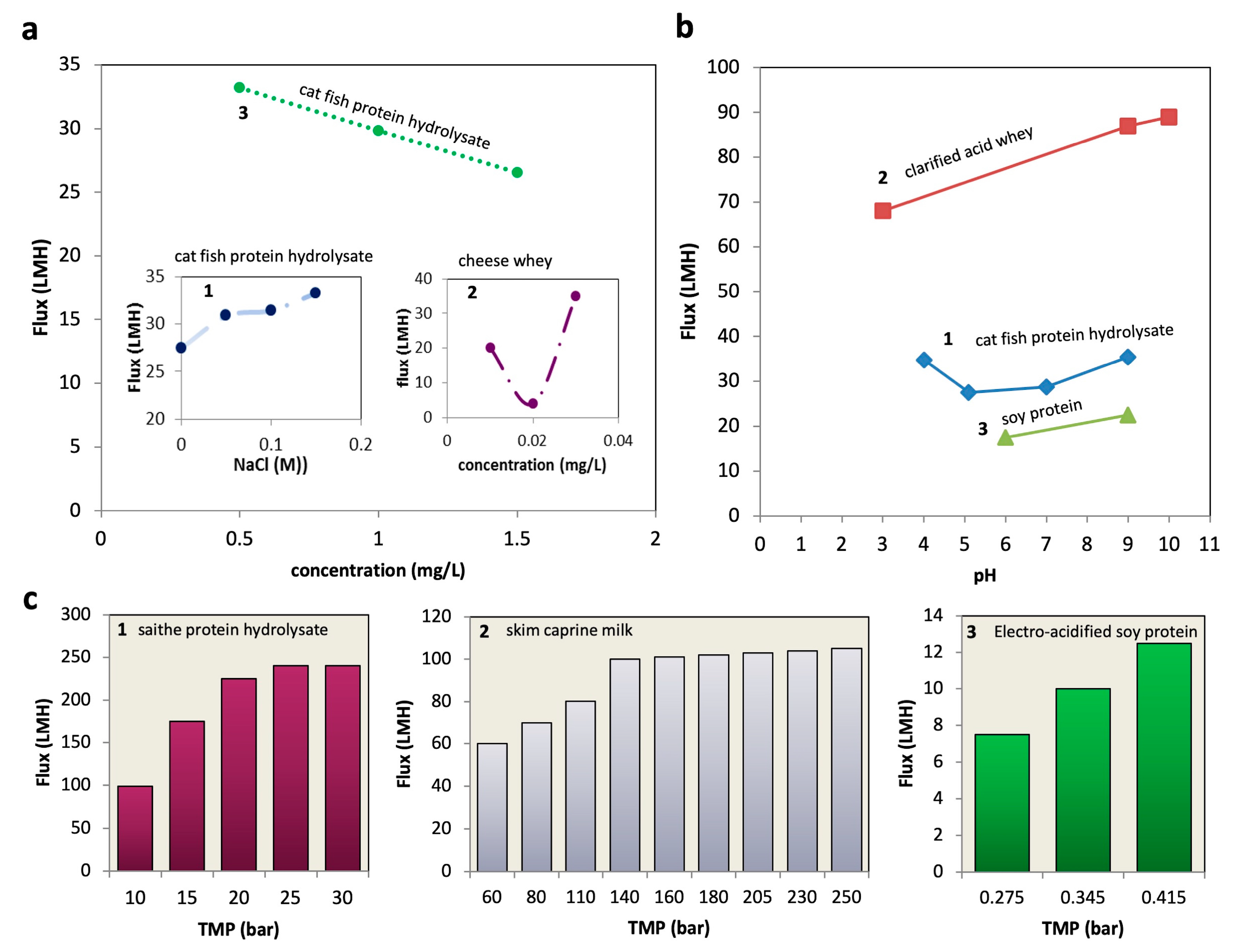
4.1. Protein Concentration
4.2. pH in Pretreatment
4.3. Ionic Strength
4.4. Operating Pressure
4.5. Temperature
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hayes, M. Food proteins and bioactive peptides: New and novel sources, characterisation strategies and applications. Foods 2018, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.J.; Brooks, M.S.-L. Food industry protein by-products and their applications. In Protein Byproducts; Dhillon, G.S., Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2016; pp. 120–132. [Google Scholar]
- Rabert, C.; Weinacker, D.; Pessoa, A.; Farias, J. Recombinants proteins for industrial uses: Utilization of Pichia pastoris expression system. Braz. J. Microbiol. 2013, 44, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides—Opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhao, G.; Ren, J.; Zhao, M.; Yang, B. Effect of denaturation during extraction on the conformational and functional properties of peanut protein isolate. Innov. Food Sci. Emerg. Technol. 2011, 12, 375–380. [Google Scholar] [CrossRef]
- Plaxton, W.C. Avoiding proteolysis during the extraction and purification of active plant enzymes. Plant. Cell Physiol. 2019, 60, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Ryan, B.J.; Henehan, G.T. Avoiding proteolysis during protein purification. In Protein Chromatography: Methods and Protocols; Walls, D., Loughran, S.T., Eds.; Springer: New York, NY, USA, 2017; pp. 53–69. [Google Scholar]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Ji, H.; Mathivanan, S.; Scott, A.M.; Simpson, R.J. Comparison of ultracentrifugation, density gradient separation, and immunoaffinity capture methods for isolating human colon cancer cell line LIM1863-derived exosomes. Methods 2012, 56, 293–304. [Google Scholar] [CrossRef]
- Coskun, O. Separation techniques: Chromatography. North. Clin. Istanb. 2016, 3, 156–160. [Google Scholar]
- Wenten, I.G.; Khoiruddin, K.; Reynard, R.; Lugito, G.; Julian, H. Advancement of forward osmosis (FO) membrane for fruit juice concentration. J. Food Eng. 2021, 290, 110216. [Google Scholar] [CrossRef]
- Wenten, I.G.; Friatnasary, D.L.; Khoiruddin, K.; Setiadi, T.; Boopathy, R. Extractive membrane bioreactor (EMBR): Recent advances and applications. Bioresour. Technol. 2020, 297, 122424. [Google Scholar] [CrossRef]
- Aryanti, P.T.P.; Subroto, E.; Mangindaan, D.; Widiasa, I.N.; Wenten, I.G. Semi-industrial high-temperature ceramic membrane clarification during starch hydrolysis. J. Food Eng. 2020, 274, 109844. [Google Scholar] [CrossRef]
- Julian, H.; Yaohanny, F.; Devina, A.; Purwadi, R.; Wenten, I.G. Apple juice concentration using submerged direct contact membrane distillation (SDCMD). J. Food Eng. 2020, 272, 109807. [Google Scholar] [CrossRef]
- Julian, H.; Rizqullah, H.; Siahaan, M.A.; Wenten, I.G. Cane sugar crystallization using submerged vacuum membrane distillation crystallization (SVMDC). J. Food Sci. Technol. 2020, in press. [Google Scholar] [CrossRef]
- Rathore, A.S.; Shirke, A. Recent developments in membrane-based separations in biotechnology processes: Review. Prep. Biochem. Biotechnol. 2011, 41, 398–421. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Tripathi, B.P.; Kumar, M.; Shahi, V.K. Membrane-based techniques for the separation and purification of proteins: An overview. Adv. Colloid Interface Sci. 2009, 145, 1–22. [Google Scholar] [CrossRef]
- Agüero, R.; Bringas, E.; San Román, M.F.; Ortiz, I.; Ibáñez, R. Membrane processes for whey proteins separation and purification. A review. Curr. Org. Chem. 2017, 21, 1740–1752. [Google Scholar] [CrossRef]
- Vedadghavami, A.; Minooei, F.; Hosseini, S.S. Practical techniques for improving the performance of polymeric membranes and processes for protein separation and purification. Iran Chem. Chem. Eng. 2018, 37, 1–23. [Google Scholar]
- Dhineshkumar, V.; Ramasamy, D. Review on membrane technology applications in food and dairy processing. J. Appl. Biotechnol. Bioeng. 2017, 3, 399–407. [Google Scholar]
- Mohammad, A.W.; Ng, C.Y.; Lim, Y.P.; Ng, G.H. Ultrafiltration in food processing industry: Review on application, membrane fouling, and fouling control. Food Bioprocess. Technol. 2012, 5, 1143–1156. [Google Scholar] [CrossRef]
- Chao, G.; Shuili, Y.; Yufei, S.; Zhengyang, G.; Wangzhen, Y.; Liumo, R. A review of ultrafiltration and forward osmosis: Application and modification. IOP Conf. Ser. Earth Environ. Sci. 2018, 128, 012150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Grimi, N.; Jaffrin, M.Y.; Ding, L.; Tang, B. A short review on the research progress in alfalfa leaf protein separation technology. J. Chem. Technol. Biotechnol. 2017, 92, 2894–2900. [Google Scholar] [CrossRef]
- Datta, D.; Bhattacharjee, C.; Datta, S. Whey protein fractionation using membrane filtration—A review. J. Inst. Eng. Chem. Eng. Div. 2008, 89, 45–50. [Google Scholar]
- Kumar, P.; Sharma, N.; Ranjan, R.; Kumar, S.; Bhat, Z.F.; Jeong, D.K. Perspective of membrane technology in dairy industry: A review. Asian Australas. J. Anim. Sci. 2013, 26, 1347–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, M.D.; Bórquez, R. Review of the treatment of seafood processing wastewaters and recovery of proteins therein by membrane separation processes—Prospects of the ultrafiltration of wastewaters from the fish meal industry. Desalination 2002, 142, 29–45. [Google Scholar] [CrossRef]
- Aluko, R.E. Technology for the production and utilization of food protein-derived anti-hypertensive peptides: A review. Recent Pat. Biotechnol. 2007, 1, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Polyakov, Y.S.; Zydney, A.L. Ultrafiltration membrane performance: Effects of pore blockage/constriction. J. Memb. Sci. 2013, 434, 106–120. [Google Scholar] [CrossRef]
- Marshall, A.D.; Munro, P.A.; Trägårdh, G. The effect of protein fouling in microfiltration and ultrafiltration on permeate flux, protein retention and selectivity: A literature review. Desalination 1993, 91, 65–108. [Google Scholar] [CrossRef]
- Fane, A.G.; Fell, C.J.D. A review of fouling and fouling control in ultrafiltration. Desalination 1987, 62, 117–136. [Google Scholar] [CrossRef]
- Kallioinen, M.; Mänttäri, M. Influence of ultrasonic treatment on various membrane materials: A review. Sep. Sci. Technol. 2011, 46, 1388–1395. [Google Scholar] [CrossRef]
- Dlask, O.; Václavíková, N. Electrodialysis with ultrafiltration membranes for peptide separation. Chem. Pap. 2018, 72, 261–271. [Google Scholar] [CrossRef]
- Jaffrin, M.Y. Dynamic shear-enhanced membrane filtration: A review of rotating disks, rotating membranes and vibrating systems. J. Memb. Sci. 2008, 324, 7–25. [Google Scholar] [CrossRef]
