Wheat Germ Agglutinin—From Toxicity to Biomedical Applications


Abstract
:Featured Application
Abstract
1. Introduction
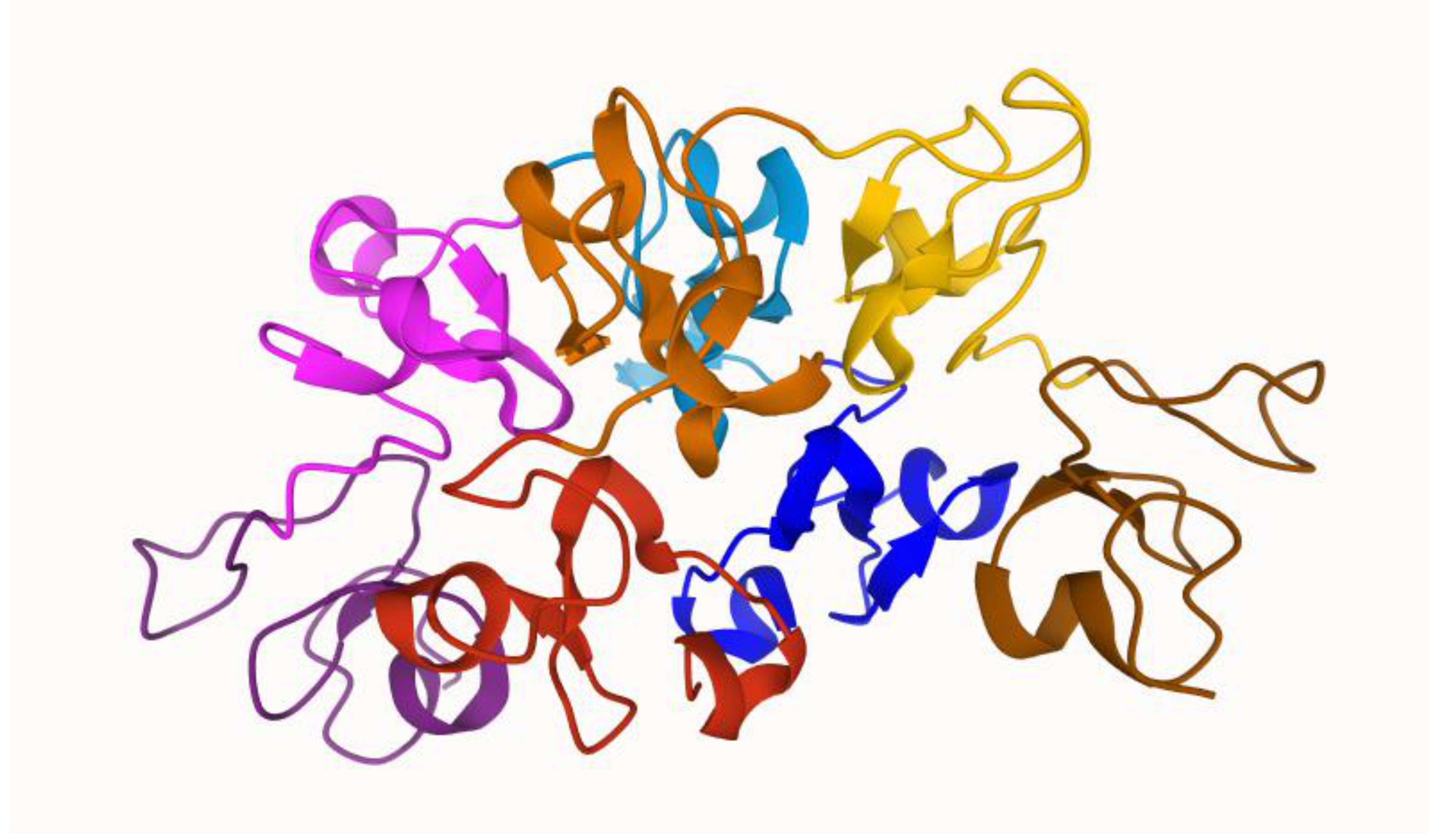
2. Wheat Germ Agglutinin Structure
3. WGA Extraction and Purification
4. WGA Biological Activity
4.1. Antifungal Activity
4.2. Cytotoxicity
5. WGA Biomedical Perspectives
5.1. Advanced Drug Delivery
5.2. Overcoming Physiological Barriers
6. Non-Medical WGA Applications
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Enghiad, A.; Ufer, D.; Countryman, A.M.; Thilmany, D.D. An Overview of Global Wheat Market Fundamentals in an Era of Climate Concerns. Available online: https://www.hindawi.com/journals/ija/2017/3931897/ (accessed on 1 April 2020).
