
A Redox-Neutral, Two-Enzyme Cascade for the Production of Malate and Gluconate from Pyruvate and Glucose



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Enzyme Production
2.2. Enzyme Activity Assays
2.3. Determination of Kinetic Constants
2.4. HPIC Analysis
3. Results
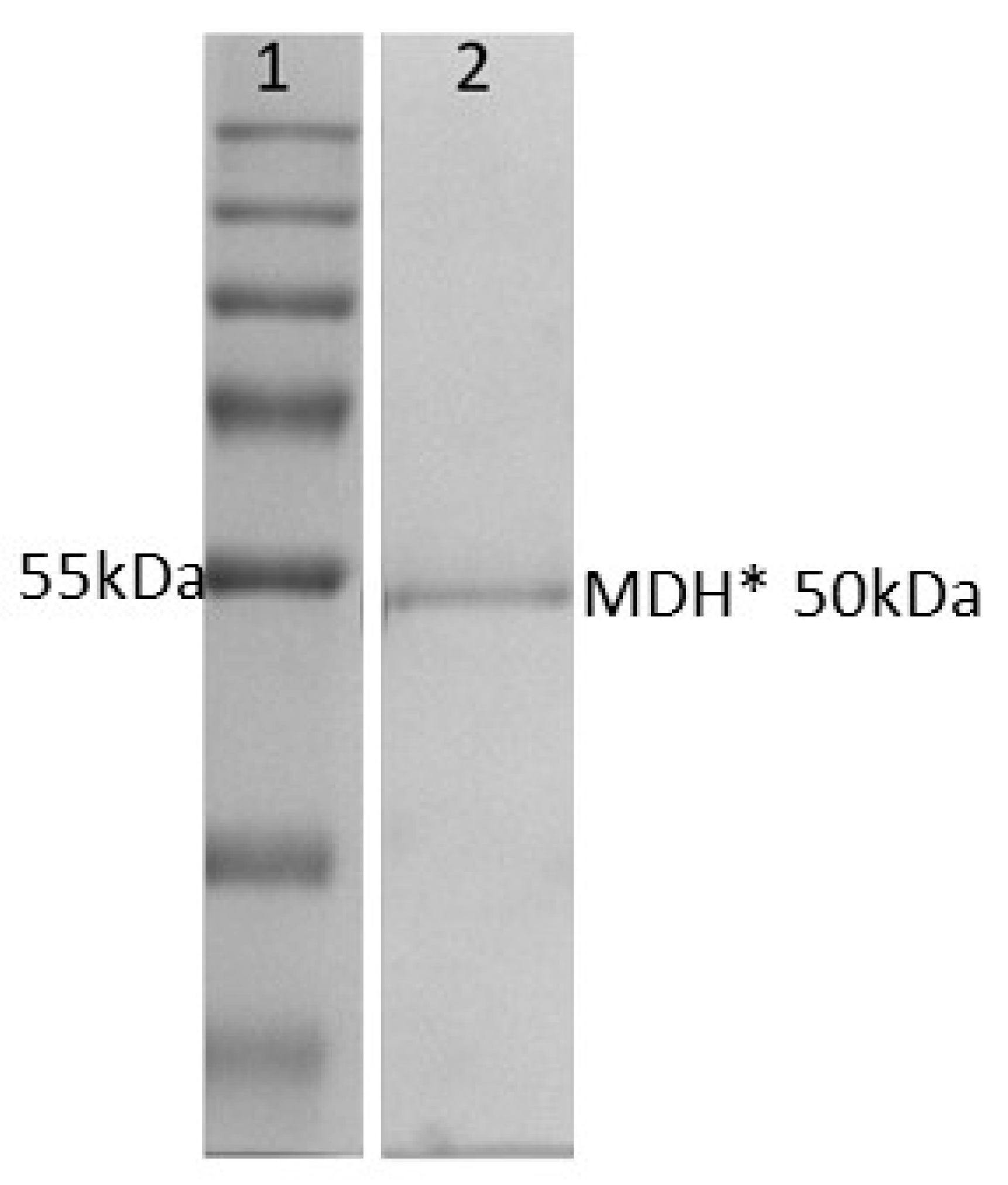
3.1. Purity and Activity of Recombinant MDH*
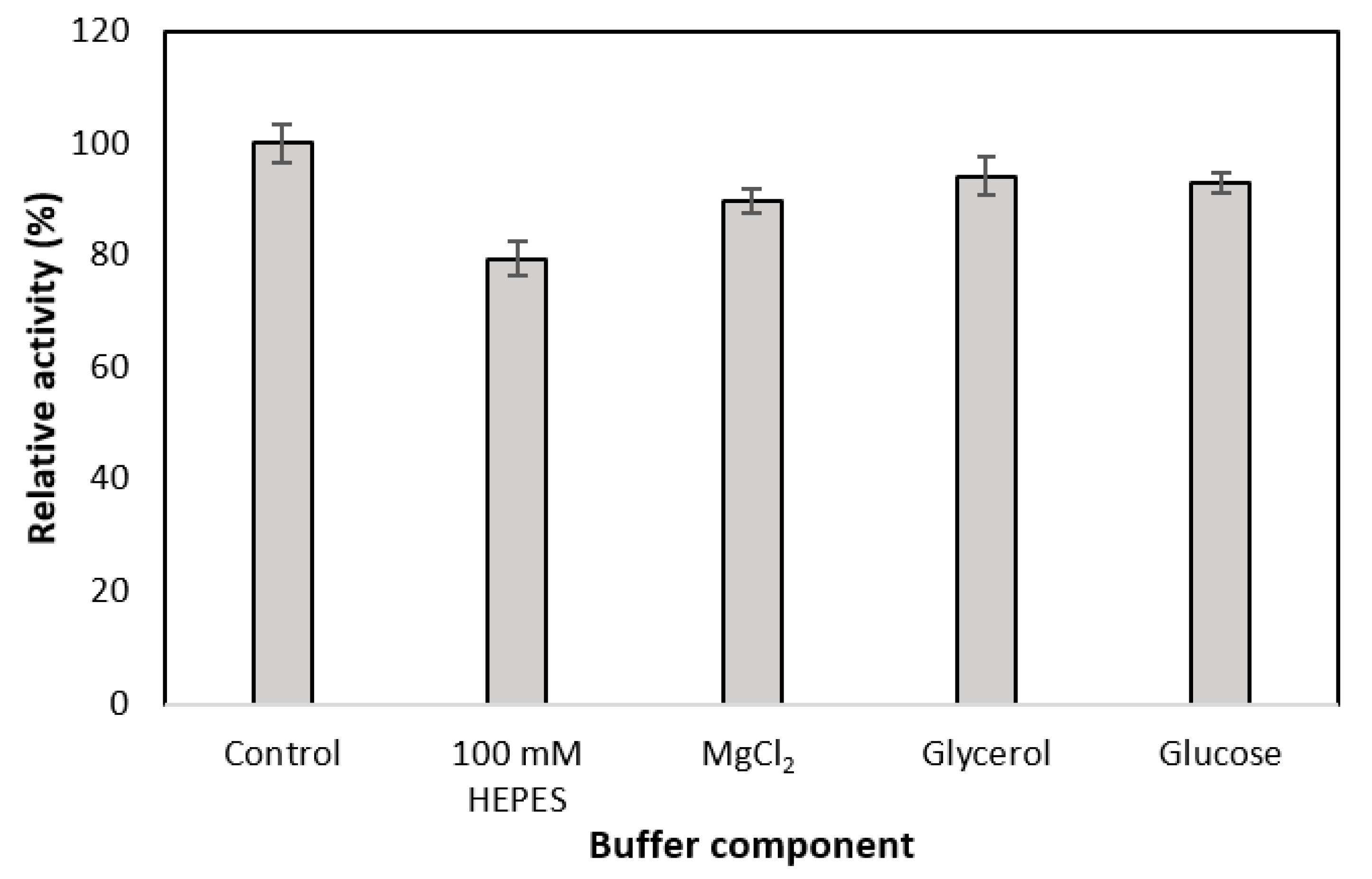
3.2. Effect of GDH Buffer Components on the Activity of MDH*
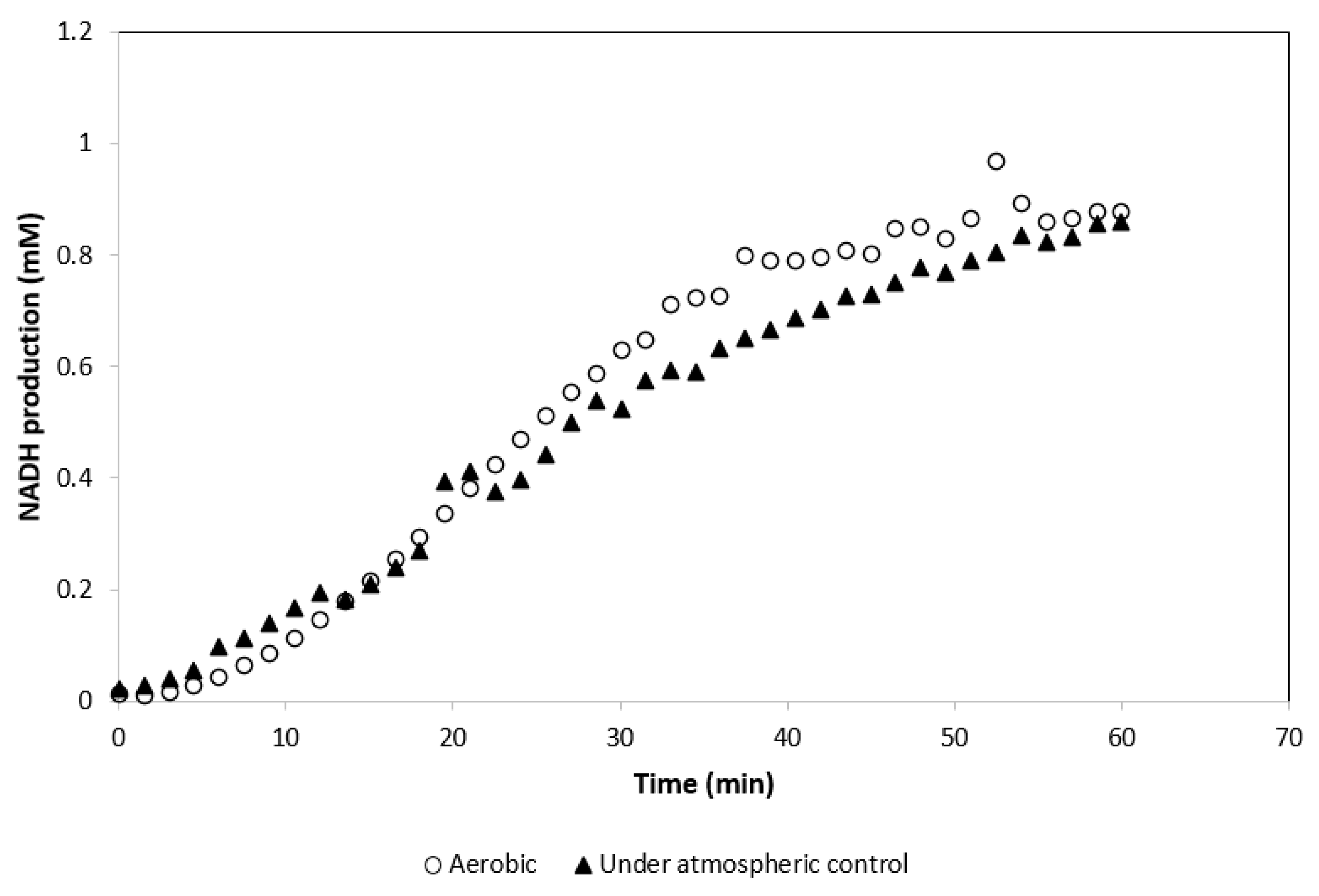
3.3. Effect of Atmospheric Conditions Required by MDH* on the Activity of GDH
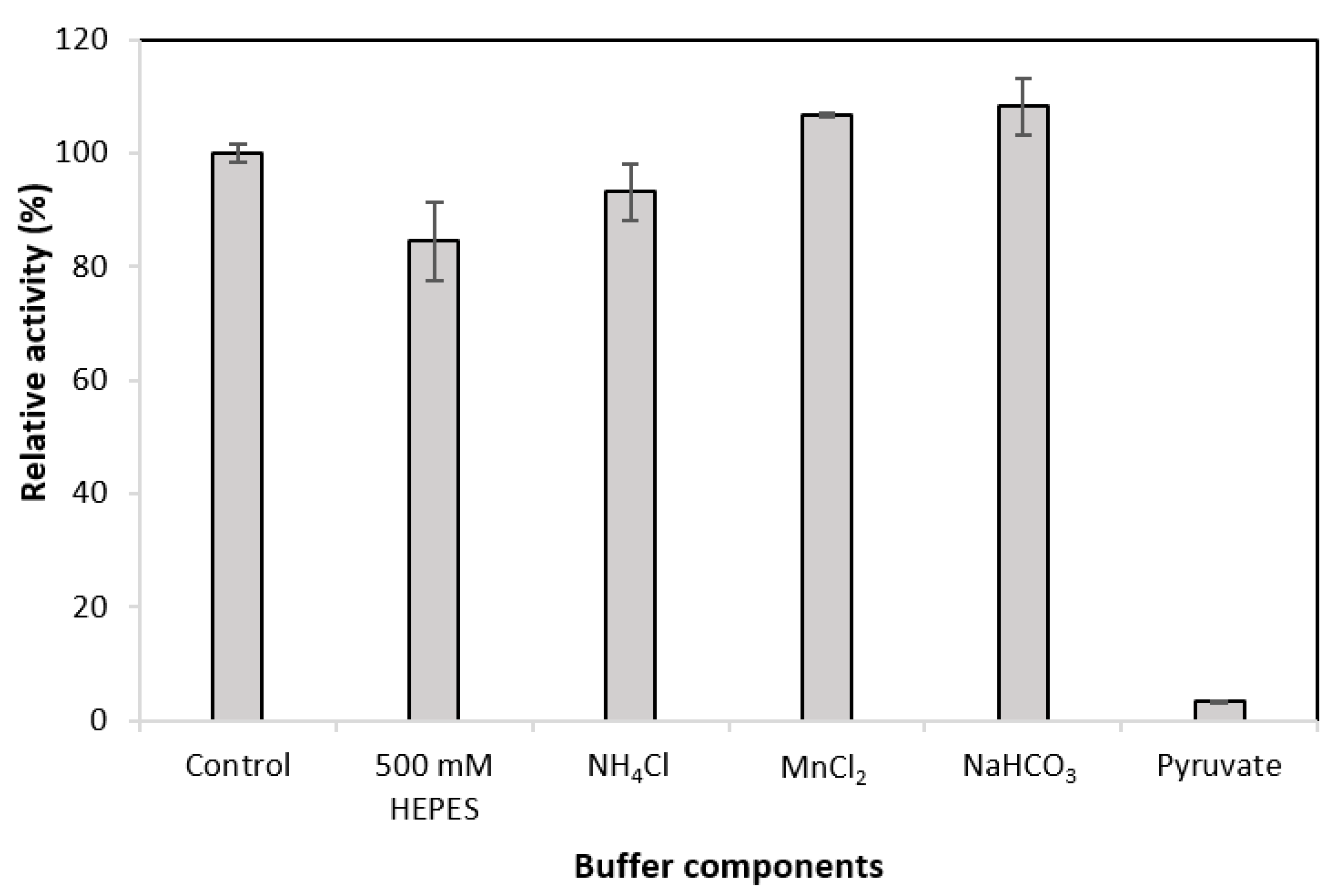
3.4. Effect of MDH* Assay Buffer Components on the Activity of GDH
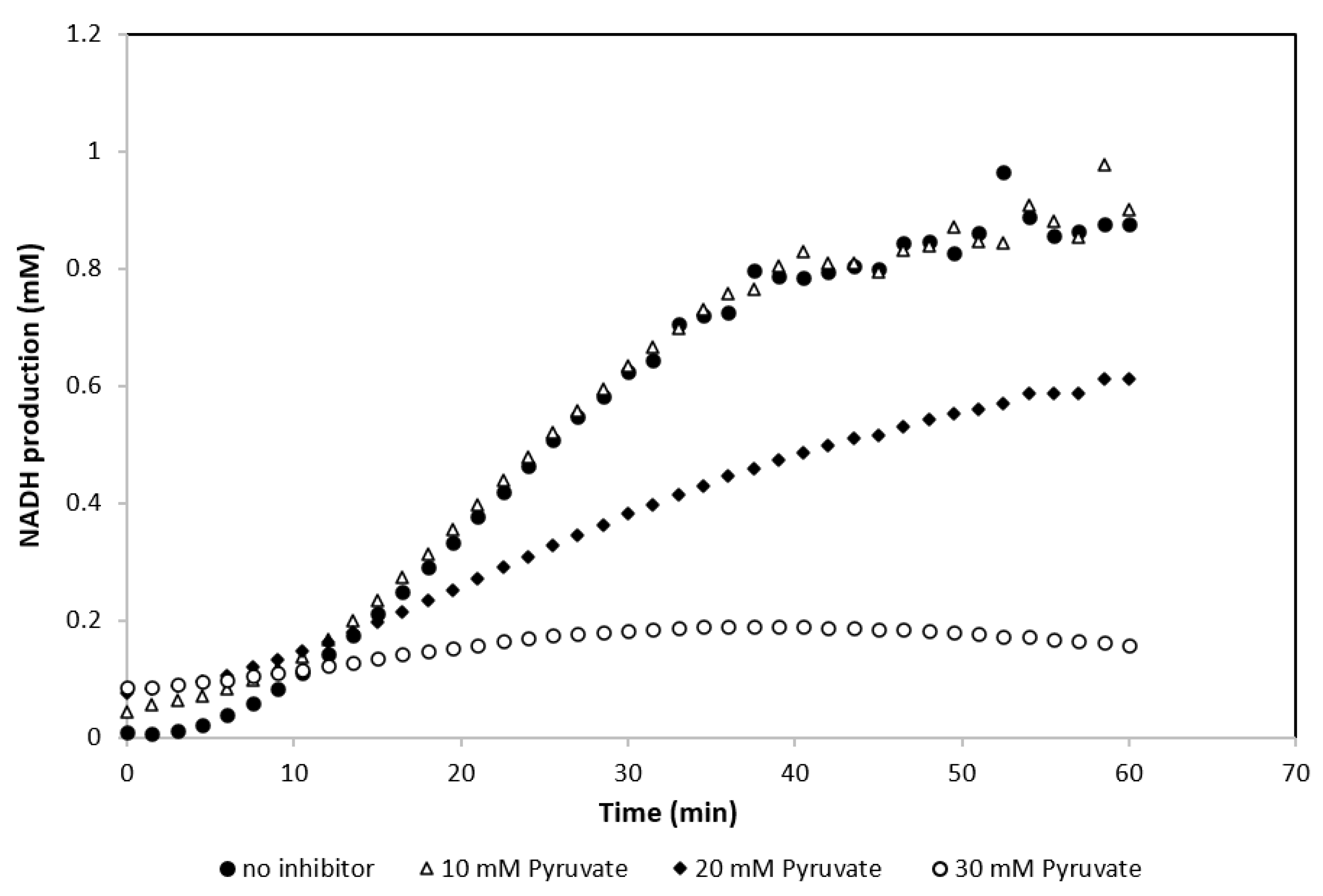
3.5. Effect of Pyruvate Concentration on the Activity of GDH