- Denis, C.; Massé, A.; Fleurence, J.; Jaouen, P. Concentration and pre-purification with ultrafiltration of a R-phycoerythrin solution extracted from macro-algae Grateloupia turuturu: Process definition and up-scaling. Sep. Purif. Technol. 2009, 69, 37–42. [Google Scholar] [CrossRef]
- Pan, S.; Wang, S.; Jing, L.; Yao, D. Purification and characterisation of a novel angiotensin-I converting enzyme (ACE)-inhibitory peptide derived from the enzymatic hydrolysate of Enteromorpha clathrata protein. Food Chem. 2016, 211, 423–430. [Google Scholar] [CrossRef]
- Olagunju, A.I.; Omoba, O.S.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Pigeon pea enzymatic protein hydrolysates and ultrafiltration peptide fractions as potential sources of antioxidant peptides: An in vitro study. LWT 2018, 97, 269–278. [Google Scholar] [CrossRef]
- Tsou, M.-J.; Lin, S.-B.; Chao, C.-H.; Chiang, W.-D. Enhancing the lipolysis-stimulating activity of soy protein using limited hydrolysis with Flavourzyme and ultrafiltration. Food Chem. 2012, 134, 1564–1570. [Google Scholar] [CrossRef]
- Campos, M.R.S.; Guerrero, L.A.C.; Ancona, D.A.B. Angiotensin-I converting enzyme inhibitory and antioxidant activities of peptide fractions extracted by ultrafiltration of cowpea Vigna unguiculata hydrolysates. J. Sci. Food Agric. 2010, 90, 2512–2518. [Google Scholar] [CrossRef]
- Wang, Z.; Cui, Y.; Liu, P.; Zhao, Y.; Wang, L.; Liu, Y.; Xie, J. Small peptides isolated from enzymatic hydrolyzate of fermented soybean meal promote endothelium-independent vasorelaxation and ACE inhibition. J. Agric. Food Chem. 2017, 65, 10844–10850. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, E.I.; Calderón de la Barca, A.M.; Sotelo-Mundo, R.R.; Arteaga-Mackinney, G.E.; Valenzuela-Melendez, M.; Peña-Ramos, E.A. Partial characterization of ultrafiltrated soy protein hydrolysates with antioxidant and free radical scavenging activities. J. Food Sci. 2013, 78, C1152–C1158. [Google Scholar] [CrossRef]
- Betancur-Ancona, D.; Dávila-Ortiz, G.; Chel-Guerrero, L.A.; Torruco-Uco, J.G. ACE-I inhibitory activity from phaseolus lunatus and Phaseolus vulgaris peptide fractions obtained by ultrafiltration. J. Med. Food 2015, 18, 1247–1254. [Google Scholar] [CrossRef]
- Seber, L.E.; Barnett, B.W.; McConnell, E.J.; Hume, S.D.; Cai, J.; Boles, K.; Davis, K.R. Scalable purification and characterization of the anticancer lunasin peptide from soybean. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Tsou, M.-J.; Lin, W.-T.; Lu, H.-C.; Tsui, Y.-L.; Chiang, W.-D. The effect of limited hydrolysis with Neutrase and ultrafiltration on the anti-adipogenic activity of soy protein. Process. Biochem. 2010, 45, 217–222. [Google Scholar] [CrossRef]
- Yu, G.-C.; Li, J.-T.; He, H.; Huang, W.-H.; Zhang, W.-J. Ultrafiltration preparation of potent bioactive corn peptide as alcohol metabolism stimulator in vivo and study on its mechanism of action. J. Food Biochem. 2013, 37, 161–167. [Google Scholar] [CrossRef]
- Zhou, K.; Sun, S.; Canning, C. Production and functional characterisation of antioxidative hydrolysates from corn protein via enzymatic hydrolysis and ultrafiltration. Food Chem. 2012, 135, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Lv, Z.; Liu, Y.; Lu, F. Preparing oligopeptides from broken rice protein by ultrafiltration-coupled enzymatic hydrolysis. Eur. Food Res. Technol. 2013, 236, 419–424. [Google Scholar] [CrossRef]
- Ghassem, M.; Arihara, K.; Babji, A.S. Isolation, purification and characterisation of angiotensin I-converting enzyme-inhibitory peptides derived from catfish (Clarias batrachus) muscle protein thermolysin hydrolysates. Int. J. Food Sci. Technol. 2012, 47, 2444–2451. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, C.; Chen, Y.; Zheng, B. Purification and characterization of antioxidant peptides of Pseudosciaena crocea protein hydrolysates. Molecules 2017, 22, 57. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Tong, T.; Sun, J.; Xu, Y.; Zhao, Z.; Liao, D. Purification and characterization of antioxidative peptides from round scad (Decapterus maruadsi) muscle protein hydrolysate. Food Chem. 2014, 154, 158–163. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; González, M.P.; Murado, M.A. Production of antihypertensive and antioxidant activities by enzymatic hydrolysis of protein concentrates recovered by ultrafiltration from cuttlefish processing wastewaters. Biochem. Eng. J. 2013, 76, 43–54. [Google Scholar] [CrossRef]
- Pezeshk, S.; Ojagh, S.M.; Rezaei, M.; Shabanpour, B. Fractionation of protein hydrolysates of fish waste using membrane ultrafiltration: Investigation of antibacterial and antioxidant activities. Probiotics Antimicrob. Proteins 2019, 11, 1015–1022. [Google Scholar] [CrossRef]
- Je, J.-Y.; Kim, S.-Y.; Kim, S.-K. Preparation and antioxidative activity of hoki frame protein hydrolysate using ultrafiltration membranes. Eur. Food Res. Technol. 2005, 221, 157–162. [Google Scholar] [CrossRef]
- Centenaro, G.S.; Salas-Mellado, M.; Pires, C.; Batista, I.; Nunes, M.L.; Prentice, C. Fractionation of protein hydrolysates of fish and chicken using membrane ultrafiltration: Investigation of antioxidant activity. Appl. Biochem. Biotechnol. 2014, 172, 2877–2893. [Google Scholar] [CrossRef] [PubMed]
- Saidi, S.; Deratani, A.; Belleville, M.-P.; Ben Amar, R. Antioxidant properties of peptide fractions from tuna dark muscle protein by-product hydrolysate produced by membrane fractionation process. Food Res. Int. 2014, 65, 329–336. [Google Scholar] [CrossRef]
- Jovanović, J.R.; Stefanović, A.B.; Žuža, M.G.; Jakovetić, S.M.; Šekuljica, N.Ž.; Bugarski, B.M.; Knežević-Jugović, Z.D. Improvement of antioxidant properties of egg white protein enzymatic hydrolysates by membrane ultrafiltration. Hem. Ind. 2016, 70, 419–428. [Google Scholar] [CrossRef]
- Ting, B.P.C.P.; Mine, Y.; Juneja, L.R.; Okubo, T.; Gauthier, S.F.; Pouliot, Y. Comparative composition and antioxidant activity of peptide fractions obtained by ultrafiltration of egg yolk protein enzymatic hydrolysates. Membranes 2011, 1, 149–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chay Pak Ting, B.P.; Pouliot, Y.; Juneja, L.R.; Okubo, T.; Gauthier, S.F.; Mine, Y. On the use of ultrafiltration for the concentration and desalting of phosvitin from egg yolk protein concentrate. Int. J. Food Sci. Technol. 2010, 45, 1633–1640. [Google Scholar] [CrossRef]
- Espejo-Carpio, F.J.; Pérez-Gálvez, R.; Del Carmen Almécija, M.; Guadix, A.; Guadix, E.M. Production of goat milk protein hydrolysate enriched in ACE-inhibitory peptides by ultrafiltration. J. Dairy Res. 2014, 81, 385–393. [Google Scholar] [CrossRef]
- Wenten, I.G. Mechanisms and control of fouling in crossflow microfiltration. Filtr. Sep. 1995, 32, 252–253. [Google Scholar] [CrossRef]
