
Antioxidant, α-Glucosidase Inhibitory, and Anti-Inflammatory Activities and Cell Toxicity of Waxy and Normal Wheat Sprouts at Various Germination Time


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
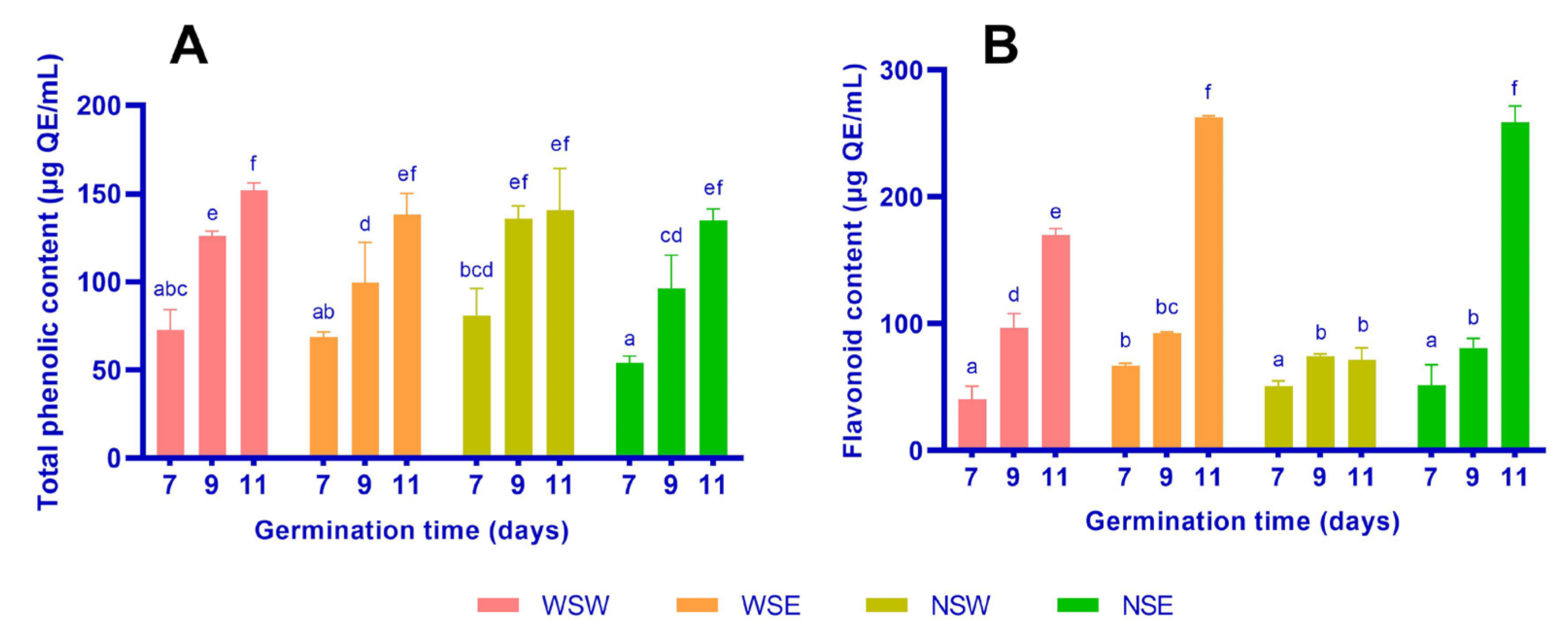
2.3. Total Phenolic Content
2.4. Flavonoid Content
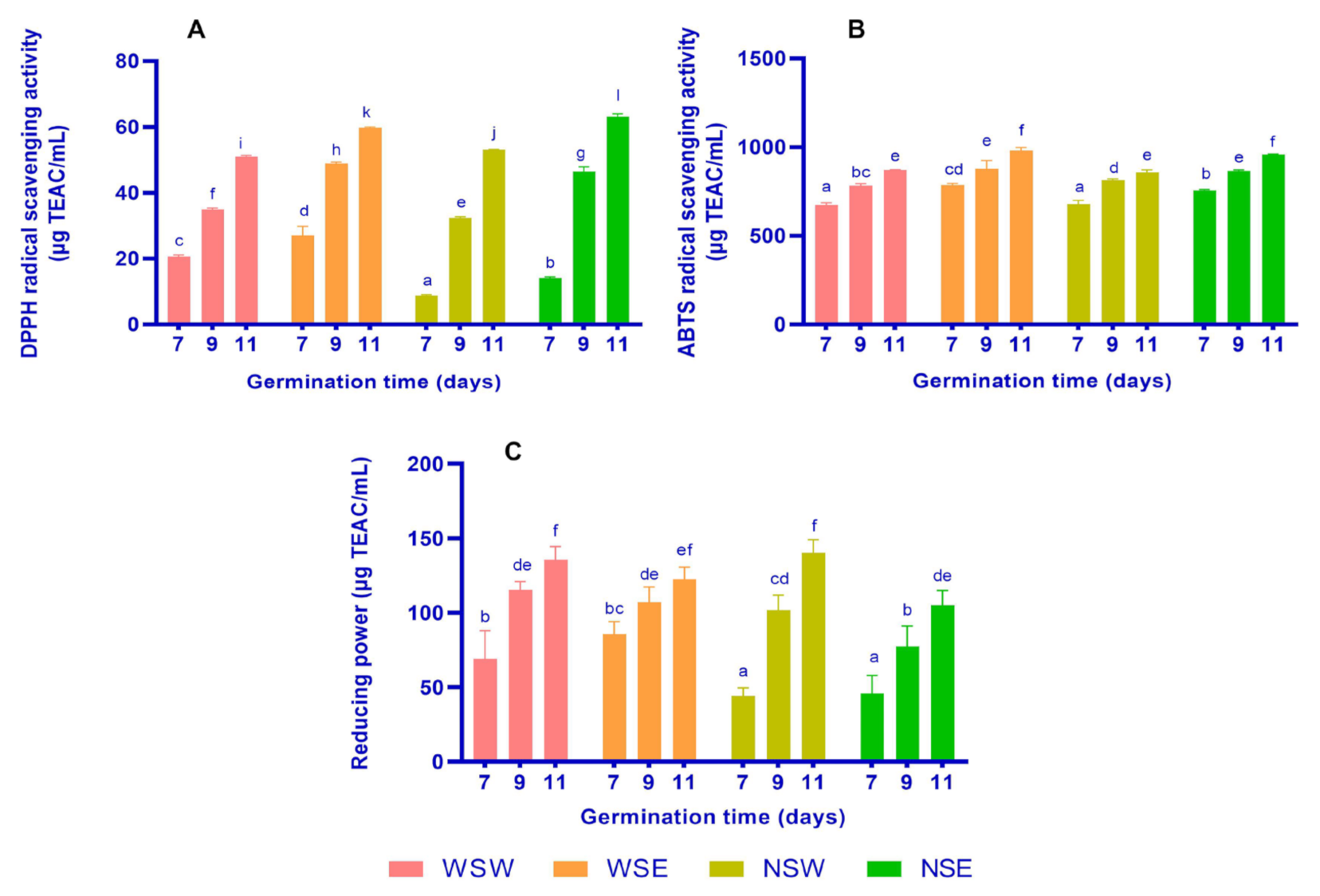
2.5. DPPH Radical Scavenging Activity
2.6. ABTS Radical Scavenging Activity
2.7. Reducing Power
2.8. α-Glucosidase Inhibitory Activity
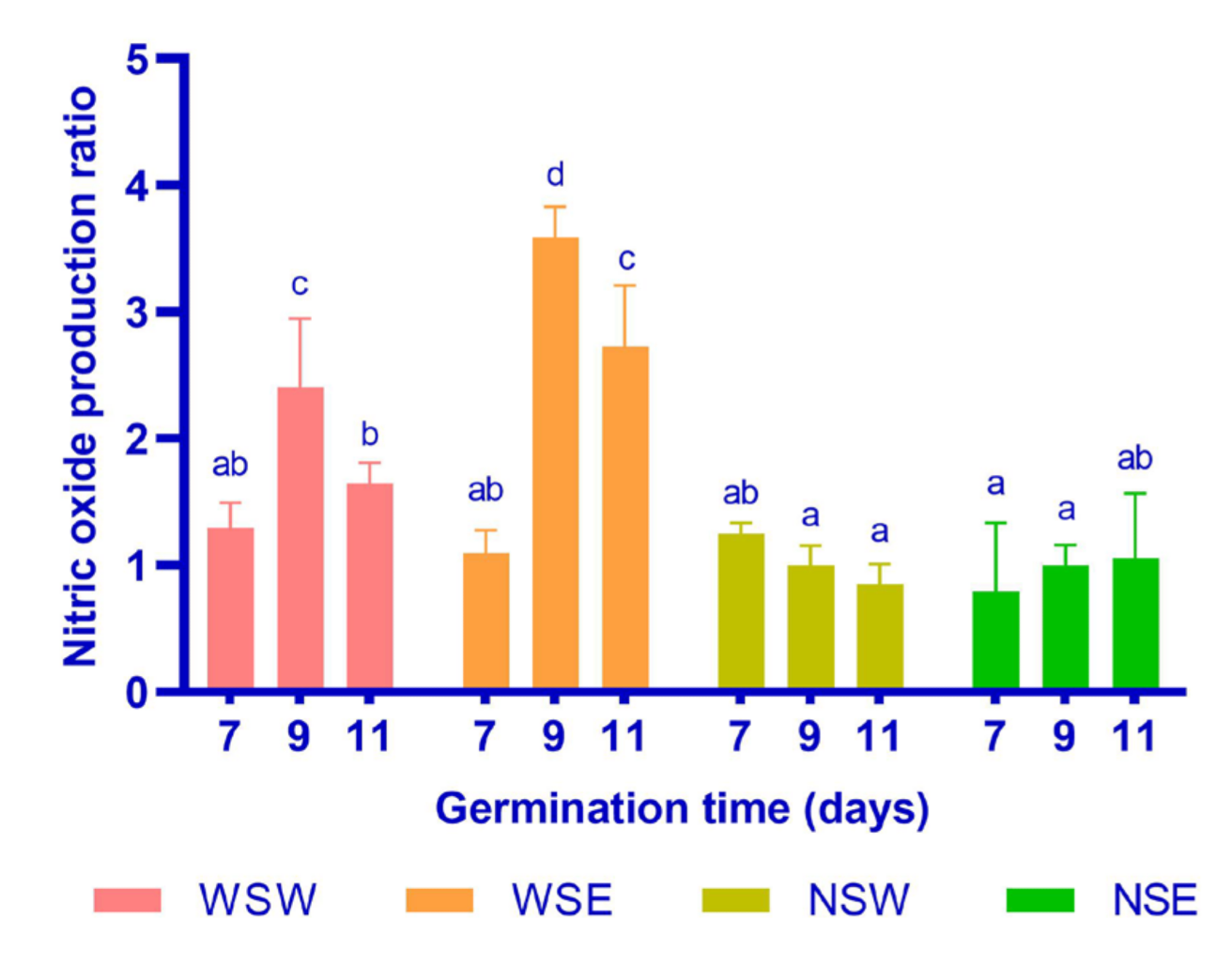
2.9. Nitric Oxide Production Ratio
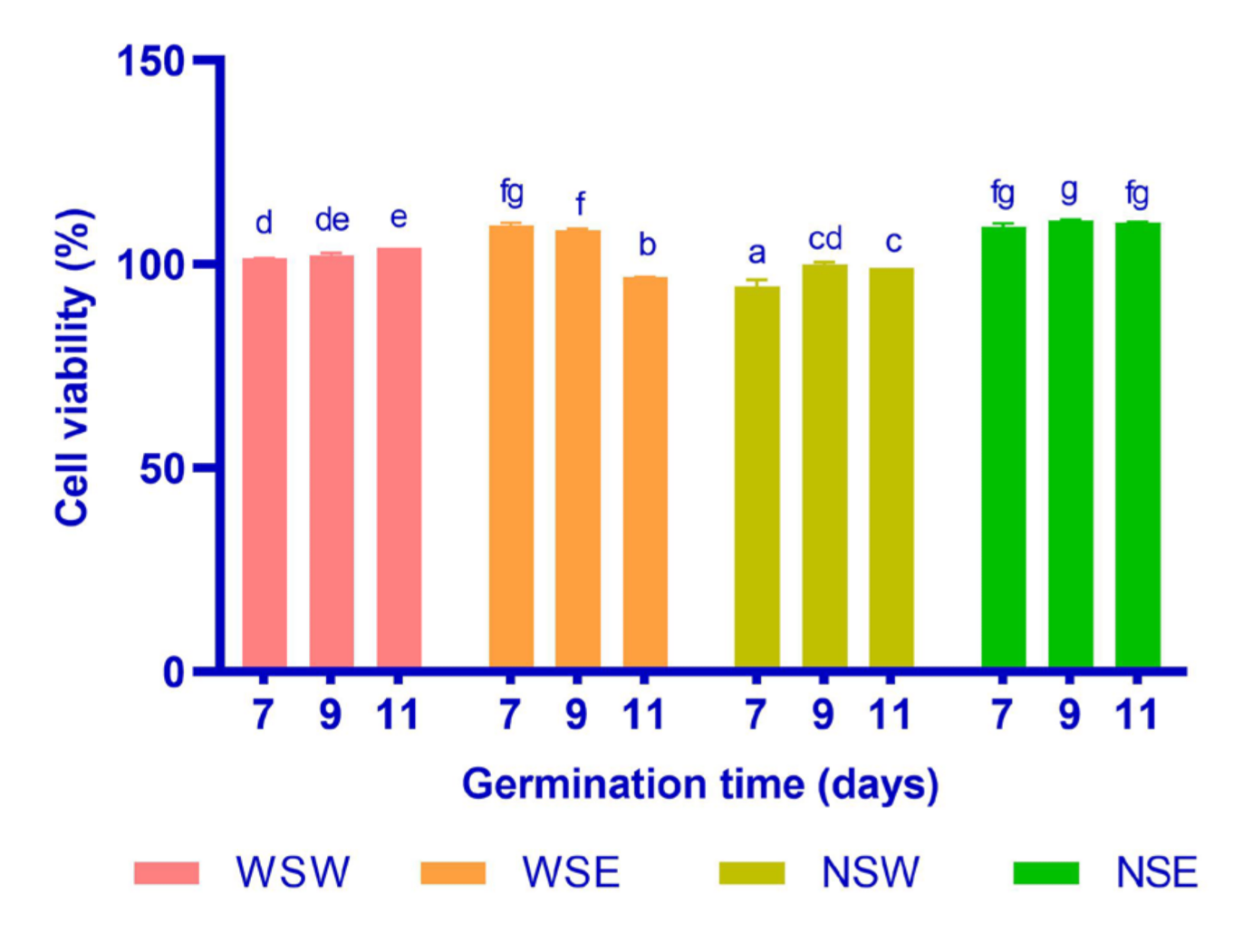
2.10. Cell Viability
2.11. Statistical Analysis
3. Results and Discussion
3.1. Total Phenolic and Flavonoid Contents
3.2. Antioxidant Activities
3.3. α-Glucosidase Inhibitory Activity
3.4. Nitric Oxide Production Ratio
3.5. Cell Viability
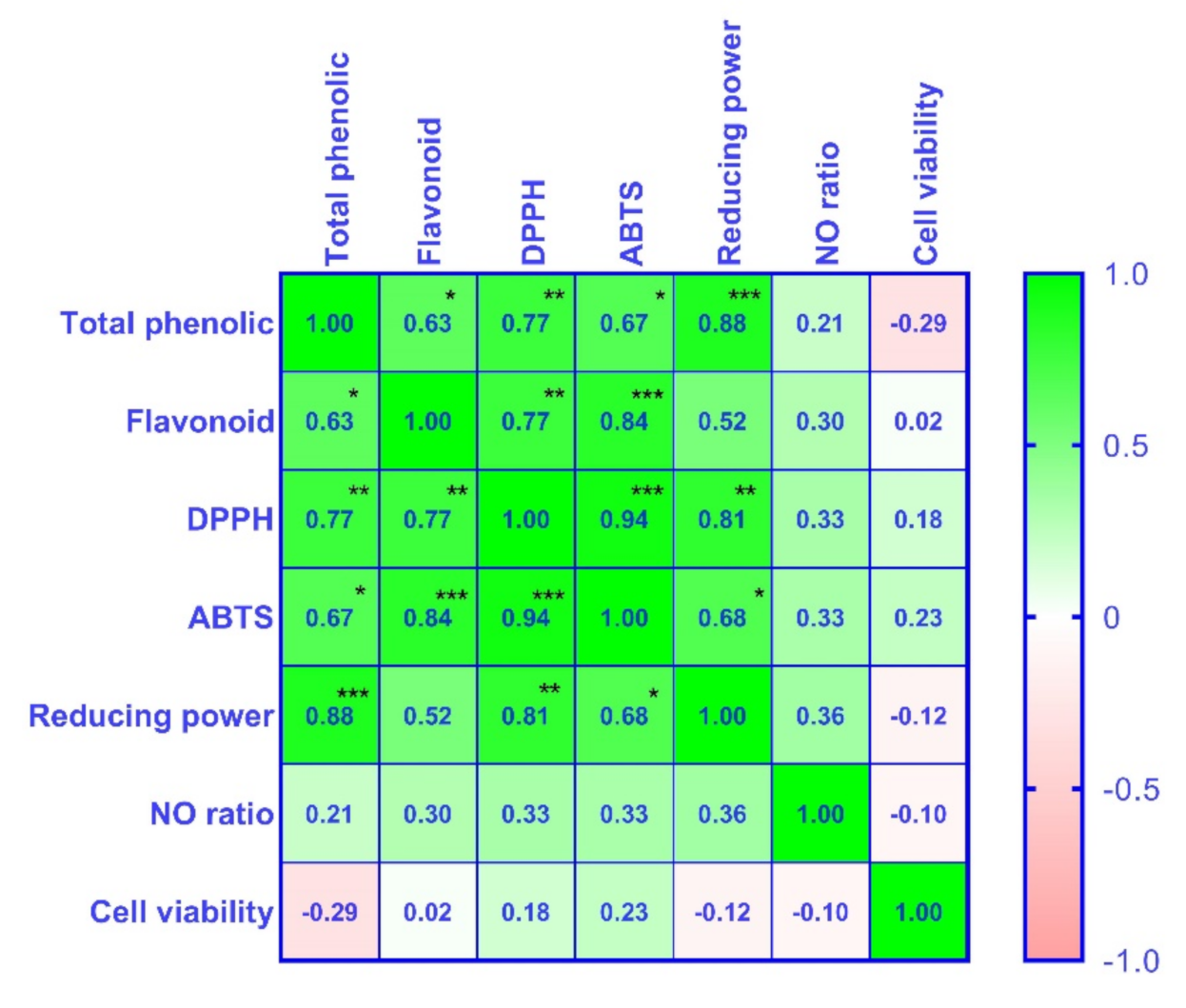
3.6. Pearson’s Correlation Coefficient
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [Green Version]
- Hussein, T.H.A.; Abd El-Shafea, Y.M.; El-Behairy, U.A.A.; Abdallah, M.M.F. Effect of soaking and sprouting using saline water on chemical composition of wheat grains. Arab Univ. J. Agric. Sci. 2019, 27, 707–715. [Google Scholar] [CrossRef]
- Lee, C.K.; Nam, J.H.; Kang, M.S.; Ku, B.C.; Park, K.K.; Kim, J.C.; Son, Y.K.; Park, J.H.; Lee, Y.H.; Son, J.R.; et al. Comparison of physico-chemaical properties between waxy and non-waxy wheat grains. Korean J. Crop. Sci. 2005, 50, 419–427. [Google Scholar]