3.6. Optimization of Atmospheric and Temperature Conditions for the Activity of Purified MDH*
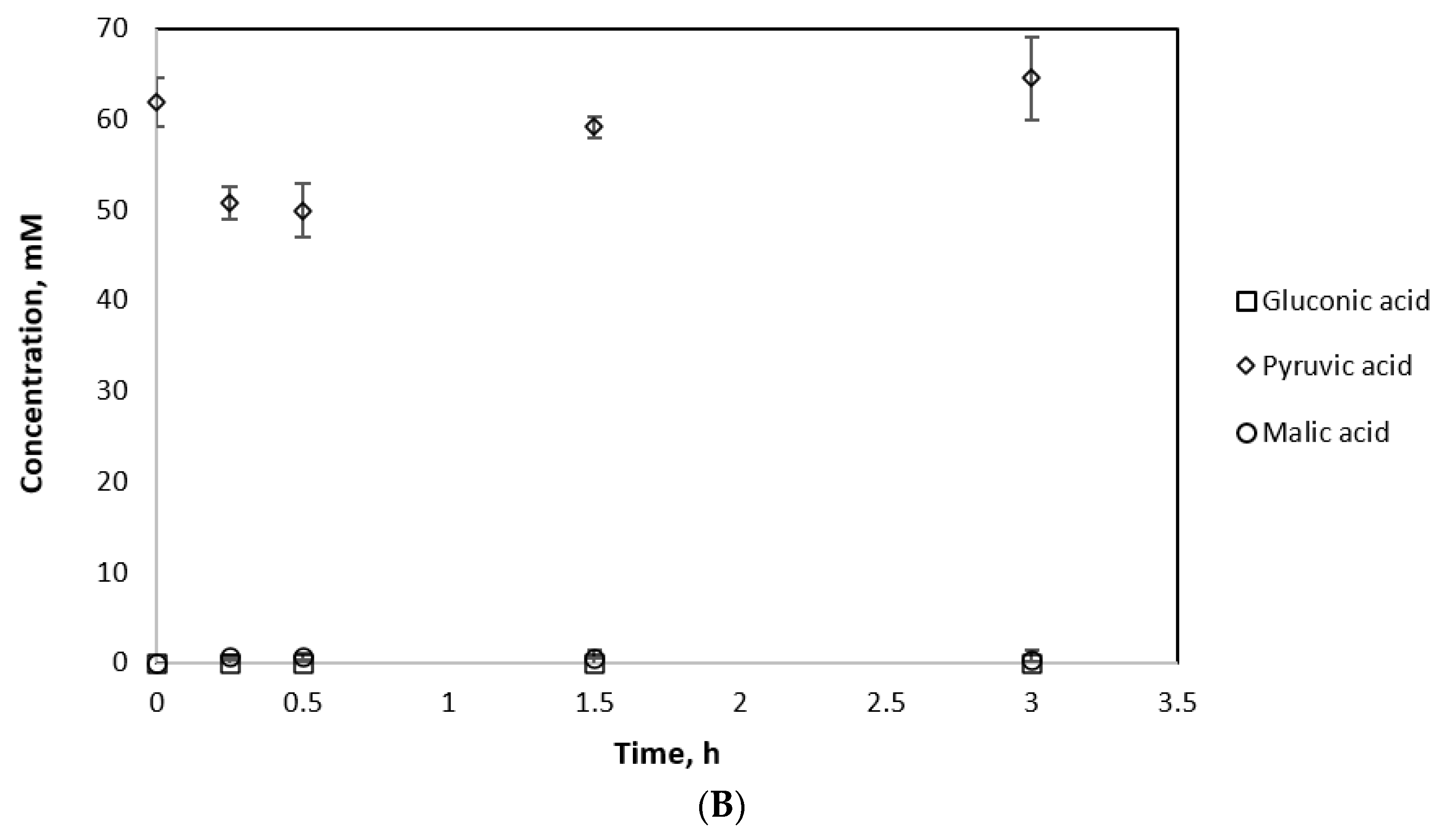
3.7. Coupling MDH* and GDH for Cofactor Recycling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Jang, Y.-S.; Kim, B.; Shin, J.H.; Choi, Y.J.; Choi, S.; Song, C.W.; Lee, J.; Park, H.G.; Lee, S.Y. Bio-based production of C2–C6 platform chemicals. Biotechnol. Bioeng. 2012, 109, 2437–2459. [Google Scholar] [CrossRef] [PubMed]
- Erickson, B.; Nelson, J.E.; Winters, P. Perspective on opportunities in industrial biotechnology in renewable chemicals. Biotechnol. J. 2012, 7, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Dudley, Q.M.; Karim, A.S.; Jewett, M.C. Cell-free metabolic engineering: Biomanufacturing beyond the cell. Biotechnol. J. 