- Widodo, S.; Ariono, D.; Khoiruddin, K.; Hakim, A.N.; Wenten, I.G. Recent advances in waste lube oils processing technologies. Environ. Prog. Sustain. Energy 2018. [Google Scholar] [CrossRef]
- Zhang, W.; Luo, J.; Ding, L.; Jaffrin, M.Y. A review on flux decline control strategies in pressure-driven membrane processes. Ind. Eng. Chem. Res. 2015, 54, 2843–2861. [Google Scholar] [CrossRef]
- Arunkumar, A.; Molitor, M.S.; Etzel, M.R. Comparison of flat-sheet and spiral-wound negatively-charged wide-pore ultrafiltration membranes for whey protein concentration. Int. Dairy J. 2016, 56, 129–133. [Google Scholar] [CrossRef]
- Mehta, A.; Zydney, A.L. Permeability and selectivity analysis for ultrafiltration membranes. J. Memb. Sci. 2005, 249, 245–249. [Google Scholar] [CrossRef]
- Metsämuuronen, S.; Nyström, M. Enrichment of α-lactalbumin from diluted whey with polymeric ultrafiltration membranes. J. Memb. Sci. 2009, 337, 248–256. [Google Scholar] [CrossRef]
- Yee, K.W.K.; Wiley, D.E.; Bao, J. Whey protein concentrate production by continuous ultrafiltration: Operability under constant operating conditions. J. Memb. Sci. 2007, 290, 125–137. [Google Scholar] [CrossRef]
- Etzel, M.R.; Arunkumar, A. Dairy protein fractionation and concentration using charged ultrafiltration membranes. In Membrane Processes for Dairy Ingredient Separation; Hu, K., Dickson, J.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 86–111. [Google Scholar]
- Mehta, A.; Zydney, A.L. Effect of membrane charge on flow and protein transport during ultrafiltration. Biotechnol. Prog. 2006, 22, 484–492. [Google Scholar] [CrossRef]
- Van Reis, R.D. Charged Filtration Membranes and Uses Therefor. U.S. Patent No. 7,001,550 B2, 21 February 2006. [Google Scholar]
- Bhushan, S.; Etzel, M.R. Charged ultrafiltration membranes increase the selectivity of whey protein separations. J. Food Sci. 2009, 74, E131–E139. [Google Scholar] [CrossRef]
- Arunkumar, A.; Etzel, M.R. Fractionation of α-lactalbumin from β-lactoglobulin using positively charged tangential flow ultrafiltration membranes. Sep. Purif. Technol. 2013, 105, 121–128. [Google Scholar] [CrossRef]
- Arunkumar, A.; Etzel, M.R. Fractionation of α-lactalbumin and β-lactoglobulin from bovine milk serum using staged, positively charged, tangential flow ultrafiltration membranes. J. Memb. Sci. 2014, 454, 488–495. [Google Scholar] [CrossRef]
- Rabiller-Baudry, M.; Chaufer, B.; Lucas, D.; Michel, F. Ultrafiltration of mixed protein solutions of lysozyme and lactoferrin: Role of modified inorganic membranes and ionic strength on the selectivity. J. Memb. Sci. 2001, 184, 137–148. [Google Scholar] [CrossRef]
- Song, W.; Su, Y.; Chen, X.; Ding, L.; Wan, Y. Rapid concentration of protein solution by a crossflow electro- ultrafiltration process. Sep. Purif. Technol. 2010, 73, 310–318. [Google Scholar] [CrossRef]
- Sarkar, B.; DasGupta, S.; De, S. Electric field enhanced fractionation of protein mixture using ultrafiltration. J. Memb. Sci. 2009, 341, 11–20. [Google Scholar] [CrossRef]
- Oussedik, S.; Belhocine, D.; Grib, H.; Lounici, H.; Piron, D.L.; Mameri, N. Enhanced ultrafiltration of bovine serum albumin with pulsed electric field and fluidized activated alumina. Desalination 2000, 127, 59–68. [Google Scholar] [CrossRef]
- Chen, G.; Song, W.; Qi, B.; Li, J.; Ghosh, R.; Wan, Y. Separation of protein mixtures by an integrated electro-ultrafiltration-electrodialysis process. Sep. Purif. Technol. 2015, 147, 32–43. [Google Scholar] [CrossRef]
- Galier, S.; Balmann, H.R. The electrophoretic membrane contactor: A mass-transfer-based methodology applied to the separation of whey proteins. Sep. Purif. Technol. 2011, 77, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Galier, S.; Roux-de Balmann, H. Study of biomolecules separation in an electrophoretic membrane contactor. J. Memb. Sci. 2004, 241, 79–87. [Google Scholar] [CrossRef]
- Ketnawa, S.; Suwal, S.; Huang, J.-Y.; Liceaga, A.M. Selective separation and characterisation of dual ACE and DPP-IV inhibitory peptides from rainbow trout (Oncorhynchus mykiss) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 1062–1073. [Google Scholar] [CrossRef]
- Mondor, M.; Ippersiel, D.; Lamarche, F.; Boye, J.I. Production of soy protein concentrates using a combination of electroacidification and ultrafiltration. J. Agric. Food Chem. 2004, 52, 6991–6996. [Google Scholar] [CrossRef]
- Mikhaylin, S.; Patouillard, L.; Margni, M.; Bazinet, L. Milk protein production by a more environmentally sustainable process: Bipolar membrane electrodialysis coupled with ultrafiltration. Green Chem. 2018, 20, 449–456. [Google Scholar] [CrossRef]
- Luján-Facundo, M.-J.; Mendoza-Roca, J.-A.; Cuartas-Uribe, B.; Álvarez-Blanco, S. Membrane fouling in whey processing and subsequent cleaning with ultrasounds for a more sustainable process. J. Clean. Prod. 2017, 143, 804–813. [Google Scholar] [CrossRef]
- Sriniworn, P.; Youravong, W.; Khongnakorn, W. Recovery of protein from mung bean starch processing wastewater by rotating ultrafiltration. J. Eng. Sci. Technol. 2016, 11, 947–961. [Google Scholar]
- Akoum, O.; Jaffrin, M.Y.; Ding, L.-H. Concentration of total milk proteins by high shear ultrafiltration in a vibrating membrane module. J. Memb. Sci. 2005, 247, 211–220. [Google Scholar] [CrossRef]
- Lu, J.; Wan, Y.; Cui, Z. Fractionation of Lysozyme and chicken egg albumin using ultrafiltration with 30-kDa commercial membranes. Ind. Eng. Chem. Res. 2005, 44, 7610–7616. [Google Scholar] [CrossRef]
- Wan, Y.; Vasan, S.; Ghosh, R.; Hale, G.; Cui, Z. Separation of monoclonal antibody alemtuzumab monomer and dimers using ultrafiltration. Biotechnol. Bioeng. 2005, 90, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Enevoldsen, A.D.; Hansen, E.B.; Jonsson, G. Electro-ultrafiltration of industrial enzyme solutions. J. Memb. Sci. 2007, 299, 28–37. [Google Scholar] [CrossRef]
- Liu, C.; Monson, C.F.; Yang, T.; Pace, H.; Cremer, P.S. Protein separation by electrophoretic-electroosmotic focusing on supported lipid bilayers. Anal. Chem. 2011, 83, 7876–7880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barba, F.J.; Parniakov, O.; Pereira, S.A.; Wiktor, A.; Grimi, N.; Boussetta, N.; Saraiva, J.A.; Raso, J.; Martin-Belloso, O.; Witrowa-Rajchert, D.; et al. Current applications and new opportunities for the use of pulsed electric fields in food science and industry. Food Res. Int. 2015, 77, 773–798. [Google Scholar] [CrossRef]
- Catzel, D.; Lalevski, H.; Marquis, C.P.; Gray, P.P.; Van Dyk, D.; Mahler, S.M. Purification of recombinant human growth hormone from CHO cell culture supernatant by Gradiflow preparative electrophoresis technology. Protein Expr. Purif. 2003, 32, 126–134. [Google Scholar] [CrossRef]
- Käppler, T.; Posten, C. Fractionation of proteins with two-sided electro-ultrafiltration. J. Biotechnol. 2007, 128, 895–907. [Google Scholar] [CrossRef]
