High-Yielding Lovastatin Producer Aspergillus terreus Shows Increased Resistance to Inhibitors of Polyamine Biosynthesis



Abstract
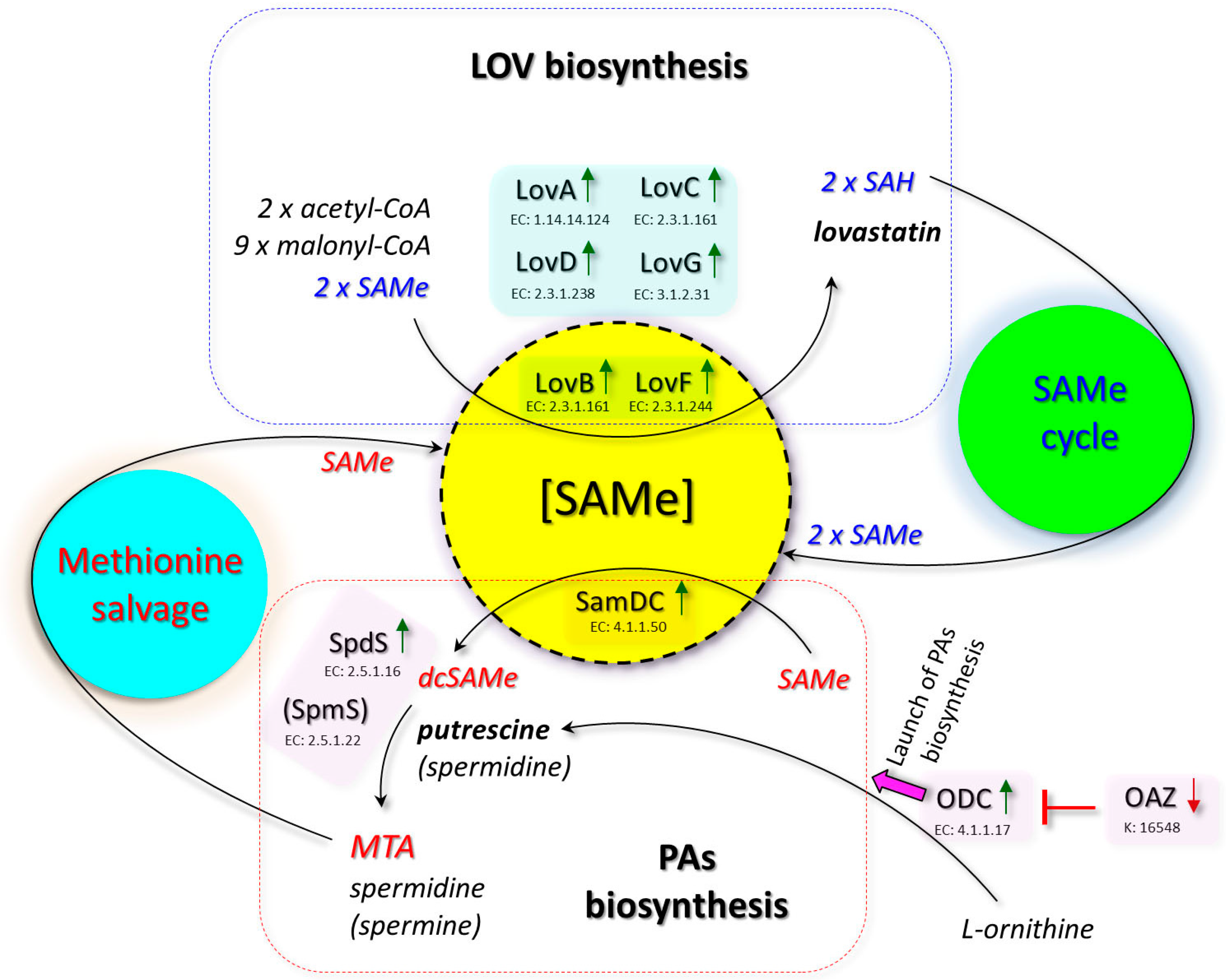
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganism Strains Used in the Work
2.3. Cultivation of A. terreus Strains
2.4. Quantification of Fungal Growth Inhibition
2.5. Preparation of Total RNA and cDNA Synthesis
2.6. qPCR Analysis
3. Results
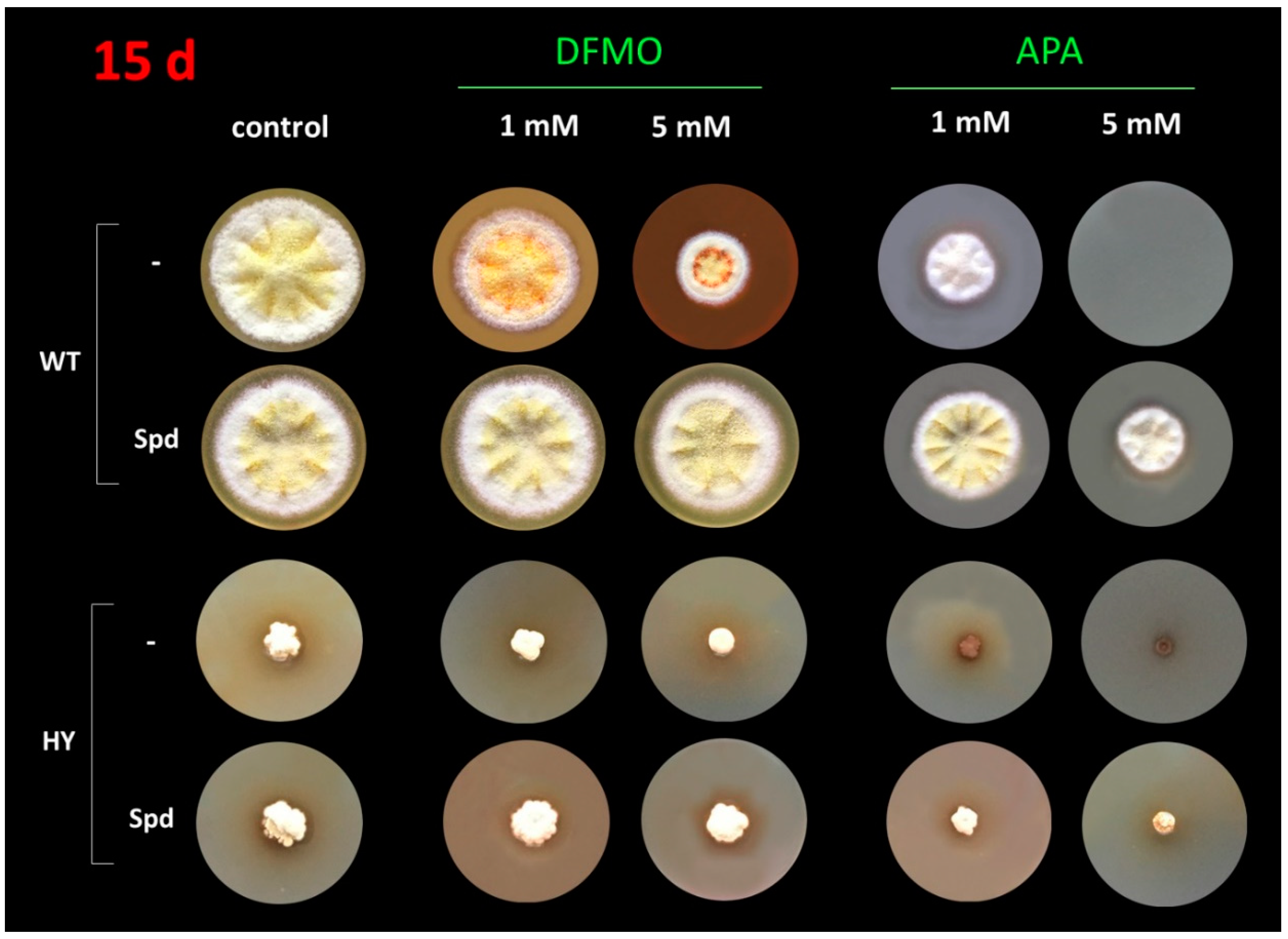
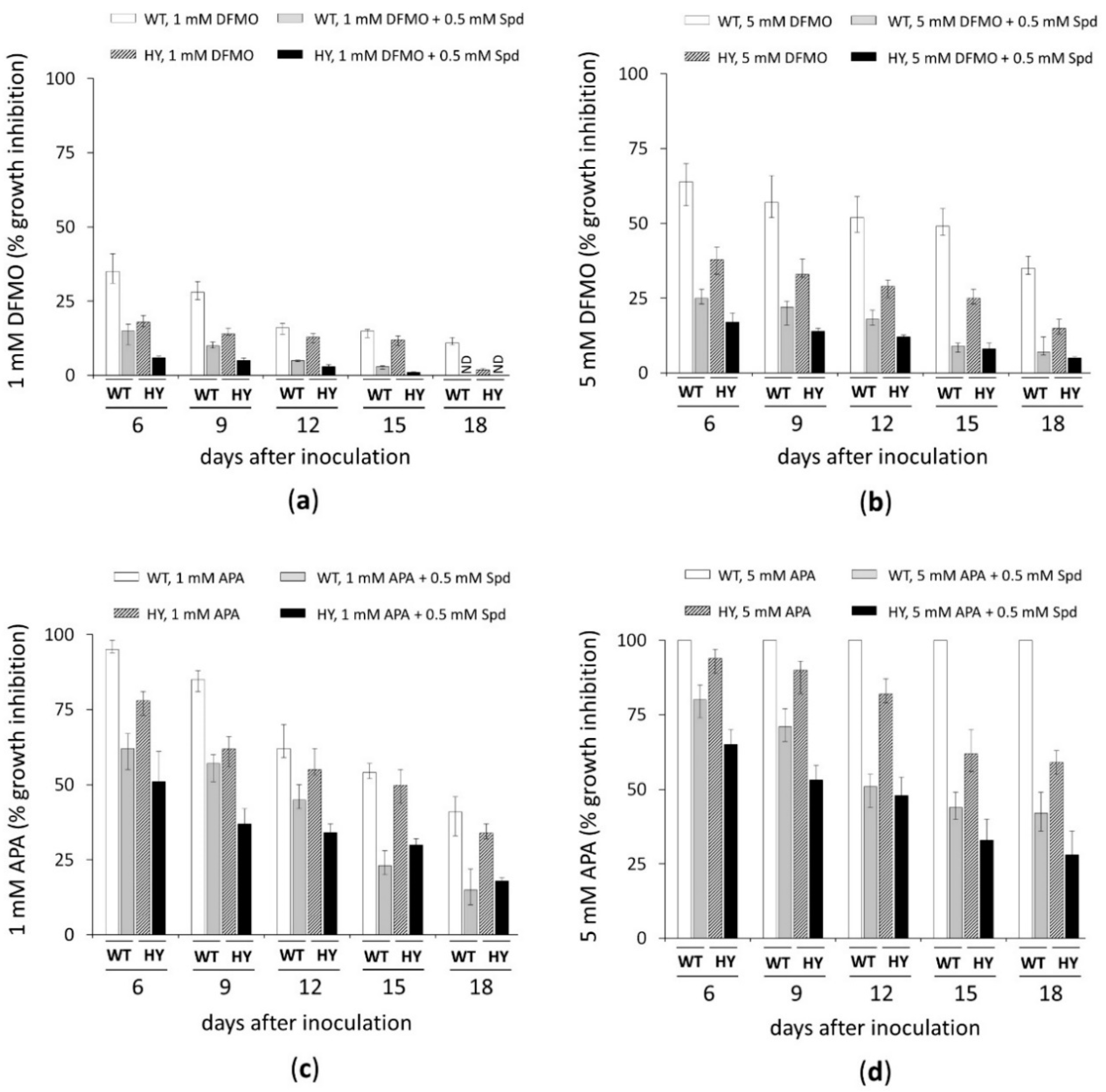
3.1. Inhibitory Analysis of A. terreus Cell Growth by DFMO and APA
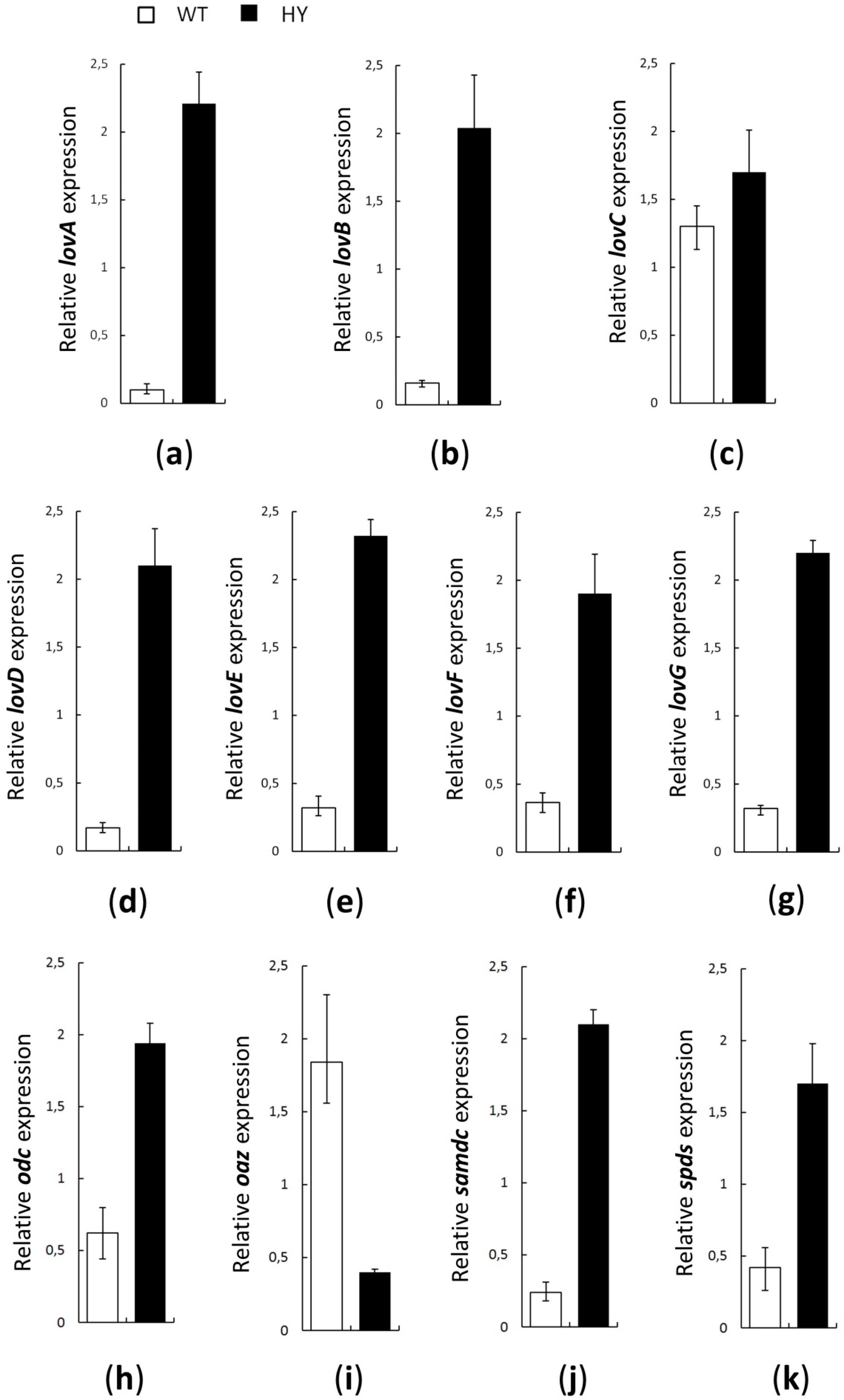
3.2. Expression Levels of PAs Metabolism Genes and lovB in A. terreus WT and HY Strains
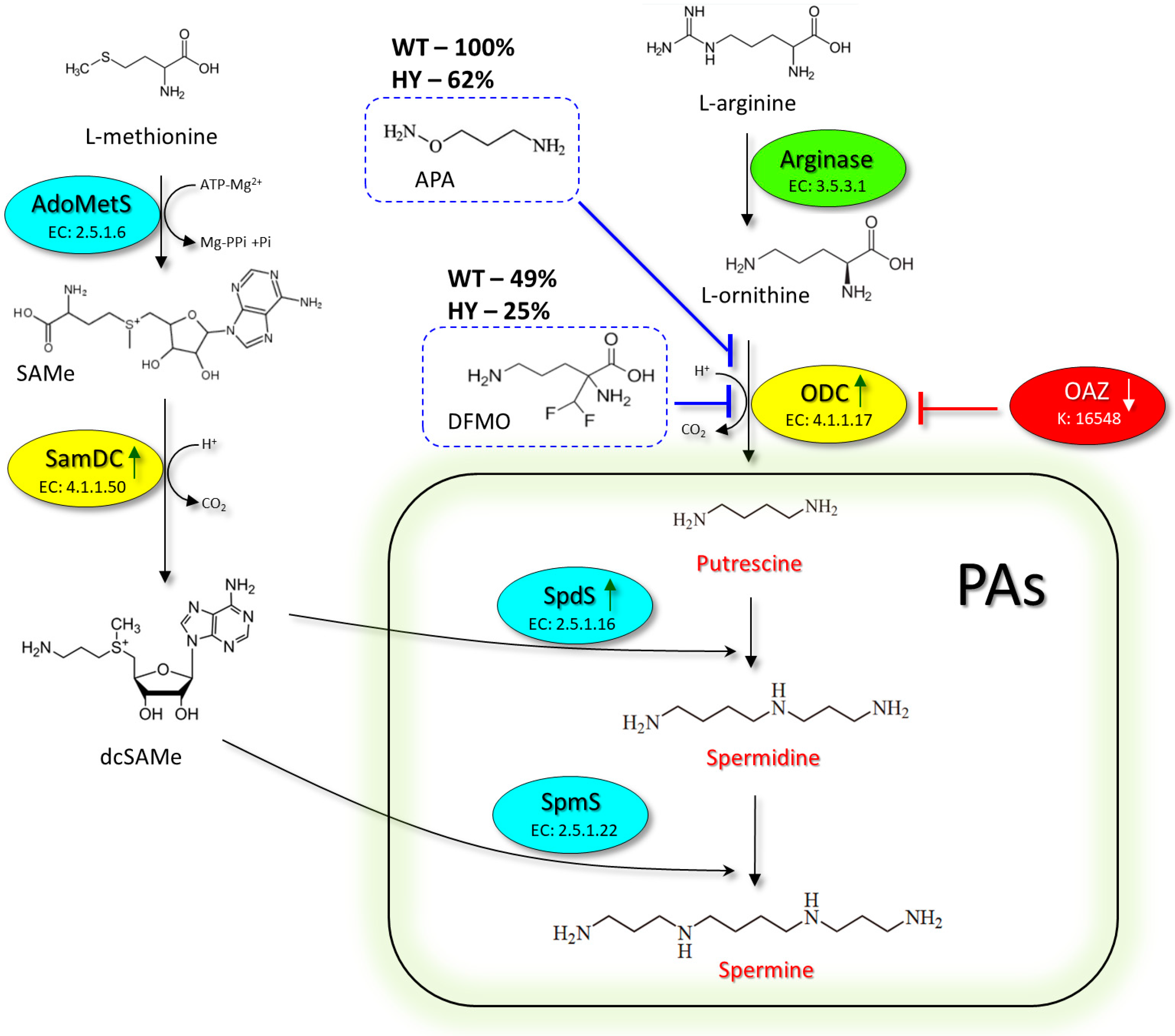
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Subhan, M.; Faryal, R.; Macreadie, I. Exploitation of Aspergillus terreus for the production of natural statins. J. Fungi 2016, 2, 13. [Google Scholar] [CrossRef]
- Stancu, C.; Sima, A. Statins: Mechanism of action and effects. J. Cell. Mol. Med. 2001, 5, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.V.; Varshney, V.K.; Pandey, A. Lovastatin: A Journey from Ascomycetes to Basidiomycetes Fungi. J. Biol. Act. Prod. Nat. 2019, 9, 162–178. [Google Scholar] [CrossRef]
- Manzoni, M.; Bergomi, S.; Rollini, M.; Cavazzoni, V. Production of statins by filamentous fungi. Biotechnol. Lett. 1999, 21, 253–257. [Google Scholar] [CrossRef]
- Javed, S.; Bukhari, S.; Zovia, I.; Meraj, M. Screening of Indigenously Isolated Fungi for Lovastatin Production and Its in vivo Evaluation. Curr. Pharm. Biotechnol. 2014, 15, 422–427. [Google Scholar] [CrossRef]
- Jia, Z.; Zhang, X.; Cao, X.; Liu, J.; Qin, B. Production of lovastatin by a self-resistant mutant of Aspergillus terreus. Ann. Microbiol. 2011, 61, 615–621. [Google Scholar] [CrossRef]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef] [Green Version]