2015, 10, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Singhvi, M.; Gokhale, D. Biomass to biodegradable polymer (PLA). RSC Adv. 2013, 3, 13558–13568. [Google Scholar] [CrossRef]
- Guterl, J.-K.; Sieber, V. Biosynthesis “debugged”: Novel bioproduction strategies. Eng. Life Sci. 2013, 13, 4–18. [Google Scholar] [CrossRef]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—the US Department of Energy’s “Top 10” revisited. Green Chem. 2010, 12, 539–554. [Google Scholar] [CrossRef]
- Wang, X.; Saba, T.; Yiu, H.H.P.; Howe, R.F.; Anderson, J.A.; Shi, J. Cofactor NAD(P)H regeneration inspired by heterogeneous pathways. Chem 2017, 2, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Rollin, J.A.; Tam, T.K.; Zhang, Y.H.P. New biotechnology paradigm: Cell-free biosystems for biomanufacturing. Green Chem. 2013, 15, 1708–1719. [Google Scholar] [CrossRef]
- Guterl, J.K.; Garbe, D.; Carsten, J.; Steffler, F.; Sommer, B.; Reiße, S.; Philipp, A.; Haack, M.; Rühmann, B.; Koltermann, A.; et al. Cell-free metabolic engineering: Production of chemicals by minimized reaction cascades. ChemSusChem 2012, 5, 2165–2172. [Google Scholar] [CrossRef]
- Petroll, K.; Kopp, D.; Care, A.; Bergquist, P.L.; Sunna, A. Tools and strategies for constructing cell-free enzyme pathways. Biotechnol. Adv. 2019, 37, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Chenault, H.K.; Simon, E.S.; Whitesides, G.M. Cofactor regeneration for enzyme-catalysed synthesis. Biotechnol. Genet. Eng. Rev. 1988, 6, 221–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, F.; Arends, I.W.C.E.; Holtmann, D. Enzymatic reductions for the chemist. Green Chem. 2011, 13, 2285–2314. [Google Scholar] [CrossRef]