- Bacenetti, J.; Duca, D.; Negri, M.; Fusi, A.; Fiala, M. Mitigation Strategies in the Agro-Food Sector: The Anaerobic Digestion of Tomato Purée by-Products. An Italian Case Study. Sci. Total Environ. 2015, 526, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Šramková, Z.; Gregová, E.; Šturdík, E. Chemical Composition and Nutritional Quality of Wheat Grain. Acta Chim. Slovaca 2009, 2, 115–138. [Google Scholar]
- Edwin Geo, V.; Prabhu, C.; Thiyagarajan, S.; Maiyalagan, T.; Aloui, F. Comparative Analysis of Various Techniques to Improve the Performance of Novel Wheat Germ Oil—An Experimental Study. Int. J. Hydrog. Energy 2020, 45, 5745–5756. [Google Scholar] [CrossRef]
- Boukid, F.; Folloni, S.; Ranieri, R.; Vittadini, E. A Compendium of Wheat Germ: Separation, Stabilization and Food Applications. Trends Food Sci. Technol. 2018, 78, 120–133. [Google Scholar] [CrossRef]
- Teslić, N.; Bojanić, N.; Rakić, D.; Takači, A.; Zeković, Z.; Fišteš, A.; Bodroža-Solarov, M.; Pavlić, B. Defatted Wheat Germ as Source of Polyphenols—Optimization of Microwave-Assisted Extraction by RSM and ANN Approach. Chem. Eng. Process.-Process Intensif. 2019, 143, 107634. [Google Scholar] [CrossRef]
- Schwefel, D.; Maierhofer, C.; Beck, J.G.; Seeberger, S.; Diederichs, K.; Möller, H.M.; Welte, W.; Wittmann, V. Structural Basis of Multivalent Binding to Wheat Germ Agglutinin. J. Am. Chem. Soc. 2010, 132, 8704–8719. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Arntfield, S.D.; Nickerson, M. Changes in Levels of Phytic Acid, Lectins and Oxalates during Soaking and Cooking of Canadian Pulses. Food Res. Int. 2018, 107, 660–668. [Google Scholar] [CrossRef]
- Smith, J.J.; Raikhel, N.V. Nucleotide Sequences of CDNA Clones Encoding Wheat Germ Agglutinin Isolectins A and D. Plant Mol. Biol. 1989, 13, 601–603. [Google Scholar] [CrossRef]
- Peumans, W.J.; Stinissen, H.M.; Carlier, A.R. Isolation and Partial Characterization of Wheat-Germ-Agglutinin-like Lectins from Rye (Secale Cereale) and Barley (Hordeum Vulgare) Embryos. Biochem. J. 1982, 203, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Rice, R.H.; Etzler, M.E. Subunit Structure of Wheat Germ Agglutinin. Biochem. Biophys. Res. Commun. 1974, 59, 414–419. [Google Scholar] [CrossRef]
- Wright, C.S.; Raikhel, N. Sequence Variability in Three Wheat Germ Agglutinin Isolectins: Products of Multiple Genes in Polyploid Wheat. J. Mol. Evol. 1989, 28, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Matucci, A.; Veneri, G.; Dalla Pellegrina, C.; Zoccatelli, G.; Vincenzi, S.; Chignola, R.; Peruffo, A.D.B.; Rizzi, C. Temperature-Dependent Decay of Wheat Germ Agglutinin Activity and Its Implications for Food Processing and Analysis. Food Control 2004, 15, 391–395. [Google Scholar] [CrossRef]