- Barrios-González, J.; Pérez-Sánchez, A.; Bibián, M.E. New knowledge about the biosynthesis of lovastatin and its production by fermentation of Aspergillus terreus. Appl. Microbiol. Biotechnol. 2020, 1–20. [Google Scholar] [CrossRef]
- Dzhavakhija, V.G.; Voinova, T.M.; Vavilova, N.A.; Santsevich, N.I.; Vinokurova, N.G.; Kadomtseva, V.M.; Dzhavakhija, V.V.; Mishin, A.G. Fungus Strain Aspergillus Terreus 44-62 as Producer of Lovastatin, Industrial Method for Isolation of Lovastatin and Method for Lactoninization of Statins. Patent RU 2261901C2, 27 June 2005. [Google Scholar]
- Zhgun, A.A.; Dumina, M.V.; Voinova, T.M.; Dzhavakhiya, V.V.; Eldarov, M.A. Role of acetyl-CoA Synthetase and LovE Regulator Protein of Polyketide Biosynthesis in Lovastatin Production by Wild-Type and Overproducing Aspergillus terreus Strains. Appl. Biochem. Microbiol. 2018, 54, 188–197. [Google Scholar] [CrossRef]
- Zhgun, A.A.; Nuraeva, G.K.; Eldarov, M. The Role of LaeA and LovE Regulators in Lovastatin Biosynthesis with Exogenous Polyamines in Aspergillus terreus. Appl. Biochem. Microbiol. 2019, 55, 626–635. [Google Scholar] [CrossRef]
- Zhgun, A.A.; Nuraeva, G.K.; Dumina, M.V.; Voinova, T.M.; Dzhavakhiya, V.V.; Eldarov, M.A. 1,3-Diaminopropane and spermidine upregulate lovastatin production and expression of lovastatin biosynthetic genes in Aspergillus terreus via LaeA regulation. Appl. Biochem. Microbiol. 2019, 55, 244–255. [Google Scholar] [CrossRef]
- Peñalva, M.A.; Rowlands, R.T.; Turner, G. The optimization of penicillin biosynthesis in fungi. Trends Biotechnol. 1998, 16, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Zhgun, A.A.; Ivanova, M.A.; Domracheva, A.G.; Novak, M.I.; Elidarov, M.A.; Skryabin, K.G.; Bartoshevich, Y.E. Genetic transformation of the mycelium fungi Acremonium chrysogenum. Appl. Biochem. Microbiol. 2008, 44, 600–607. [Google Scholar] [CrossRef]
- Dumina, M.V.; Zhgun, A.A.; Domracheva, A.G.; Novak, M.I.; El’darov, M.A. Chromosomal polymorphism of Acremonium chrysogenum strains producing cephalosporin C. Russ. J. Genet. 2012, 48, 778–784. [Google Scholar] [CrossRef]