- Zhang, B.; Zhang, Q.; Wu, H.; Su, C.; Ge, X.; Shen, H.; Han, L.; Yu, X.; Li, W. The influence of repeated versus continuous dry-heating on the performance of wheat starch with different amylose content. LWT 2021, 136, 110380. [Google Scholar] [CrossRef]
- Jaksics, E.; Paszerbovics, B.; Egri, B.; Rakszegi, M.; Tremmel-Bede, K.; Vida, G.; Gergely, S.; Németh, R.; Tömösközi, S. Complex rheological characterization of normal, waxy and high-amylose wheat lines. J. Cereal Sci. 2020, 93, 102982. [Google Scholar] [CrossRef]
- Bai, X.; Yang, S.; Zeng, L.; Han, W.; Ran, X. Study on physicochemical properties of purple waxy wheat starch. Int. J. Food Prop. 2021, 24, 471–481. [Google Scholar] [CrossRef]
- Kiribuchi-Otobe, C.; Fujita, M.; Matsunaka, H.; Sekine, M. Properties of cross-linked starch from waxy mutant wheat Tanikei A6599-4. Cereal Chem. 2006, 83, 590–594. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, W.; Xu, C.; Zhou, X. Studies on the rheological and gelatinization characteristics of waxy wheat flour. Int. J. Biol. Macromol. 2014, 64, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Koehler, P.; Hartmann, G.; Wieser, H.; Rychlik, M. Changes of folates, dietary fiber, and proteins in wheat as affected by germination. J. Agric. Food Chem. 2007, 55, 4678–4683. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Bośko, P.; Živčák, M.; Brestic, M.; Smetanska, I. Bioactive phytochemicals and antioxidant properties of the grains and sprouts of colored wheat genotypes. Molecules 2018, 23, 2282. [Google Scholar] [CrossRef] [Green Version]