- Portillo-Téllez, M.D.C.; Bello, M.; Salcedo, G.; Gutiérrez, G.; Gómez-Vidales, V.; García-Hernández, E. Folding and Homodimerization of Wheat Germ Agglutinin. Biophys. J. 2011, 101, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Romero, A.; Ravichandran, K.G.; Soriano-García, M. Crystal Structure of Hevein at 2.8 Å Resolution. Febs. Lett. 1991, 291, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Leyva, E.; Medrano-Cerano, J.L.; Cano-Sánchez, P.; López-González, I.; Gómez-Velasco, H.; Del Río-Portilla, F.; García-Hernández, E. Bacterial Expression, Purification and Biophysical Characterization of Wheat Germ Agglutinin and Its Four Hevein-like Domains. Biopolymers 2019, 110, e23242. [Google Scholar] [CrossRef]
- Nagata, Y.; Goldberg, A.R.; Burger, M.M. The isolation and purification of wheat germ and other agglutinins. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1974; Volume 32, pp. 611–615. ISBN 978-0-12-181895-1. [Google Scholar]
- Wright, H.T.; Sandrasegaram, G.; Wright, C.S. Evolution of a Family of N-Acetylglucosamine Binding Proteins Containing the Disulfide-Rich Domain of Wheat Germ Agglutinin. J. Mol. Evol. 1991, 33, 283–294. [Google Scholar] [CrossRef]
- Hernández-Arana, A.; Rojo-Domínguez, A.; Soriano-García, M.; Rodríguez-Romero, A. The Thermal Unfolding of Hevein, a Small Disulfide-Rich Protein. Eur. J. Biochem. 1995, 228, 649–652. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Peumans, W.J.; Raikhel, N.V. Wheat-Germ Agglutinin Is Synthesized as a Glycosylated Precursor. Planta 1988, 173, 482–489. [Google Scholar] [CrossRef]
- Allen, A.K.; Neuberger, A.; Sharon, N. The Purification, Composition and Specificity of Wheat-Germ Agglutinin. Biochem. J. 1973, 131, 155–162. [Google Scholar] [CrossRef]
- Gallagher, J.T.; Morris, A.; Dexter, T.M. Identification of Two Binding Sites for Wheat-Germ Agglutinin on Polylactosamine-Type Oligosaccharides. Biochem. J. 1985, 231, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Peumans, W.J.; van Damme, E.J.M.; Barre, A.; Rougé, P. Classification of Plant Lectins in Families Of Structurally and Evolutionary Related Proteins. Adv. Exp. Med. Biol. 2001, 491, 27–54. [Google Scholar] [CrossRef] [PubMed]
- Harata, K.; Nagahora, H.; Jigami, Y. X-ray Structure of Wheat Germ Agglutinin Isolectin 3. Acta Cryst. D 1995, 51, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Lotan, R.; Gussin, A.E.S.; Lis, H.; Sharon, N. Purification of Wheat Germ Agglutinin by Affinity Chromatography on a Sepharose-Bound N-Acetylglucosamine Derivative. Biochem. Biophys. Res. Commun. 1973, 52, 656–662. [Google Scholar] [CrossRef]
- Baieli, M.F.; Urtasun, N.; Miranda, M.V.; Cascone, O.; Wolman, F.J. Efficient Wheat Germ Agglutinin Purification with a Chitosan-Based Affinity Chromatographic Matrix. J. Sep. Sci. 2012, 35, 231–238. [Google Scholar] [CrossRef]
- Ciopraga, J.; Gozia, O.; Tudor, R.; Brezuica, L.; Doyle, R.J. Fusarium Sp. Growth Inhibition by Wheat Germ Agglutinin. Biochim. Biophys. Acta 1999, 1428, 424–432. [Google Scholar] [CrossRef]
- Tonkal, A.M. In Vitro Antitrichomonal Effect of Nigella Sativa Aqueous Extract and Wheat Germ Agglutinin. J. King Abdulaziz Univ. Med. Sci. 2009, 16, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Liedke, S.C.; Miranda, D.Z.; Gomes, K.X.; Gonçalves, J.L.S.; Frases, S.; Nosanchuk, J.D.; Rodrigues, M.L.; Nimrichter, L.; Peralta, J.M.; Guimarães, A.J. Characterization of the Antifungal Functions of a WGA-Fc (IgG2a) Fusion Protein Binding to Cell Wall Chitin Oligomers. Sci. Rep. 2017, 7, 12187. [Google Scholar] [CrossRef]