- Rothemund, D.L.; Thomas, T.M.; Rylatt, D.B. Purification of the basic protein avidin using Gradiflow technology. Protein Expr. Purif. 2002, 26, 149–152. [Google Scholar] [CrossRef]
- Robinson, C.W.; Siegel, M.; Condemine, A.; Fee, C.; Fahidy, T.Z.; Glick, B. Pulsed electric field cross-flow ultrafiltration of Bovine Serum Albumin. J. Memb. Sci. 1993, 80, 209–220. [Google Scholar] [CrossRef]
- Wang, K.; Wang, J.; Zhao, P.; Lin, S.; Liu, B.; Liu, J.; Jones, G.; Huang, H.-C. Optimized PEF treatment for antioxidant polypeptides with MW 10–30kDa and preliminary analysis of structure change. Int. J. Biol. Macromol. 2012, 51, 819–825. [Google Scholar] [CrossRef]
- Wenten, I.G.; Khoiruddin, K.; Alkhadra, M.A.; Tian, H.; Bazant, M.Z. Novel ionic separation mechanisms in electrically driven membrane processes. Adv. Colloid Interface Sci. 2020, 284, 102269. [Google Scholar] [CrossRef] [PubMed]
- Hakim, A.N.; Khoiruddin, K.; Ariono, D.; Wenten, I.G. Ionic separation in electrodeionization system: Mass transfer mechanism and factor affecting separation performance. Sep. Purif. Rev. 2020, 49, 294–316. [Google Scholar] [CrossRef]
- Handojo, L.; Wardani, A.K.; Regina, D.; Bella, C.; Kresnowati, M.T.A.P.; Wenten, I.G. Electro-membrane processes for organic acid recovery. RSC Adv. 2019, 9, 7854–7869. [Google Scholar] [CrossRef] [Green Version]
- Durand, R.; Fraboulet, E.; Marette, A.; Bazinet, L. Simultaneous double cationic and anionic molecule separation from herring milt hydrolysate and impact on resulting fraction bioactivities. Sep. Purif. Technol. 2019, 210, 431–441. [Google Scholar] [CrossRef]
- Langevin, M.-E.; Roblet, C.; Moresoli, C.; Ramassamy, C.; Bazinet, L. Comparative application of pressure- and electrically-driven membrane processes for isolation of bioactive peptides from soy protein hydrolysate. J. Memb. Sci. 2012, 403–404, 15–24. [Google Scholar] [CrossRef]
- Cecile Urbain Marie, G.; Perreault, V.; Henaux, L.; Carnovale, V.; Aluko, R.E.; Marette, A.; Doyen, A.; Bazinet, L. Impact of a high hydrostatic pressure pretreatment on the separation of bioactive peptides from flaxseed protein hydrolysates by electrodialysis with ultrafiltration membranes. Sep. Purif. Technol. 2019, 211, 242–251. [Google Scholar] [CrossRef]
- Kattan Readi, O.M.; Gironès, M.; Nijmeijer, K. Separation of complex mixtures of amino acids for biorefinery applications using electrodialysis. J. Memb. Sci. 2013, 429, 338–348. [Google Scholar] [CrossRef]
- Suwal, S.; Roblet, C.; Amiot, J.; Bazinet, L. Presence of free amino acids in protein hydrolysate during electroseparation of peptides: Impact on system efficiency and membrane physicochemical properties. Sep. Purif. Technol. 2015, 147, 227–236. [Google Scholar] [CrossRef]
- Zabolotskiy, V.I.; But, A.Y.; Vasil’eva, V.I.; Akberova, E.M.; Melnikov, S.S. Ion transport and electrochemical stability of strongly basic anion-exchange membranes under high current electrodialysis conditions. J. Memb. Sci. 2017, 526, 60–72. [Google Scholar] [CrossRef]
- Nikonenko, V.V.; Vasil’eva, V.I.; Akberova, E.M.; Uzdenova, A.M.; Urtenov, M.K.; Kovalenko, A.V.; Pismenskaya, N.P.; Mareev, S.A.; Pourcelly, G. Competition between diffusion and electroconvection at an ion-selective surface in intensive current regimes. Adv. Colloid Interface Sci. 2016, 235, 233–246. [Google Scholar] [CrossRef]
- Doyen, A.; Roblet, C.; L’Archevêque-Gaudet, A.; Bazinet, L. Mathematical sigmoid-model approach for the determination of limiting and over-limiting current density values. J. Memb. Sci. 2014, 452, 453–459. [Google Scholar] [CrossRef]
- Strathmann, H. Electrodialysis, a mature technology with a multitude of new applications. Desalination 2010, 264, 268–288. [Google Scholar] [CrossRef]
- Mikhaylin, S.; Bazinet, L. Electrodialysis in food processing. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–8. [Google Scholar]
- Balster, J.; Pünt, I.; Stamatialis, D.F.; Lammers, H.; Verver, A.B.; Wessling, M. Electrochemical acidification of milk by whey desalination. J. Memb. Sci. 2007, 303, 213–220. [Google Scholar] [CrossRef]
- Mier, M.P.; Ibañez, R.; Ortiz, I. Influence of process variables on the production of bovine milk casein by electrodialysis with bipolar membranes. Biochem. Eng. J. 2008, 40, 304–311. [Google Scholar] [CrossRef]
- Skorepova, J.; Moresoli, C. Carbohydrate and mineral removal during the production of low-phytate soy protein isolate by combined electroacidification and high shear tangential flow ultrafiltration. J. Agric. Food Chem. 2007, 55, 5645–5652. [Google Scholar] [CrossRef]
- Mikhaylin, S.; Nikonenko, V.; Pourcelly, G.; Bazinet, L. Hybrid bipolar membrane electrodialysis/ultrafiltration technology assisted by a pulsed electric field for casein production. Green Chem. 2015, 18, 307–314. [Google Scholar] [CrossRef]
- Ali, F.; Ippersiel, D.; Lamarche, F.; Mondor, M. Characterization of low-phytate soy protein isolates produced by membrane technologies. Innov. Food Sci. Emerg. Technol. 2010, 11, 162–168. [Google Scholar] [CrossRef]
- Mondor, M.; Ali, F.; Ippersiel, D.; Lamarche, F. Impact of ultrafiltration/diafiltration sequence on the production of soy protein isolate by membrane technologies. Innov. Food Sci. Emerg. Technol. 2010, 11, 491–497. [Google Scholar] [CrossRef]
- Bazinet, L.; Montpetit, D.; Ippersiel, D.; Amiot, J.; Lamarche, F. Identification of Skim Milk Electroacidification Fouling: A Microscopic Approach. J. Colloid Interface Sci. 2001, 237, 62–69. [Google Scholar] [CrossRef]
- Bazinet, L.; Ippersiel, D.; Gendron, C.; Beaudry, J.; Mahdavi, B.; Amiot, J.; Lamarche, F. Cationic balance in skim milk during bipolar membrane electroacidification. J. Memb. Sci. 2000, 173, 201–209. [Google Scholar] [CrossRef]
- Cifuentes-Araya, N.; Pourcelly, G.; Bazinet, L. Impact of pulsed electric field on electrodialysis process performance and membrane fouling during consecutive demineralization of a model salt solution containing a high magnesium/calcium ratio. J. Colloid Interface Sci. 2011, 361, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylin, S.; Sion, A.-V. Improvement of a sustainable hybrid technology for caseins isoelectric precipitation (electrodialysis with bipolar membrane/ultrafiltration) by mitigation of scaling on cation-exchange membrane. Innov. Food Sci. Emerg. Technol. 2016, 33, 571–579. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, M.; Fang, Z.-X.; Liu, Y. Effects of ultrasound and microwave pretreatments on the ultrafiltration desalination of salted duck egg white protein. Food Bioprod. Process. 2015, 96, 306–313. [Google Scholar] [CrossRef]
- Jiang, M.; Yan, H.; He, R.; Ma, Y. Purification and a molecular docking study of α-glucosidase-inhibitory peptides from a soybean protein hydrolysate with ultrasonic pretreatment. Eur. Food Res. Technol. 2018, 244, 1995–2005. [Google Scholar] [CrossRef]