- Bonfili, L.; Amici, M.; Cecarini, V.; Cuccioloni, M.; Tacconi, R.; Angeletti, M.; Fioretti, E.; Keller, J.N.; Eleuteri, A.M. Wheat sprout extract-induced apoptosis in human cancer cells by proteasomes modulation. Biochimie 2009, 91, 1131–1144. [Google Scholar] [CrossRef]
- You, S.H.; Pyo, Y.H. Antioxidant anti-inflammatory activities of ethanol extracts from wheat sprout. J. Invest. Cosmetol. 2015, 11, 231–238. [Google Scholar]
- Randhir, R.; Kwon, Y.I.; Shetty, K. Effect of thermal processing on phenolics, antioxidant activity and health-relevant functionality of select grain sprouts and seedlings, Innovative. Food Sci. Emerg. Technol. 2008, 9, 355–364. [Google Scholar] [CrossRef]
- Lee, S.; Lee, Y.; Lee, H.; Kim, D. Anti-oxidative and anti-hyperglycemia effect of Triticum aestivum wheat sprout water extracts on the streptozotocin-induced diabetes mice. Kor. J. Pharmacogn. 2009, 40, 408–414. [Google Scholar]
- Rhee, K.S.; Ziprin, Y.A.; Rhee, K.C. Antioxidant activity of methanolic extracts of various oilseed protein ingredients. J. Food Sci. 1981, 46, 75–77. [Google Scholar] [CrossRef]
- Park, J.S.; Han, I. Effect of extraction solvent on the physiological properties of Korean pear peel (Pyrus pyrifolia cv. Niitaka). Korean J. Food Sci. Technol. 2015, 47, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Fellegrini, N.; Ke, R.; Yang, M.; Rice-Evans, C. Screening of dietary carotenoids and carotenoid-rich fruit extracts for antioxidant activities applying 2,2′-azinobis-3-ethylbenzothiazoline-6-sulfonic acid radical cation decolorization assay. Method Enzymol. 1999, 299, 379–389. [Google Scholar]