- da Silva, P.M.; de Moura, M.C.; Gomes, F.S.; da Silva Trentin, D.; Silva de Oliveira, A.P.; de Mello, G.S.V.; da Rocha Pitta, M.G.; de Melo Rego, M.J.B.; Coelho, L.C.B.B.; Macedo, A.J.; et al. PgTeL, the Lectin Found in Punica Granatum Juice, Is an Antifungal Agent against Candida Albicans and Candida Krusei. Int. J. Biol. Macromol. 2018, 108, 391–400. [Google Scholar] [CrossRef]
- Kanokwiroon, K.; Teanpaisan, R.; Wititsuwannakul, D.; Hooper, A.B.; Wititsuwannakul, R. Antimicrobial Activity of a Protein Purified from the Latex of Hevea Brasiliensis on Oral Microorganisms. Mycoses 2008, 51, 301–307. [Google Scholar] [CrossRef]
- Dickers, K.J.; Bradberry, S.M.; Rice, P.; Griffiths, G.D.; Vale, J.A. Abrin Poisoning. Toxicol. Rev. 2003, 22, 137–142. [Google Scholar] [CrossRef]
- Lopez Nunez, O.F.; Pizon, A.F.; Tamama, K. Ricin Poisoning after Oral Ingestion of Castor Beans: A Case Report and Review of the Literature and Laboratory Testing. J. Emerg. Med. 2017, 53, e67–e71. [Google Scholar] [CrossRef]
- Gorelik, E. Mechanisms of Cytotoxic Activity of Lectins. Trends Glycosci. Glycotechnol. 1994, 6, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.; Sadofsky, L.; Hart, S. Peripheral Blood Leukocyte Immune Responses Are Distinctly Altered in Sarcoidosis. Eur. Respir. J. 2015, 46. [Google Scholar] [CrossRef]
- Clevers, H.C.; de Bresser, A.; Kleinveld, H.; Gmelig-Meyling, F.H.; Ballieux, R.E. Wheat Germ Agglutinin Activates Human T Lymphocytes by Stimulation of Phosphoinositide Hydrolysis. J. Immunol. 1986, 136, 3180–3183. [Google Scholar] [PubMed]
- Kawakami, K.; Yamamoto, Y.; Onoue, K. Effect of Wheat Germ Agglutinin on T Lymphocyte Activation. Microbiol. Immunol. 1988, 32, 413–422. [Google Scholar] [CrossRef]
- Reed, J.C.; Robb, R.J.; Greene, W.C.; Nowell, P.C. Effect of Wheat Germ Agglutinin on the Interleukin Pathway of Human T Lymphocyte Activation. J. Immunol. 1985, 134, 314–323. [Google Scholar]
- Ryva, B.; Zhang, K.; Asthana, A.; Wong, D.; Vicioso, Y.; Parameswaran, R. Wheat Germ Agglutinin as a Potential Therapeutic Agent for Leukemia. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Boligan, K.F.; Mesa, C.; Fernandez, L.E.; von Gunten, S. Cancer Intelligence Acquired (CIA): Tumor Glycosylation and Sialylation Codes Dismantling Antitumor Defense. Cell. Mol. Life Sci. 2015, 72, 1231–1248. [Google Scholar] [CrossRef]
- Pearce, O.M.T.; Läubli, H. Sialic Acids in Cancer Biology and Immunity. Glycobiology 2016, 26, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Raub, T.J.; Koroly, M.J.; Roberts, R.M. Endocytosis of Wheat Germ Agglutinin Binding Sites from the Cell Surface into a Tubular Endosomal Network. J. Cell. Physiol. 1990, 143, 1–12. [Google Scholar] [CrossRef]