- Zhgun, A.A. Random Mutagenesis of Filamentous Fungi Stains for High-Yield Production of Secondary Metabolites: The Role of Polyamines. In Genotoxicity and Mutagenicity—Mechanisms and Test Methods; Soloneski, S., Ed.; IntechOpen: London, UK, 2020; pp. 1–17. ISBN 978-1-83880-042-0. [Google Scholar]
- Domratcheva, A.G.; Zhgun, A.A.; Novak, N.V.; Dzhavakhiya, V.V. The Influence of Chemical Mutagenesis on the Properties of the Cyclosporine a High-Producer Strain Tolypocladium inflatum VKM F-3630D. Appl. Biochem. Microbiol. 2018, 54, 53–57. [Google Scholar] [CrossRef]
- Bartoshevich, Y.; Novak, M.; Domratcheva, A.; Skryabin, K. Method of cephalosporin C biosynthesis by using new Acremonium chrysogenum strain RNCM NO F-4081D. Russian Federation 2,426,793, 2011. [Google Scholar]
- Martín, J.; García-Estrada, C.; Kosalková, K.; Ullán, R.V.; Albillos, S.M.; Martín, J.-F. The inducers 1,3-diaminopropane and spermidine produce a drastic increase in the expression of the penicillin biosynthetic genes for prolonged time, mediated by the laeA regulator. Fungal Genet. Biol. 2012, 49, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Zhgun, A.A.; Kalinin, S.G.; Novak, M.I.; Domratcheva, A.G.; Petukhov, D.V.; Dzhavakhiya, V.V.; Eldarov, M.A.; Bartoshevitch, I.E. The influence of polyamines on cephalosporine C biosynthesis in Acremonium chrysogenum strains. Izv Vuzov. Prikl Khim i Biotech. 2015, 14, 47–54. [Google Scholar]
- Hyvönen, M.T.; Keinänen, T.A.; Nuraeva, G.K.; Yanvarev, D.V.; Khomutov, M.; Khurs, E.N.; Kochetkov, S.N.; Vepsäläinen, J.; Zhgun, A.A.; Khomutov, A.R. Hydroxylamine analogue of agmatine: Magic bullet for arginine decarboxylase. Biomolecules 2020, 10, 406. [Google Scholar] [CrossRef] [Green Version]
- Khomutov, A.R.; Khomutov, R.M. Synthesis of putrescine and spermidine aminooxy analogues. Bioorganicheskaya Khimiya 1989, 15, 698–703. [Google Scholar]
- Zhgun, A.; Avdanina, D.; Shumikhin, K.; Simonenko, N.; Lyubavskaya, E.; Volkov, I.; Ivanov, V. Detection of potential biodeterioration risks for tempera painting in 16th century exhibits from State Tretyakov Gallery. PLoS ONE 2020, 15, e0230591. [Google Scholar] [CrossRef]