- Oyaizu, M. Studies on product of browning reaction: Antioxidative of product of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Yoshimoto, C.; Osajima, K.; Oki, T.; Osajima, Y. In vitro survey of alpha-glucosidase inhibitory food components. Biosci. Biotechnol. Biochem. 1996, 60, 2019–2022. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef] [PubMed]
- Ibidapoa, O.; Henshawb, F.; Shittub, T.; Afolabic, W. Bioactive components of malted millet (Pennisetum glaucum), Soy Residue “okara” and wheat flour and their antioxidant properties. Int. J. Food Prop. 2019, 22, 1886–1898. [Google Scholar] [CrossRef] [Green Version]
- Donkor, O.N.; Stojanovsk, L.; Ginn, P.; Ashton, J.; Vasiljevic, T. Germinated grains-sources of bioactive compounds. Food Chem. 2012, 135, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Dziki, D.; Gawlik-Dziki, U.; Kordowska-Wiater, M.; Doma-Pytka, M. Influence of elicitation and germination conditions on biological activity of wheat sprouts. J. Chem. 2015, 649709. [Google Scholar] [CrossRef] [Green Version]
- Benincasa, P.; Galieni, A.; Manetta, A.C.; Pace, R.; Guiducci, M.; Pisante, M.; Stagnari, F. Phenolic compounds in grains, sprouts and wheatgrass of hulled and non-hulled wheat species. J. Sci. Food Agric. 2015, 95, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Durairaj, V.; Hoda, M.; Shakya, G.; Babu, S.P.P.; Rajagopalan, R. Phytochemical screening and analysis of antioxidant properties of aqueous extract of wheatgrass. Asian Pac. J. Trop. Med. 2014, 7, 398–404. [Google Scholar] [CrossRef]