- Gastman, B.; Wang, K.; Han, J.; Zhu, Z.; Huang, X.; Wang, G.-Q.; Rabinowich, H.; Gorelik, E. A Novel Apoptotic Pathway as Defined by Lectin Cellular Initiation. Biochem. Biophys. Res. Commun. 2004, 316, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Wirth, C.; Schwuchow, J.; Jonas, L. Internalization of Wheat Germ Agglutinin (WGA) by Rat Pancreatic Cells in Vivo and in Vitro. Acta Histochem. 1996, 98, 165–172. [Google Scholar] [CrossRef]
- Pellegrina, C.D.; Matucci, A.; Zoccatelli, G.; Rizzi, C.; Vincenzi, S.; Veneri, G.; Andrighetto, G.; Peruffo, A.D.B.; Chignola, R. Studies on the Joint Cytotoxicity of Wheat Germ Agglutinin and Monensin. Toxicol. Vitr. 2004, 18, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, E.; Wozniak, R.; Blobel, G. An Integral Membrane Protein of the Pore Membrane Domain of the Nuclear Envelope Contains a Nucleoporin-like Region. J. Cell. Biol. 1993, 122, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Hanover, J.A. Functional Nuclear Pores Reconstituted with Beta 1-4 Galactose-Modified O-Linked N-Acetylglucosamine Glycoproteins. J. Biol. Chem. 1994, 269, 9289–9297. [Google Scholar] [CrossRef]
- Heese-Peck, A.; Cole, R.N.; Borkhsenious, O.N.; Hart, G.W.; Raikhel, N.V. Plant Nuclear Pore Complex Proteins Are Modified by Novel Oligosaccharides with Terminal N-Acetylglucosamine. Plant Cell 1995, 7, 1459–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, C.; Nebe, B.; Walzel, H.; Weber, H.; Jonas, L. Inhibitory Effect of the Lectin Wheat Germ Agglutinin (WGA) on the Proliferation of AR42J Cells. Acta Histochem. 2009, 111, 336–343. [Google Scholar] [CrossRef]
- Tsai, T.L.; Wang, H.C.; Hung, C.H.; Lin, P.C.; Lee, Y.S.; Chen, H.H.W.; Su, W.C. Wheat Germ Agglutinin-Induced Paraptosis-like Cell Death and Protective Autophagy Is Mediated by Autophagy-Linked FYVE Inhibition. Oncotarget 2017, 8, 91209–91222. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, R.E.; Wojciechowicz, D.C.; Picon, A.I.; Schwarz, M.A.; Paty, P.B. Wheatgerm Agglutinin-Mediated Toxicity in Pancreatic Cancer Cells. Br. J. Cancer 1999, 80, 1754–1762. [Google Scholar] [CrossRef] [Green Version]
- Garcia, M.; Seigner, C.; Bastid, C.; Choux, R.; Payan, M.J.; Reggio, H. Carcinoembryonic Antigen Has a Different Molecular Weight in Normal Colon and in Cancer Cells Due to N-Glycosylation Differences. Cancer Res. 1991, 51, 5679–5686. [Google Scholar]
- Weissenböck, A.; Wirth, M.; Gabor, F. WGA-Grafted PLGA-Nanospheres: Preparation and Association with Caco-2 Single Cells. J. Control. Release 2004, 99, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Lim, L.-Y. Preparation and in Vitro Anticancer Activity of Wheat Germ Agglutinin (WGA)-Conjugated PLGA Nanoparticles Loaded with Paclitaxel and Isopropyl Myristate. J. Control. Release 2005, 107, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ho, P.C.; Lim, L.Y. Wheat Germ Agglutinin-Conjugated PLGA Nanoparticles for Enhanced Intracellular Delivery of Paclitaxel to Colon Cancer Cells. Int. J. Pharm. 