- Zhgun, A.A.; Avdanina, D.A.; Shagdarova, B.T.; Troyan, E.V.; Nuraeva, G.K.; Potapov, M.P.; Il’ina, A.V.; Shitov, M.V.; Varlamov, V.P. Search for Efficient Chitosan-Based Fungicides to Protect the 15th‒16th Centuries Tempera Painting in Exhibits from the State Tretyakov Gallery. Microbiology 2020, 89, 750–755. [Google Scholar] [CrossRef]
- Dumina, M.V.; Zhgun, A.A.; Kerpichnikov, I.V.; Domracheva, A.G.; Novak, M.I.; Valiachmetov, A.Y.; Knorre, D.A.; Severin, F.F.; Eldarov, M.A.; Bartoshevich, Y.E. Functional analysis of MFS protein CefT involved in the transport of beta-lactam antibiotics in Acremonium chrysogenum and Saccharomyces cerevisiae. Appl. Biochem. Microbiol. 2013, 49, 368–377. [Google Scholar] [CrossRef]
- Dumina, M.V.; Zhgun, A.A.; Novak, M.I.; Domratcheva, A.G.; Petukhov, D.V.; Dzhavakhiya, V.V.; Eldarov, M.A.; Bartoshevitch, I.E. Comparative gene expression profiling reveals key changes in expression levels of cephalosporin C biosynthesis and transport genes between low and high-producing strains of Acremonium chrysogenum. World J. Microbiol. Biotechnol. 2014, 30, 2933–2941. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; George, N.M.; Yassin, M.A.; Alaidaroos, B.A.; Bolbol, A.A.; Mohamed, M.S.; Rady, A.M.; Aziz, S.W.; Zayed, R.A.; Sitohy, M.Z. Purification and Characterization of Ornithine Decarboxylase from Aspergillus terreus; Kinetics of Inhibition by Various Inhibitors. Molecules 2019, 24, 2756. [Google Scholar] [CrossRef] [Green Version]
- Khomutov, A.R.; Dzhavakhiya, V.G.; Voinova, T.M.; Ermolinskii, B.S.; Khomutov, R.M. Aminooxy analogue of putrescine inhibits polyketide biosynthetic pathway of natural products. Bioorganicheskaya Khimiya 1989, 15, 706–709. [Google Scholar]
- Campbell, C.D.; Vederas, J.C. Biosynthesis of lovastatin and related metabolites formed by fungal iterative PKS enzymes. Biopolymers 2010, 93, 755–763. [Google Scholar] [CrossRef]