- Khaire, R.A.; Gogate, P.R. Intensified recovery of lactose from whey using thermal, ultrasonic and thermosonication pretreatments. J. Food Eng. 2018, 237, 240–248. [Google Scholar] [CrossRef]
- Uluko, H.; Zhang, S.; Liu, L.; Tsakama, M.; Lu, J.; Lv, J. Effects of thermal, microwave, and ultrasound pretreatments on antioxidative capacity of enzymatic milk protein concentrate hydrolysates. J. Funct. Foods 2015, 18, 1138–1146. [Google Scholar] [CrossRef]
- Nemitz, M.C.; Moraes, R.C.; Koester, L.S.; Bassani, V.L.; von Poser, G.L.; Teixeira, H.F. Bioactive soy isoflavones: Extraction and purification procedures, potential dermal use and nanotechnology-based delivery systems. Phytochem. Rev. 2015, 14, 849–869. [Google Scholar] [CrossRef]
- Zhu, Z.; Jiang, T.; He, J.; Barba, F.J.; Cravotto, G.; Koubaa, M. Ultrasound-assisted extraction, centrifugation and ultrafiltration: Multistage process for polyphenol recovery from purple sweet potatoes. Molecules 2016, 21, 1584. [Google Scholar] [CrossRef]
- Yu, W.; Graham, N.; Liu, T. Effect of intermittent ultrasound on controlling membrane fouling with coagulation pre-treatment: Significance of the nature of adsorbed organic matter. J. Memb. Sci. 2017, 535, 168–177. [Google Scholar] [CrossRef]
- Luján-Facundo, M.J.; Mendoza-Roca, J.A.; Cuartas-Uribe, B.; Álvarez-Blanco, S. Cleaning efficiency enhancement by ultrasounds for membranes used in dairy industries. Ultrason. Sonochem. 2016, 33, 18–25. [Google Scholar] [CrossRef]
- Rajha, H.N.; Boussetta, N.; Louka, N.; Maroun, R.G.; Vorobiev, E. Effect of alternative physical pretreatments (pulsed electric field, high voltage electrical discharges and ultrasound) on the dead-end ultrafiltration of vine-shoot extracts. Sep. Purif. Technol. 2015, 146, 243–251. [Google Scholar] [CrossRef]
- Ahmad, A.L.; Che Lah, N.F.; Ismail, S.; Ooi, B.S. Membrane antifouling methods and alternatives: Ultrasound approach. Sep. Purif. Rev. 2012, 41, 318–346. [Google Scholar] [CrossRef]
- Ashokkumar, M.; Bhaskaracharya, R.; Kentish, S.; Lee, J.; Palmer, M.; Zisu, B. The ultrasonic processing of dairy products—An overview. Dairy Sci. Technol. 2010, 90, 147–168. [Google Scholar] [CrossRef] [Green Version]
- Gajendragadkar, C.N.; Gogate, P.R. Intensified recovery of valuable products from whey by use of ultrasound in processing steps—A review. Ultrason. Sonochem. 2016, 32, 102–118. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Avila, S.R.; Prabowo, F.; Kumar, A.; Ohl, C.-D. Improved ultrasonic cleaning of membranes with tandem frequency excitation. J. Memb. Sci. 2012, 415–416, 776–783. [Google Scholar] [CrossRef]
- Villamiel, M.; de Jong, P. Influence of high-intensity ultrasound and heat treatment in continuous flow on fat, proteins, and native enzymes of milk. J. Agric. Food Chem. 2000, 48, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Lamminen, M.O.; Walker, H.W.; Weavers, L.K. Mechanisms and factors influencing the ultrasonic cleaning of particle-fouled ceramic membranes. J. Memb. Sci. 2004, 237, 213–223. [Google Scholar] [CrossRef]
- Shanmugam, A.; Chandrapala, J.; Ashokkumar, M. The effect of ultrasound on the physical and functional properties of skim milk. Innov. Food Sci. Emerg. Technol. 2012, 16, 251–258. [Google Scholar] [CrossRef]
- Jin, Y.; Hengl, N.; Baup, S.; Pignon, F.; Gondrexon, N.; Sztucki, M.; Gésan-Guiziou, G.; Magnin, A.; Abyan, M.; Karrouch, M.; et al. Effects of ultrasound on cross-flow ultrafiltration of skim milk: Characterization from macro-scale to nano-scale. J. Memb. Sci. 2014, 470, 205–218. [Google Scholar] [CrossRef]
- Popović, S.; Djurić, M.; Milanović, S.; Tekić, M.N.; Lukić, N. Application of an ultrasound field in chemical cleaning of ceramic tubular membrane fouled with whey proteins. J. Food Eng. 2010, 101, 296–302. [Google Scholar] [CrossRef]
- Qu, T.; Pan, K.; Li, L.; Liang, B.; Wang, L.; Cao, B. Influence of ultrasonication conditions on the structure and performance of poly(vinylidene fluoride) membranes prepared by the phase inversion method. Ind. Eng. Chem. Res. 2014, 53, 8228–8234. [Google Scholar] [CrossRef]
- Kertész, S.; Bor, P.; Hodúr, C.; Csanádi, J.; Veréb, G.; Kovács, I.; Keszthelyi-Szabó, G.; László, Z. Effects of shear rate on membrane filtration. Desalin. Water Treat. 2017, 69, 43–49. [Google Scholar] [CrossRef]
- Serra, C.; Wiesner, M. A comparison of rotating and stationary membrane disk filters using computational fluid dynamics. J. Memb. Sci. 2000, 165, 19–29. [Google Scholar] [CrossRef]
- Engler, J.; Wiesner, M.R. Particle fouling of a rotating membrane disk. Water Res. 2000, 34, 557–565. [Google Scholar] [CrossRef]
- Ding, L.; Al-Akoum, O.; Abraham, A.; Jaffrin, M.Y. Milk protein concentration by ultrafiltration with rotating disk modules. Desalination 2002, 144, 307–311. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, W.; Ould-Dris, A.; Jaffrin, M.Y.; Tang, B. Concentration of milk proteins for producing cheese using a shear-enhanced ultrafiltration technique. Ind. Eng. Chem. Res. 2016, 55, 11130–11138. [Google Scholar] [CrossRef]
- Luo, J.; Ding, L.; Wan, Y.; Jaffrin, M.Y. Flux decline control in nanofiltration of detergent wastewater by a shear-enhanced filtration system. Chem. Eng. J. 2012, 181–182, 397–406. [Google Scholar] [CrossRef]
- Espina, V.; Jaffrin, M.Y.; Ding, L.; Cancino, B. Fractionation of pasteurized skim milk proteins by dynamic filtration. Food Res. Int. 2010, 43, 1335–1346. [Google Scholar] [CrossRef]
- Das, R.; Bhattacherjee, C.; Ghosh, S. Studies on membrane processing of sesame protein isolate and sesame protein hydrolysate using rotating disk module. Sep. Sci. Technol. 2009, 44, 131–150. [Google Scholar] [CrossRef]
- Zhang, W.; Ding, L.; Grimi, N.; Jaffrin, M.Y.; Tang, B. Application of UF-RDM (Ultafiltration Rotating Disk Membrane) module for separation and concentration of leaf protein from alfalfa juice: Optimization of operation conditions. Sep. Purif. Technol. 2017, 175, 365–375. [Google Scholar] [CrossRef]
- Zhang, W.; Ding, L.; Grimi, N.; Jaffrin, M.Y.; Tang, B. A rotating disk ultrafiltration process for recycling alfalfa wastewater. Sep. Purif. Technol. 2017, 188, 476–484. [Google Scholar] [CrossRef]
- Sarkar, P.; Ghosh, S.; Dutta, S.; Sen, D.; Bhattacharjee, C. Effect of different operating parameters on the recovery of proteins from casein whey using a rotating disc membrane ultrafiltration cell. Desalination 2009, 249, 5–11. [Google Scholar] [CrossRef]
- Al-Akoum, O.; Ding, L.; Chotard-Ghodsnia, R.; Jaffrin, M.Y.; Gésan-Guiziou, G. Casein micelles separation from skimmed milk using a VSEP dynamic filtration module. Desalination 2002, 144, 325–330. [Google Scholar] [CrossRef]