2010, 400, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.I.; Ayub, A.D.; Mat Yusuf, S.N.A.; Yahaya, N.; Abd Kadir, E.; Lim, V. Docetaxel-Loaded Disulfide Cross-Linked Nanoparticles Derived from Thiolated Sodium Alginate for Colon Cancer Drug Delivery. Pharmaceutics 2020, 12, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.-C.; Chang, Y.-H.; Rajesh, R. Targeted Delivery of Etoposide, Carmustine and Doxorubicin to Human Glioblastoma Cells Using Methoxy Poly(Ethylene Glycol)-poly(Ε-caprolactone) Nanoparticles Conjugated with Wheat Germ Agglutinin and Folic Acid. Mater. Sci. Eng. C 2019, 96, 114–128. [Google Scholar] [CrossRef] [PubMed]
- Gabor, F.; Schwarzbauer, A.; Wirth, M. Lectin-Mediated Drug Delivery: Binding and Uptake of BSA-WGA Conjugates Using the Caco-2 Model. Int. J. Pharm. 2002, 237, 227–239. [Google Scholar] [CrossRef]
- Murata, M.; Yonamine, T.; Tanaka, S.; Tahara, K.; Tozuka, Y.; Takeuchi, H. Surface Modification of Liposomes Using Polymer-Wheat Germ Agglutinin Conjugates to Improve the Absorption of Peptide Drugs by Pulmonary Administration. J. Pharm. Sci. 2013, 102, 1281–1289. [Google Scholar] [CrossRef]
- Leong, K.H.; Chung, L.Y.; Noordin, M.I.; Onuki, Y.; Morishita, M.; Takayama, K. Lectin-Functionalized Carboxymethylated Kappa-Carrageenan Microparticles for Oral Insulin Delivery. Carbohydr. Polym. 2011, 86, 555–565. [Google Scholar] [CrossRef]
- AlSadek, D.M.M.; Badr, H.A.; Al-Shafie, T.A.; El-Bahr, S.M.; El-Houseini, M.E.; Djansugurova, L.B.; Li, C.-Z.; Ahmed, H. Cancer Cell Death Induced by Nanomagnetolectin. Eur. J. Cell Biol. 2017, 96, 600–611. [Google Scholar] [CrossRef]
- Yang, K.; Gitter, B.; Rüger, R.; Albrecht, V.; Wieland, G.D.; Fahr, A. Wheat Germ Agglutinin Modified Liposomes for the Photodynamic Inactivation of Bacteria†. Photochem. Photobiol. 2012, 88, 548–556. [Google Scholar] [CrossRef]
- Gao, X.; Wu, B.; Zhang, Q.; Chen, J.; Zhu, J.; Zhang, W.; Rong, Z.; Chen, H.; Jiang, X. Brain Delivery of Vasoactive Intestinal Peptide Enhanced with the Nanoparticles Conjugated with Wheat Germ Agglutinin Following Intranasal Administration. J. Control. Release 2007, 121, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Xiao, C.; Davis, F.; Lambert, M.; Viola, K.; Lacor, P.; Dravid, V.; Klein, W. Intranasal Passive Immunization Using WGA-Modified Oligomer Antibodies Greatly Improves Learning and Memory in Alzheimer’s 5XFAD Mice. Alzheimer’s Dement. 2012, 8, P197. [Google Scholar] [CrossRef]
- Zhang, Y.; Walker, J.B.; Minic, Z.; Liu, F.; Goshgarian, H.; Mao, G. Transporter Protein and Drug-Conjugated Gold Nanoparticles Capable of Bypassing the Blood-Brain Barrier. Sci. Rep. 2016, 6, 25794. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, Y.; Jia, X.-R.; Du, J.; Ying, X.; Lu, W.-L.; Lou, J.-N.; Wei, Y. PEGylated Poly(Amidoamine) Dendrimer-Based Dual-Targeting Carrier for Treating Brain Tumors. Biomaterials 2011, 32, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Sun, B.; Gao, X.; Dong, X.; Fu, L.; Zhang, Y.; Li, Z.; Wang, Y.; Jiang, H.; Han, B. Intranasal Delivery of Targeted Nanoparticles Loaded With MiR-132 to Brain for the Treatment of Neurodegenerative Diseases. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Plattner, V.E.; Wagner, M.; Ratzinger, G.; Gabor, F.; Wirth, M. Targeted Drug Delivery: Binding and Uptake of Plant Lectins Using Human 5637 Bladder Cancer Cells. Eur. J. Pharm. Biopharm. 2008, 70, 572–576. [Google Scholar] [CrossRef]