- Barrios-González, J.; Baños, J.G.; Covarrubias, A.A.; Garay-Arroyo, A. Lovastatin biosynthetic genes of Aspergillus terreus are expressed differentially in solid-state and in liquid submerged fermentation. Appl. Microbiol. Biotechnol. 2008, 79, 179–186. [Google Scholar] [CrossRef]
- Teresa, P.; Armando, M.; Javier, B.-G. Amplification of laeA Gene in Aspergillus terreus: A Strategy to Generate Lovastatin-Overproducing Strains for Solid-State Fermentation. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 537–555. [Google Scholar]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in aging and disease. Aging (Albany. NY) 2011, 3, 716–732. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of polyamines in mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campilongo, R.; Di Martino, M.L.; Marcocci, L.; Pietrangeli, P.; Leuzzi, A.; Grossi, M.; Casalino, M.; Nicoletti, M.; Micheli, G.; Colonna, B.; et al. Molecular and functional profiling of the polyamine content in enteroinvasive E. coli: Looking into the gap between commensal E. coli and harmful Shigella. PLoS ONE 2014, 9, e106589. [Google Scholar] [CrossRef]
- Valdés-Santiago, L.; Cervantes-Chávez, J.A.; León-Ramírez, C.G.; Ruiz-Herrera, J. Polyamine Metabolism in Fungi with Emphasis on Phytopathogenic Species. J. Amino Acids 2012, 2012, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nickerson, K.W.; Dunkle, L.D.; Van Etten, J.L. Absence of spermine in filamentous fungi. J. Bacteriol. 1977, 129, 173–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, M.; Russo, G.; Van Etten, J.; Nickerson, K. Polyamines in dimorphic fungi. Curr. Microbiol. 1979, 2, 187–190. [Google Scholar] [CrossRef]
- Hart, D.; Winther, M.; Stevens, L. Polyamine distribution and S-adenosyl methionine decarboxylase activity in filamentous fungi. FEMS Microbiol. Lett. 1978, 3, 173–175. [Google Scholar] [CrossRef]