- Akoum, O.; Chotard-Ghodsnia, R.; Ding, L.H.; Jaffrin, M.Y. Ultrafiltration of low-heat and UHT skim milks with a shear-enhanced vibrating filtration system. Sep. Sci. Technol. 2003, 38, 571–589. [Google Scholar] [CrossRef]
- Hodúr, C.; Kertész, S.; Csanádi, J.; Szabó, G.; László, Z. Investigation of vibratory shear-enhanced processing system. Prog. Agric. Eng. Sci. 2009, 5, 97–110. [Google Scholar] [CrossRef]
- Hodúr, C.; Kertèsz, S.; Csanádi, J.; Szabó, G. Comparison of 3DTA and VSEP systems during the ultrafiltration of sweet whey. Desalin. Water Treat. 2009, 10, 265–271. [Google Scholar] [CrossRef]
- Meyer, P.; Mayer, A.; Kulozik, U. High concentration of skim milk proteins by ultrafiltration: Characterisation of a dynamic membrane system with a rotating membrane in comparison with a spiral wound membrane. Int. Dairy J. 2015, 51, 75–83. [Google Scholar] [CrossRef]
- Lebreton, B.; Brown, A.; van Reis, R. Application of high-performance tangential flow filtration (HPTFF) to the purification of a human pharmaceutical antibody fragment expressed in Escherichia coli. Biotechnol. Bioeng. 2008, 100, 964–974. [Google Scholar] [CrossRef]
- Rao, S.; Ager, K.; Zydney, A.L. High performance tangential flow filtration using charged affinity ligands. Sep. Sci. Technol. 2007, 42, 2365–2385. [Google Scholar] [CrossRef]
- Van Reis, R.; Saksena, S. Optimization diagram for membrane separations. J. Memb. Sci. 1997, 129, 19–29. [Google Scholar] [CrossRef]
- Pujar, N.S.; Zydney, A.L. Electrostatic effects on protein partitioning in size-exclusion chromatography and membrane ultrafiltration. J. Chromatogr. A 1998, 796, 229–238. [Google Scholar] [CrossRef]
- Burns, D.B.; Zydney, A.L. Effect of solution pH on protein transport through ultrafiltration membranes. Biotechnol. Bioeng. 1999, 64, 27–37. [Google Scholar] [CrossRef]
- Van Eijndhoven, R.H.C.M.; Saksena, S.; Zydney, A.L. Protein fractionation using electrostatic interactions in membranel filtration. Biotechnol. Bioeng. 1995, 48, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Van Reis, R.; Brake, J.M.; Charkoudian, J.; Burns, D.B.; Zydney, A.L. High-performance tangential flow filtration using charged membranes. J. Memb. Sci. 1999, 159, 133–142. [Google Scholar] [CrossRef]
- Das, B.; Sarkar, S.; Sarkar, A.; Bhattacharjee, S.; Bhattacharjee, C. Recovery of whey proteins and lactose from dairy waste: A step towards green waste management. Process. Saf. Environ. Prot. 2016, 101, 27–33. [Google Scholar] [CrossRef]
- Kanani, D.M.; Ghosh, R.; Filipe, C.D.M. A novel approach for high-resolution protein–protein separation by ultrafiltration using a dual-facilitating agent. J. Memb. Sci. 2004, 243, 223–228. [Google Scholar] [CrossRef]
- Rao, S.; Zydney, A.L. High resolution protein separations using affinity ultrafiltration with small charged ligands. J. Memb. Sci. 2006, 280, 781–789. [Google Scholar] [CrossRef]
- Rao, S.; Zydney, A.L. Controlling protein transport in ultrafiltration using small charged ligands. Biotechnol. Bioeng. 2005, 91, 733–742. [Google Scholar] [CrossRef]
- Bourseau, P.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Massé, A.; Guérard, F.; Chabeaud, A.; Fouchereau-Péron, M.; Le Gal, Y.; Ravallec-Plé, R.; et al. Fractionation of fish protein hydrolysates by ultrafiltration and nanofiltration: Impact on peptidic populations. Desalination 2009, 244, 303–320. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Cao, W.; Ding, L.; Zhu, Z.; Wan, Y.; Jaffrin, M.Y. Treatment of dairy effluent by shear-enhanced membrane filtration: The role of foulants. Sep. Purif. Technol. 2012, 96, 194–203. [Google Scholar] [CrossRef]
- Saidi, S.; Deratani, A.; Ben Amar, R.; Belleville, M.-P. Fractionation of a tuna dark muscle hydrolysate by a two-step membrane process. Sep. Purif. Technol. 2013, 108, 28–36. [Google Scholar] [CrossRef]
- Vandanjon, L.; Grignon, M.; Courois, E.; Bourseau, P.; Jaouen, P. Fractionating white fish fillet hydrolysates by ultrafiltration and nanofiltration. J. Food Eng. 2009, 95, 36–44. [Google Scholar] [CrossRef]
- Field, R.W.; Md Yunos, K.F.; Cui, Z. Separation of proteins using sandwich membranes. Desalination 2009, 245, 597–605. [Google Scholar] [CrossRef]
- Md Yunos, K.F.; Field, R.W. Rejection amplification in the ultrafiltration of binary protein mixtures using sandwich configurations. Chem. Eng. Process. Process. Intensif. 2008, 47, 1053–1060. [Google Scholar] [CrossRef]
- Feins, M.; Sirkar, K.K. Novel internally staged ultrafiltration for protein purification. J. Memb. Sci. 2005, 248, 137–148. [Google Scholar] [CrossRef]
- Roslan, J.; Mustapa Kamal, S.M.; Yunos, K.F.M.; Abdullah, N. Assessment on multilayer ultrafiltration membrane for fractionation of tilapia by-product protein hydrolysate with angiotensin I-converting enzyme (ACE) inhibitory activity. Sep. Purif. Technol. 2017, 173, 250–257. [Google Scholar] [CrossRef]
- Patil, N.V.; Janssen, A.E.M.; Boom, R.M. Separation of whey proteins using cascaded ultrafiltration. Sep. Sci. Technol. 2014, 49, 2280–2288. [Google Scholar] [CrossRef]
- Yang, J.; Guo, J.; Yang, X.-Q.; Wu, N.-N.; Zhang, J.-B.; Hou, J.-J.; Zhang, Y.-Y.; Xiao, W.-K. A novel soy protein isolate prepared from soy protein concentrate using jet-cooking combined with enzyme-assisted ultra-filtration. J. Food Eng. 2014, 143, 25–32. [Google Scholar] [CrossRef]
- Krishna Kumar, N.S.; Yea, M.K.; Cheryan, M. Ultrafiltration of soy protein concentrate: Performance and modelling of spiral and tubular polymeric modules. J. Memb. Sci. 2004, 244, 235–242. [Google Scholar] [CrossRef]
- Marcelo, P.A.; Rizvi, S.S.H. Applications of membrane technology in the dairy industry. In Handbook of Membrane Separation: Chemical, Pharmaceutical, Food, and Biotechnological Applications; Pabby, A., Rizvi, S., Sastre, A., Eds.; CRC Press: New York, NY, USA, 2009; pp. 636–669. [Google Scholar]
- Mondor, M.; Tuyishime, O.; Drolet, H. Production of pea protein concentrates by ultrafiltration: Influence of hollow-fibre module. Innov. Food Sci. Emerg. Technol. 2012, 14, 135–138. [Google Scholar] [CrossRef]
- Almécija, M.C.; Ibáñez, R.; Guadix, A.; Guadix, E.M. Effect of pH on the fractionation of whey proteins with a ceramic ultrafiltration membrane. J. Memb. Sci. 2007, 288, 28–35. [Google Scholar] [CrossRef]
- Van der Marel, P.; Zwijnenburg, A.; Kemperman, A.; Wessling, M.; Temmink, H.; van der Meer, W. Influence of membrane properties on fouling in submerged membrane bioreactors. J. Memb. Sci. 2010, 348, 66–74. [Google Scholar] [CrossRef]
- Zhao, C.; Xue, J.; Ran, F.; Sun, S. Modification of polyethersulfone membranes—A review of methods. Prog. Mater. Sci. 2013, 58, 76–150. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Guérard, F. Fractionation by ultrafiltration of a saithe protein hydrolysate (Pollachius virens): Effect of material and molecular weight cut-off on the membrane performances. J. Food Eng. 2009, 91, 408–414. [Google Scholar] [CrossRef]