- Kaur, H.; Gill, B.S. Changes in physicochemical, nutritional characteristics and ATR–FTIR molecular interactions of cereal grains during germination. J. Food Sci. Technol. 2020, 58, 2313–2324. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa, buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Tarasevičienė, Ž.; Viršilė, A.; Danilčenko, H.; Duchovskis, P.; Paulauskienė, A.; Gajewski, M. Effects of germination time on the antioxidant properties of edible seeds. CyTA J. Food 2019, 17, 447–454. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, S.H.; Hwang, I.G.; Woo, K.S.; Kim, K.J.; Lee, M.J.; Kim, D.J.; Kim, T.J.; Lee, J.; Jeong, H.S. Antioxidant and antiproliferation activities of winter cereal crops before and after germination. Food Sci. Biotechnol. 2013, 22, 181–186. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, B.G.; Han, I. Effect of drying and extraction methods on antioxidant activity of Gnaphalium affine D. DON. J. Korean Soc. Food Sci. Nutr. 2015, 44, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Vocadlo, D.J.; Davies, G.J. Mechanistic insights into glycosidase chemistry. Curr. Opin. Chem. Biol. 2008, 12, 539–555. [Google Scholar] [CrossRef]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Natural products as α-amylase and α-glucosidase inhibitors and