- Sannazzaro, A.I.; Álvarez, C.L.; Menéndez, A.B.; Pieckenstain, F.L.; Albertó, E.O.; Ruiz, O.A. Ornithine and arginine decarboxylase activities and effect of some polyamine biosynthesis inhibitors on Gigaspora rosea germinating spores. FEMS Microbiol. Lett. 2004, 230, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R. Inhibition of polyamine biosynthesis in fungi. Mycol. Res. 1995, 99, 129–139. [Google Scholar] [CrossRef]
- Shapira, R.; Altman, A.; Henis, Y.; Chet, I. Polyamines and Ornithine Decarboxylase Activity during Growth and Differentiation in Sclerotium rolfsii. Microbiology 1989, 135, 1361–1367. [Google Scholar] [CrossRef] [Green Version]
- Rajam, M.V.; Galston, A.W. The Effects of Some Polyamine Biosynthetic Inhibitors on Growth and Morphology of Phytopathogenic Fungi. Plant Cell Physiol. 1985, 26, 683–692. [Google Scholar] [CrossRef]
- Pieckenstain, F.L.; Gárriz, A.; Chornomaz, E.M.; Sánchez, D.H.; Ruiz, O.A. The effect of polyamine biosynthesis inhibition on growth and differentiation of the phytopathogenic fungus Sclerotinia sclerotiorum. Antonie van Leeuwenhoek 2001, 80, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.; Keenan, J.; Cowley, T.; McPherson, A.; Havis, N. Inhibition of polyamine biosynthesis in Phytophthora infestans and Pythium ultimum. Plant Pathol. 1995, 44, 80–85. [Google Scholar]
- Akasaka, N.; Fujiwara, S. The therapeutic and nutraceutical potential of agmatine, and its enhanced production using Aspergillus oryzae. Amino Acids 2020, 52, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Soda, K. Polyamine Metabolism and Gene Methylation in Conjunction with One-Carbon Metabolism. Int. J. Mol. Sci. 2018, 19, 3106. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C.; Mato, J.M. S-adenosylmethionine in liver health, injury, and cancer. Physiol. Rev. 2012, 92, 1515–1542. [Google Scholar] [CrossRef] [Green Version]
- Perez-Leal, O.; Merali, S. Regulation of polyamine metabolism by translational control. Amino Acids 2012, 42, 611–617. [Google Scholar] [CrossRef]