- Apfelthaler, C.; Anzengruber, M.; Gabor, F.; Wirth, M. Poly-(l)-Glutamic Acid Drug Delivery System for the Intravesical Therapy of Bladder Cancer Using WGA as Targeting Moiety. Eur. J. Pharm. Biopharm. 2017, 115, 131–139. [Google Scholar] [CrossRef]
- Brauner, B.; Semmler, J.; Rauch, D.; Nokaj, M.; Haiss, P.; Schwarz, P.; Wirth, M.; Gabor, F. Trimethoprim-Loaded PLGA Nanoparticles Grafted with WGA as Potential Intravesical Therapy of Urinary Tract Infections—Studies on Adhesion to SV-HUCs Under Varying Time, PH, and Drug-Loading Conditions. Acs Omega 2020, 5, 17377–17384. [Google Scholar] [CrossRef]
- Wijetunge, S.S.; Wen, J.; Yeh, C.-K.; Sun, Y. Wheat Germ Agglutinin Liposomes with Surface Grafted Cyclodextrins as Bioadhesive Dual-Drug Delivery Nanocarriers to Treat Oral Cells. Colloids Surf. B Biointerfaces 2020, 185, 110572. [Google Scholar] [CrossRef]
- Liu, S.-L.; Zhang, Z.-L.; Sun, E.-Z.; Peng, J.; Xie, M.; Tian, Z.-Q.; Lin, Y.; Pang, D.-W. Visualizing the Endocytic and Exocytic Processes of Wheat Germ Agglutinin by Quantum Dot-Based Single-Particle Tracking. Biomaterials 2011, 32, 7616–7624. [Google Scholar] [CrossRef]
- Mikaelyan, M.V.; Poghosyan, G.G.; Hendrickson, O.D.; Dzantiev, B.B.; Gasparyan, V.K. Wheat Germ Agglutinin and Lens Culinaris Agglutinin Sensitized Anisotropic Silver Nanoparticles in Detection of Bacteria: A Simple Photometric Assay. Anal. Chim. Acta 2017, 981, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Stanley, S.M.R.; Chua, D. Improved Recovery of Erythropoietin and Darbepoetin from Equine Plasma by the Application of a Wheat Germ Agglutinin Mediated Pre-Extraction Prior to Immunoaffinity Chromatography. Adv. Biosci. Biotechnol. 2014. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Defatting Agent | Purification Strategy | Number of Steps | Yield of WGA (mg Per 1 g Raw Wheat Germ) | Reference |
|---|---|---|---|---|
| Light petroleum | Extraction in water Protein precipitation 55% (NH4)2SO4 Dialysis in water Ion exchange chromatography using a DEAE 1-cellulose column, two SE 2-Sephadex columns, and QAE 3-Sephadex (×4) | 7 | 0.61 | [22] |
| Light petroleum | Extraction in water Protein precipitation 55% (NH4)2SO4 Dialysis in water Affinity chromatography CNAG 4-Sepharose (×1) | 4 | 0.3 | [25] |
| Acetone | Extraction in 0.05 N HC1 buffer Protein precipitation 35% (NH4)2SO4 n-Butanol treatment Dialysis in 0.05 N HC1 and second protein precipitation 35% (NH4)2SO4 Ion exchange chromatography on DEAE-cellulose column (×1) | 6 | 0.48 | [17] |
| n-Hexane | Extraction in 0.05 N HC1 buffer Tangential flow filtration MWCO 5 10 kDa Batch affinity purification on chitosan matrices (×1) | 3 | [26] |
| Carrier Particle | Drug | Excipients | Animal/Cell Line | Outcome | Reference |
|---|---|---|---|---|---|