- Morimoto, Y.; Honda, K.; Ye, X.; Okano, K.; Ohtake, H. Directed evolution of thermotolerant malic enzyme for improved malate production. J. Biosci. Bioeng. 2014, 117, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagoz, P.; Mandair, R.; Manayil, J.C.; Lad, J.; Chong, K.; Kyriakou, G.; Lee, A.F.; Wilson, K.; Bill, R.M. Purification and immobilization of engineered glucose dehydrogenase: A new approach to producing gluconic acid from breadwaste. Biotechnol. Biofuels 2020, 13, 100. [Google Scholar] [CrossRef]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Ryan, B.; Henehan, G.T.M. β-Glucosidase from streptomyces griseus: Nanoparticle immobilisation and application to alkyl glucoside synthesis. Protein Expr. Purif. 2017, 132, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Misson, M.; Zhang, H.; Jin, B. Nanobiocatalyst advancements and bioprocessing applications. J. R. Soc. Interface 2015, 12, 20140891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GDH Standard Assay Conditions at 55 °C | MDH* Standard Assay Conditions at 55 °C |
|---|---|
| No atmospheric control required 100 mM HEPES pH 7.5 2.5% glycerol 30 mM MgCl2 100 mM glucose 5 mM NAD+ | 15% CO2 and 5% O2 500 mM HEPES pH 7.5 5 mM NH4Cl 0.5 mM MnCl2 30 mM pyruvate 85 mM NaHCO3 1 mM NADH |
| Experiment Number | O2 (%) | CO2 (%) | Temperature (°C) | Initial Rate (mM/min) | Initial Rate * (mM/min) |
|---|---|---|---|---|---|
| 1 | 2.5 | 5 | 55 | 0.068 | 0.066 |
| 2 | 7.5 | 5 | 55 | 0.061 | 0.064 |