- Zhgun, A.; Dumina, M.; Valiakhmetov, A.; Eldarov, M. The critical role of plasma membrane H+-ATPase activity in cephalosporin C biosynthesis of Acremonium chrysogenum. PLoS ONE 2020, 15, e0238452. [Google Scholar] [CrossRef]
- Murray Stewart, T.; Dunston, T.T.; Woster, P.M.; Casero, R.A. Polyamine catabolism and oxidative damage. J. Biol. Chem. 2018, 293, 18736–18745. [Google Scholar] [CrossRef] [Green Version]
- Ha, H.C.; Sirisoma, N.S.; Kuppusamy, P.; Zweier, J.L.; Woster, P.M.; Casero, R.A. The natural polyamine spermine functions directly as a free radical scavenger. Proc. Natl. Acad. Sci. USA 1998, 95, 11140–11145. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-Y.; Su, G.-C.; Huang, W.-Y.; Ko, M.-Y.; Yeh, H.-Y.; Chang, G.-D.; Lin, S.-J.; Chi, P. Promotion of homology-directed DNA repair by polyamines. Nat. Commun. 2019, 10, 65. [Google Scholar] [CrossRef]
- Kennedy, J.; Auclair, K.; Kendrew, S.G.; Park, C.; Vederas, J.C.; Hutchinson, C.R.; Auclais, K.; Kendrew, S.G.; Park, C.; Vederas, J.C.; et al. Modulation of polyketide synthase activity by accessory proteins during lovastatin biosynthesis. Science 1999, 284, 1368–1372. [Google Scholar] [CrossRef] [PubMed]
- Brosch, G.; Loidl, P.; Graessle, S. Histone modifications and chromatin dynamics: A focus on filamentous fungi. FEMS Microbiol. Rev. 2008, 32, 409–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarikaya-Bayram, Ö.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, Ö. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Gene | Product, Function | Oligonucleotide (Sequence 5→3) | Source Sequence |
|---|---|---|---|---|
| Act_RT_F | actin | A major component of the cytoskeleton | CCACGTTACCACTTTCAACTCC | XM_001209659.1, [10] |
| Act_RT_R | GAGGAGCGATGATCTTGACCT | |||
| E2_6_RT_F | ube2 6 | Ubiquitin-conjugating enzyme E2 6 | TGACCAGCGAAGAAATGACA | XM_001211932.1, [10] |
| E2_6_RT_R | TTATCTTTCATCCATTTCCA | |||
| LovA_F | lovA | Dihydromonacolin L monooxygenase | GCGATGTCAAGCCACTCCTCATTATG | AH007774.2, [12] |
| LovA_R | AGACCCAAGCTCCCAAGTACGTCAAG | |||
| LovB_F1 | lovB | Lovastatin nonaketide synthase | GCCCCATTCTATAAAAACCTGAGGATTC | AF151722, [12] |