their hypoglycaemic potential in the treatment of diabetes: An update. Mini Rev. Med. Chem. 2010, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Feng, D.; Wang, T.; Ren, Y.; Liu, Y.; Wang, J. Inhibitors of α-amylase and α-glucosidase: Potential linkage for whole cereal foods on prevention of hyperglycemia. Food Sci. Nutr. 2020, 8, 6320–6337. [Google Scholar] [CrossRef] [PubMed]
- Abbs, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Green, S.J.; Scheller, L.F.; Marietta, M.A.; Seguin, M.C.; Klotz, F.W.; Slayter, M.; Nelson, B.J.; Nacy, C.A. Nitric oxide: Cytokine-regulation of nitric oxide in host resistance to intracellular pathogens. Immunol. Lett. 1994, 43, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Majumder, K.; Mine, Y.; Wu, J. The potential of food protein-derived antiinflammatory peptides against various chronic inflammatory diseases. J. Sci. Food Agric. 2016, 96, 2303–2311. [Google Scholar] [CrossRef]
- Sandoval-Sicairos, E.S.; Milán-Noris, A.K.; Luna-Vital, D.A.; Milán-Carrillo, J.; Montoya-Rodríguez, A. Anti-inflammatory and antioxidant effects of peptides released from germinated amaranth during in vitro simulated gastrointestinal digestion. Food Chem. 2021, 343, 128394. [Google Scholar] [CrossRef]
- Tomé-Sánchez, I.; Martín-Diana, A.B.; Peñas, E.; Bautista-Expósito, S.; Frias, J.; Rico, D.; González-Maillo, L.; Martinez-Villaluenga, C. Soluble phenolic composition tailored by germination conditions accompany antioxidant and anti-inflammatory properties of wheat. Antioxidants 2020, 9, 426. [Google Scholar] [CrossRef]