- Del Rosario, E.J.; Duldulao, F.D. Preparation and characterization of polysulfone ultrafiltration membranes for concentrating waste tuna broth. Desalination 1988, 70, 2930298. [Google Scholar] [CrossRef]
- Leberknight, J.; Wielenga, B.; Lee-Jewett, A.; Menkhaus, T.J. Recovery of high value protein from a corn ethanol process by ultrafiltration and an exploration of the associated membrane fouling. J. Memb. Sci. 2011, 366, 405–412. [Google Scholar] [CrossRef]
- Damar Huner, I.; Gulec, H.A. Fouling behavior of poly(ether)sulfone ultrafiltration membrane during concentration of whey proteins: Effect of hydrophilic modification using atmospheric pressure argon jet plasma. Colloids Surf. B Biointerfaces 2017, 160, 510–519. [Google Scholar] [CrossRef]
- Susanto, H.; Arafat, H.; Janssen, E.M.L.; Ulbricht, M. Ultrafiltration of polysaccharide-protein mixtures: Elucidation of fouling mechanisms and fouling control by membrane surface modification. Sep. Purif. Technol. 2008, 63, 558–565. [Google Scholar] [CrossRef]
- Sayed Razavi, S.K.; Harris, J.L.; Sherkat, F. Fouling and cleaning of membranes in the ultrafiltration of the aqueous extract of soy flour. J. Memb. Sci. 1996, 114, 93–104. [Google Scholar] [CrossRef]
- Cowan, S.; Ritchie, S. Modified polyethersulfone (PES) ultrafiltration membranes for enhanced filtration of whey proteins. Sep. Sci. Technol. 2007, 42, 2405–2418. [Google Scholar] [CrossRef]
- Saksena, S.; Zydney, A.L. Effect of solution pH and ionic strength on the separation of albumin from immunoglobulins (IgG) by selective filtration. Biotechnol. Bioeng. 1994, 43, 960–968. [Google Scholar] [CrossRef]
- Lucas, D.; Rabiller-Baudry, M.; Millesime, L.; Chaufer, B.; Daufin, G. Extraction of α-lactalbumin from whey protein concentrate with modified inorganic membranes. J. Memb. Sci. 1998, 148, 1–12. [Google Scholar] [CrossRef]
- Weigert, S.; Sára, M. Surface modification of an ultrafiltration membrane with crystalline structure and studies on interactions with selected protein molecules. J. Memb. Sci. 1995, 106, 147–159. [Google Scholar] [CrossRef]
- Stevens, P.V.; Nyström, M.; Ehsani, N. Modification of ultrafiltration membrane with gelatin protein. Biotechnol. Bioeng. 1998, 57, 26–34. [Google Scholar] [CrossRef]
- Yogarathinam, L.T.; Gangasalam, A.; Ismail, A.F.; Arumugam, S.; Narayanan, A. Concentration of whey protein from cheese whey effluent using ultrafiltration by combination of hydrophilic metal oxides and hydrophobic polymer. J. Chem. Technol. Biotechnol. 2018, 93, 2576–2591. [Google Scholar] [CrossRef]
- Kanani, D. Membrane fouling: A challenge during dairy ultrafiltration. In Membrane Processessing for Dairy Ingredient Separation; Hu, K., Dickson, J.M., Eds.; Wiley: Hoboken, NJ, USA, 2015; pp. 67–85. [Google Scholar]
- Aimar, P.; Howell, J.A.; Clifton, M.J.; Sanchez, V. Concentration polarisation build-up in hollow fibers: A method of measurement and its modelling in ultrafiltration. J. Memb. Sci. 1991, 59, 81–99. [Google Scholar] [CrossRef]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Chaplain-Derouiniot, M.; Guerard, F. Performances of ultrafiltration membranes for fractionating a fish protein hydrolysate: Application to the refining of bioactive peptidic fractions. Sep. Purif. Technol. 2009, 66, 463–471. [Google Scholar] [CrossRef]
- Cheryan, M. Ultrafiltration and Microfiltration Handbook, 1st ed.; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]
- Sannier, F.; LeCoeur, C.; Zhao, Q.; Garreau, I.; Piot, J.M. Separation of hemoglobin and myoglobin from yellowfin tuna red muscle by ultrafiltration: Effect of pH and ionic strength. Biotechnol. Bioeng. 1996, 52, 501–506. [Google Scholar] [CrossRef]
- Novin, D.; Md Yunos, K.F. Effect of operating parameters on performance of ultrafiltration (UF) to fractionate catfish protein hydrolysate. J. Teknol. 2014, 70, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Nigam, M.O.; Bansal, B.; Chen, X.D. Fouling and cleaning of whey protein concentrate fouled ultrafiltration membranes. Desalination 2008, 218, 313–322. [Google Scholar] [CrossRef]
- Ali, F.; Mondor, M.; Ippersiel, D.; Lamarche, F. Production of low-phytate soy protein isolate by membrane technologies: Impact of salt addition to the extract on the purification process. Innov. Food Sci. Emerg. Technol. 2011, 12, 171–177. [Google Scholar] [CrossRef]
- Martinez-Ferez, A.; Guadix, A.; Zapata-Montoya, J.E.; Guadix, E.M. Influence of transmembrane pressure on the separation of caprine milk oligosaccharides from protein by cross-flow ultrafiltration. Int. J. Dairy Technol. 2008, 61, 333–339. [Google Scholar] [CrossRef]
- Rajabzadeh, A.R.; Moresoli, C.; Marcos, B. Fouling behavior of electroacidified soy protein extracts during cross-flow ultrafiltration using dynamic reversible-irreversible fouling resistances and CFD modeling. J. Memb. Sci. 2010, 361, 191–205. [Google Scholar] [CrossRef]
- Rabiller-Baudry, M.; Gesan-Guiziou, G.; Roldan-Calbo, D.; Beaulieu, S.; Michel, F. Limiting flux in skimmed milk ultrafiltration: Impact of electrostatic repulsion due to casein micelles. Desalination 2005, 175, 49–59. [Google Scholar] [CrossRef]
- Koumfieg Noudou, V.Y.; Suwal, S.; Amiot, J.; Mikhaylin, S.; Beaulieu, L.; Bazinet, L. Simultaneous electroseparation of anionic and cationic peptides: Impact of feed peptide concentration on migration rate, selectivity and relative energy consumption. Sep. Purif. Technol. 2016, 157, 53–59. [Google Scholar] [CrossRef]
- Rohani, M.M.; Zydney, A.L. Role of electrostatic interactions during protein ultrafiltration. Adv. Colloid Interface Sci. 2010, 160, 40–48. [Google Scholar] [CrossRef]
- Palecek, S.P.; Zydney, A.L. Intermolecular electrostatic interactions and their effect on flux and protein deposition during protein filtration. Biotechnol. Prog. 1994, 10, 207–213. [Google Scholar] [CrossRef]
- Fane, A.G.; Fell, C.J.D.; Waters, A.G. Ultrafiltration of protein solutions through partially permeable membranes—The effect of adsorption and solution environment. J. Memb. Sci. 1983, 16, 211–224. [Google Scholar] [CrossRef]
- Lai, Y.P.; Mondor, M.; Moresoli, C.; Drolet, H.; Gros-Louis, M.; Ippersiel, D.; Lamarche, F.; Arcand, Y. Production of soy protein isolates with low phytic acid content by membrane technologies: Impact of the extraction and ultrafiltration/diafiltration conditions. J. Food Eng. 2013, 114, 221–227. [Google Scholar] [CrossRef]
- Mondor, M.; Aksay, S.; Drolet, H.; Roufik, S.; Farnworth, E.; Boye, J.I. Influence of processing on composition and antinutritional factors of chickpea protein concentrates produced by isoelectric precipitation and ultrafiltration. Innov. Food Sci. Emerg. Technol. 2009, 10, 342–347. [Google Scholar] [CrossRef]