| Polymeric nanoparticles | Paclitaxel | Poly (lactic-co-glycolic acid) Isopropyl myristate | A549 H1299 CCL-186 cell lines | Demonstrated higher cytotoxicity | [54] |
| Polymeric nanoparticles | Paclitaxel | Poly (lactic-co-glycolic acid) | Caco-2 HT-29 cell lines | Increased intracellular retention in the Caco-2 and HT-29 cells Endocytosed nanoparticles could successfully escape from the endo-lysosome compartment and release into the cytosol with increasing incubation time | [55] |
| Polymeric nanoparticles | Docetaxel | Thiolated sodium alginate | HT-29 L929 cell lines | IC50 values of 52.9 µg/mL for HT-29 cells and 201.6 µg/mL for L929 cells Selectivity towards HT-29 cells over L929 cells | [56] |
| Polymeric nanoparticles | Etoposide Carmustine Doxorubicin | Methoxy poly(ethylene glycol)-poly(ε-caprolactone) Folic acid | U87MG cell line | WGA-modified surface promoted BBB permeation, and folic acid facilitated target site on U87MG cells Anti-proliferation against U87MG cells | [57] |
| Liposomes | Calcitonin | Carbopol | A549 cell line | Carbopol-WGA modification enhanced interaction with A549 lung epithelial cells compared with unmodified or CP-modified liposomes Enhanced and prolonged efficacy of calcitonin. | [59] |
| Microparticles | Insulin | Carboxymethylated kappa–carrageenan microparticles | Caco-2 cell line Sprague–Dawley rats | WGA-functionalized microparticles at 20 mg/mL showed a reduction in cell viability upon exposure The oral administration of insulin entrapped in the microparticles led to a prolonged duration of the hypoglycemic effect, up to 12–24 h, in diabetic rats. | [60] |
| Metal-oxide based nanoparticles | - | Iron oxide Fe3O4 | PZ-HPV-7 DU-145 PC-3 LNCaP cell lines DU-145 e BALB/c-nu/nu mice | 2.46 nM nanomagnet lectin exposure for 15 min induce apoptosis of cancer cells; Xenografted (DU-145) e BALB/c-nu/nu mice, where the tumor was not only completely arrested but also reduced. | [61] |
| Liposomes | Temoporfin | 1,2-dipalmitoyl-sn-glycero-3-phosphocholin N-[1-(2,3-dioleoyloxy)propyl]-N,N,N-trimethylammonium methyl sulfate 1,2- distearoyl-sn-glycero-3-phosphoethanolamine-N-[3-(N-succinimidyloxyglutaryl)aminopropyl(polyethyleneglycol)-2000-carbamyl] | Methicillin-resistant Staphylococcus aureus (MRSA) and Pseudomonas aeruginosa cultures | The WGA-modified liposomes eradicated all MRSA and significantly enhanced the photodynamic inactivation of P. aeruginosa. | [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiūnaitė-Murzienė, G.; Dzikaras, M. Wheat Germ Agglutinin—From Toxicity to Biomedical Applications. Appl. Sci. 2021, 11, 884. https://doi.org/10.3390/app11020884
Balčiūnaitė-Murzienė G, Dzikaras M. Wheat Germ Agglutinin—From Toxicity to Biomedical Applications. Applied Sciences. 2021; 11(2):884. https://doi.org/10.3390/app11020884
Chicago/Turabian StyleBalčiūnaitė-Murzienė, Gabrielė, and Mindaugas Dzikaras. 2021. "Wheat Germ Agglutinin—From Toxicity to Biomedical Applications" Applied Sciences 11, no. 2: 884. https://doi.org/10.3390/app11020884