- Whent, M.; Huang, H.; Xie, Z.; Lutterodt, H.; Yu, L.; Fuerst, E.P.; Morris, C.F.; Yu, L.; Luthria, D. Phytochemical composition, anti-inflammatory, and antiproliferative activity of whole wheat flour. J. Agric. Food Chem. 2012, 60, 2129–2135. [Google Scholar] [CrossRef]
- Pomothy, J.M.; Pászti-Gere, E.; Barna, R.F.; Prokoly, D.; Jerzsele, Á. The impact of fermented wheat germ extract on porcine epithelial cell line exposed to deoxynivalenol and T2 mycotoxins. Oxidative Med. Cell. Longev. 2020, 3854247. [Google Scholar] [CrossRef]
- Tillmann, M.T.; de Mello Mendes, C.B.; Fischer, G.; Júnior, S.A.V.; Fernandes, C.G.; de Oliveria, N.M. Triticum aestivum in open skin wounds: Cytotoxicity and collagen histopathology. Vet. Med. 2018, 39, 1547–1554. [Google Scholar] [CrossRef]
- Lim, W.; Park, S.; Bazer, F.W.; Song, G. Apigenin reduces survival of choriocarcinoma cells by inducing apoptosis via the PI3K/AKT and ERK1/2 MAPK pathways. J. Cell. Physiol. 2016, 231, 2690–2699. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Encinas, M.A.; Carvajal-Millan, E.; Rascón-Chu, A.; Astiazarán-García, H.; Valencia-Rivera, D.E.; Brown-Bojorquez, F.; Alday, E.; Velazquez, C. Arabinoxylan-based particles: In vitro antioxidant capacity and cytotoxicity on a human colon cell line. Medicina 2019, 55, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogala-Kałucka, M.; Kawka, A.; Dwiecki, K.; Siger, A. Evaluation of bioactive compound in cereals study of wheat, barley, oat and selected grain products. Acta Sci. Pol. Technol. Aliment. 2020, 19, 405–423. [Google Scholar] [PubMed]