- Roblet, C.; Doyen, A.; Amiot, J.; Bazinet, L. Impact of pH on ultrafiltration membrane selectivity during electrodialysis with ultrafiltration membrane (EDUF) purification of soy peptides from a complex matrix. J. Memb. Sci. 2013, 435, 207–217. [Google Scholar] [CrossRef]
- Suwal, S.; Roblet, C.; Amiot, J.; Doyen, A.; Beaulieu, L.; Legault, J.; Bazinet, L. Recovery of valuable peptides from marine protein hydrolysate by electrodialysis with ultrafiltration membrane: Impact of ionic strength. Food Res. Int. 2014, 65, 407–415. [Google Scholar] [CrossRef]
- Mondor, M.; Ippersiel, D.; Lamarche, F.; Boye, J.I. Effect of electro-acidification treatment and ionic environment on soy protein extract particle size distribution and ultrafiltration permeate flux. J. Memb. Sci. 2004, 231, 169–179. [Google Scholar] [CrossRef]
- Mukai, Y.; Iritani, E.; Murase, T. Effect of protein charge on cake properties in dead-end ultrafiltration of protein solutions. J. Memb. Sci. 1997, 137, 271–275. [Google Scholar] [CrossRef]
- Schock, G.; Miquel, A.; Birkenberger, R. Characterization of ultrafiltration membranes: Cut-off determination by gel permeation chromatography. J. Memb. Sci. 1989, 41, 55–67. [Google Scholar] [CrossRef]
- Tsakali, E.; Petrotos, K.; D’Alessandro, A.G.; Mantas, C.; Tripolitsiotis, I.; Goulas, P.; Chatzilazarou, A.; Van Impe, J.F. Exploring the effect of ultrafiltration/diafiltration processing conditions on the Lactoferrin and Immunoglobulin G content of feta whey protein concentrates. J. Food Process. Eng. 2015, 38, 363–373. [Google Scholar] [CrossRef]
- Cassano, A.; Donato, L.; Drioli, E. Ultrafiltration of kiwifruit juice: Operating parameters, juice quality and membrane fouling. J. Food Eng. 2007, 79, 613–621. [Google Scholar] [CrossRef]
- Yan, S.H.; Hill, C.G.; Amundson, C.H. Ultrafiltration of whole milk. J. Dairy Sci. 1979, 62, 23–40. [Google Scholar] [CrossRef]
- Luo, X.; Ramchandran, L.; Vasiljevic, T. Lower ultrafiltration temperature improves membrane performance and emulsifying properties of milk protein concentrates. Dairy Sci. Technol. 2015, 95, 15–31. [Google Scholar] [CrossRef] [Green Version]







| Process | Feeds | Membrane and/or Protocols | Remarks | Ref |
|---|---|---|---|---|
| UF with charged membrane | Whey protein | Regenerated cellulose (RC), negatively charged 100 kDa (flat sheet/spiral-wound) | Both modules retain 98% of total whey proteins 85% higher flux than 10 kDa pristine membrane | [62] |
| α-lactalbumin and β-lactoglobulin | RC, positively charged 300 kDa (two-staged UF) | 490% improvement of selectivity to retain β-lactoglobulin. Purity of α-lactalbumin: 87% | [70] | |
| Milk serum permeate | RC, positively charged 300 kDa (three-staged UF) | 180% improvement of selectivity to retain β-lactoglobulin. Purity of α-lactalbumin: 87% | [71] | |
| Lysozyme and lactoferrin | Zirconia, positively grafted 300 kDa by ethylenediamine | Lysozyme transmission selectivity up to 165 100% purity in permeate | [72] | |
| EUF | BSA | Continuous current, PS (polysulfone) 100 kDa, effective area 45 cm2, 3000 V m−1 | Reduction of 80% in BSA concentration time (from 0.5–1.0 g L−1) | [73] |
| BSA and lysozyme | External DC current, PES (polyethersulfone) 30 kDa, effective filtration area: 132 cm2, 1000 V m−1 | Decline of lysozyme retention by 53% Permeate flux increased around 23.4–36.7 L m−2 h−1 | [74] | |
| Pulsed electric fields, fluoride polyvinylden flat sheet 25 kDa, effective filtration area: 51 cm2, 700 V m−1 | A 300% increase in permeate flux Reduction of membrane abrasion | [75] | ||
| BSA and lysozyme | Electrodialysis integration, UF membrane: PES 30 kDa, effective filtration area: 35 cm2, 150 mA | Permeate flux increase by 20% while operated in high conductivity feed | [76] | |
| EDUF | α-lactalbumin and β-lactoglobulin | CA, 100 kDa (elution mode), membrane active area: 35 cm2, 3–22 A m−2, 100–800 V m−1 | Separation factor of 1.2 reached at pH 4.8 (nearby IEP of α-lactalbumin) (optimum) | [77] |
| α-lactalbumin and bovine hemoglobin | CA (100 kDa), membrane active area: 32 cm2, 3–20 A m−2, 170–800 V m−1 | Elution mode: 99% purity of α-lactalbumin Separation mode: 20% increase in yield | [78] | |
| Herring milt hydrolysate | Electrocell unit (membrane active area: 10 cm2), UF membrane 20 kDa, 11 Vcm−1 | 214% increase in antioxidant activity | [79] | |
| EDBM | Soy protein hydrolysate | Electrocell AB unit (eff electrode area: 100 cm2) UF integration (hydrophilic RC 100 kDa, 13.4 cm2) | Soy protein concentrate 70% with phytate below 1% Eliminate clogging, but not fouling | [80] |
| Lactose-enriched hydrolysate | New 6-compartment EDBM, Electrocell AB unit (eff electrode area: 100 cm2), UF membrane: spiral-wound 10 kDa (area 2.13 m2) | Complete reduction of scaling | [81] | |
| Ultrasonic-assisted UF | Whey protein concentrate | PES 30 kDa and permanently hydrophilic PES 5 kDa (effective area 100 cm2), 20 kHz, 300 W (low frequency) | Reduce irreversible fouling and increase cleaning efficiency up to 17.23% (5 kDa) and 5.47% (30 kDa) | [82] |
| Dynamic UF | Mungbean wastewater | RDM: ceramic (MgAl2O4) 15 kDa (eff membrane area: 360 cm2), TMP 1.2 bar, 1403 rpm, and VRF 5.0 (optimum) | Protein recovery by tenfold higher flux at 42 L m−2 h−1 than conventional cross-flow UF. Protein retention: 96% | [83] |
| UHT milk | VSEP: PES 10 kDa, effective area: 503 cm2, 60.75 Hz, 45 °C | Operation at higher VRF (9.0) than cross-flow UF with nominal commercial flux (20 L m−2 h−1) | [84] | |
| HPTFF | Chicken egg white | 30 kDa PES, 2100 rpm, pH 10–11, and NaCl 100 mM | Lysozyme–ovalbumin (different in size and charge) attained high resolution (transmission: 99%, selectivity: 2400; flux of 25.6 L m−2 h−1) | [85] |
| Monoclonal antibody Alemtuzumab | PES 300 and 100 kDa, PVDF 100 kDa constant flux: 7.368 × 10−6 m s−1 | Only PVDF 100 kDa gave 93.3% purity for monomer (155 kDa) after one-staged diafiltration | [86] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratnaningsih, E.; Reynard, R.; Khoiruddin, K.; Wenten, I.G.; Boopathy, R. Recent Advancements of UF-Based Separation for Selective Enrichment of Proteins and Bioactive Peptides—A Review. Appl. Sci. 2021, 11, 1078. https://doi.org/10.3390/app11031078
Ratnaningsih E, Reynard R, Khoiruddin K, Wenten IG, Boopathy R. Recent Advancements of UF-Based Separation for Selective Enrichment of Proteins and Bioactive Peptides—A Review. Applied Sciences. 2021; 11(3):1078. https://doi.org/10.3390/app11031078
Chicago/Turabian StyleRatnaningsih, Enny, Reynard Reynard, Khoiruddin Khoiruddin, I Gede Wenten, and Ramaraj Boopathy. 2021. "Recent Advancements of UF-Based Separation for Selective Enrichment of Proteins and Bioactive Peptides—A Review" Applied Sciences 11, no. 3: 1078. https://doi.org/10.3390/app11031078