| LovB_R1 | AGTCCTCATTATTCGAGACTCGCAGC | |||
| LovC_F1 | lovC | Enoyl reductase | GCAGAGGAGGTCTTTGACTATCG | AH007774.2, [10] |
| LovC_R1 | GACTCGACGTTGGTGATACAGTCG | |||
| LovD_F1 | lovD | 2-Methylbutyryl/monacolin J transesterase | GGATCTGGACGGAGAGAACTG | |
| LovD_R1 | CAGGGTTCCAGTTGGAAGAAC | |||
| LovE_F | lovE | GAL4-like transcription factor | TCGATGCGTCTACAGTGAGC | |
| LovE_R | TAGCTGTCCGGTGGATCAAG | |||
| LovF_F | lovF | Lovastatin diketide synthase | TTGCATCTTGCCATTCAGAG | |
| LovF_R | TCGAGTCAAATGAGTAGGA | |||
| LovG_F | lovG | Thioesterase | GCTCCGTTCCCTTCCTCTGCA | AH007774.2, [12] |
| LovG_R | GGGGTGTTGAGTCTGCCAGTCG | |||
| ODC_RT_F | odc | Ornithine decarboxylase | CCCCGGTGAGGAAGATGCGT | CH476600.1, [12] |
| ODC_RT_R | TCGATCTCCGCCTTGGACGC | |||
| SamDC_RT_F | samdc | S-adenosylmethionine decarboxylase | TACACGACCTCGCCGTCATCCT | CH476605.1, [12] |
| SamDC_RT_R | CCTTCCAGATCTCCTCCGACACG | |||
| SpdS_RT_F1 | spds | Spermidine synthase | GAAGGTCCTGGTCATTGGCGGT | CH476594.1, [12] |
| SpdS_RT_R1 | TCTTGAGGAACTCGAAGCCGTCG | |||
| AZ_RT_F | oaz | Antizyme, Ornithine decarboxylase inhibitor | ATCTCAGTCTCCGAAGCGTCCTGG | XM_001214792.1, [12] |
| AZ_RT_R | CGAGGATTTGTGACCGACATAAGTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhgun, A.A.; Nuraeva, G.K.; Volkov, I.A. High-Yielding Lovastatin Producer Aspergillus terreus Shows Increased Resistance to Inhibitors of Polyamine Biosynthesis. Appl. Sci. 2020, 10, 8290. https://doi.org/10.3390/app10228290
Zhgun AA, Nuraeva GK, Volkov IA. High-Yielding Lovastatin Producer Aspergillus terreus Shows Increased Resistance to Inhibitors of Polyamine Biosynthesis. Applied Sciences. 2020; 10(22):8290. https://doi.org/10.3390/app10228290
Chicago/Turabian StyleZhgun, Alexander A., Gulgina K. Nuraeva, and Ivan A. Volkov. 2020. "High-Yielding Lovastatin Producer Aspergillus terreus Shows Increased Resistance to Inhibitors of Polyamine Biosynthesis" Applied Sciences 10, no. 22: 8290. https://doi.org/10.3390/app10228290