- Ali, L.; Khan, S.; Nazir, M.; Raiz, N.; Naz, S.; Zengin, G.; Mukhtar, M.; Parveen, S.; Shazmeen, N.; Saleem, M.; et al. Chemical profiling, in vitro biological activities and Pearson correlation between phenolic contents and antioxidant activities of Caragana brachyantha Rech.f. S. Afr. J. Bot. 2021, 140, 189–193. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Time | Glucosidase Inhibitory Activity (%) 1 | |
|---|---|---|
| Waxy Wheat Sprout | Normal Wheat Sprout | |
| 7 days | 84.61 ± 6.14 a,2 | 90.85 ± 0.24 b |
| 9 days 11 days | 90.92 ± 0.38 b 94.19 ± 0.40 b | 93.58 ± 0.85 b 94.47 ± 0.49 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, I. Antioxidant, α-Glucosidase Inhibitory, and Anti-Inflammatory Activities and Cell Toxicity of Waxy and Normal Wheat Sprouts at Various Germination Time. Appl. Sci. 2021, 11, 7755. https://doi.org/10.3390/app11167755
Han I. Antioxidant, α-Glucosidase Inhibitory, and Anti-Inflammatory Activities and Cell Toxicity of Waxy and Normal Wheat Sprouts at Various Germination Time. Applied Sciences. 2021; 11(16):7755. https://doi.org/10.3390/app11167755
Chicago/Turabian StyleHan, Inhwa. 2021. "Antioxidant, α-Glucosidase Inhibitory, and Anti-Inflammatory Activities and Cell Toxicity of Waxy and Normal Wheat Sprouts at Various Germination Time" Applied Sciences 11, no. 16: 7755. https://doi.org/10.3390/app11167755