| 3 | 2.5 | 15 | 55 | 0.067 | 0.065 |
| 4 | 7.5 | 15 | 55 | 0.086 | 0.088 |
| 5 | 2.5 | 10 | 45 | 0.030 | 0.039 |
| 6 | 7.5 | 10 | 45 | 0.059 | 0.063 |
| 7 | 2.5 | 10 | 65 | 0.098 | 0.095 |
| 8 | 7.5 | 10 | 65 | 0.103 | 0.094 |
| 9 | 5 | 5 | 45 | 0.052 | 0.045 |
| 10 | 5 | 15 | 45 | 0.054 | 0.082 |
| 11 | 5 | 5 | 65 | 0.076 | 0.049 |
| 12 | 5 | 15 | 65 | 0.092 | 0.099 |
| 13 | 5 | 10 | 55 | 0.078 | 0.086 |
| 14 | 5 | 10 | 55 | 0.092 | 0.086 |
| 15 | 5 | 10 | 55 | 0.088 | 0.086 |
| Scenario 1: Presence of Both Enzymes (GDH and MDH*), NADH as Cofactor | Scenario 2: Presence of Both Enzymes (GDH and MDH*), NAD+ as Cofactor | Control: No Enzyme, Contains Same Amount of NADH as Scenario 1 | |
|---|---|---|---|
| Final NADH concentration, mM | 1.21 ± 0.06 | 1.31 ± 0.01 | 1.74 ± 0.04 |
| Gluconic acid concentration, mM | 8.81 | 10.45 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandair, R.; Karagoz, P.; Bill, R.M. A Redox-Neutral, Two-Enzyme Cascade for the Production of Malate and Gluconate from Pyruvate and Glucose. Appl. Sci. 2021, 11, 4877. https://doi.org/10.3390/app11114877
Mandair R, Karagoz P, Bill RM. A Redox-Neutral, Two-Enzyme Cascade for the Production of Malate and Gluconate from Pyruvate and Glucose. Applied Sciences. 2021; 11(11):4877. https://doi.org/10.3390/app11114877
Chicago/Turabian StyleMandair, Ravneet, Pinar Karagoz, and Roslyn M. Bill. 2021. "A Redox-Neutral, Two-Enzyme Cascade for the Production of Malate and Gluconate from Pyruvate and Glucose" Applied Sciences 11, no. 11: 4877. https://doi.org/10.3390/app11114877