Sialic Acids: An Important Family of Carbohydrates Overlooked in Environmental Biofilms

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pilot Set-Up
2.2. Samples Collection
2.3. Microscopic Pictures
2.4. EPS Extraction
2.5. Fourier Transform Infrared Spectroscopy (FTIR)
2.6. High-Resolution Mass Spectrometry (MS)
2.7. DNA Extraction and 16S rRNA Gene Amplicon Sequencing
2.8. Basic Local Alignment Search Tool (BLAST)
3. Results and Discussion
3.1. Sialic Acids Are Present in Environmental Biofilms
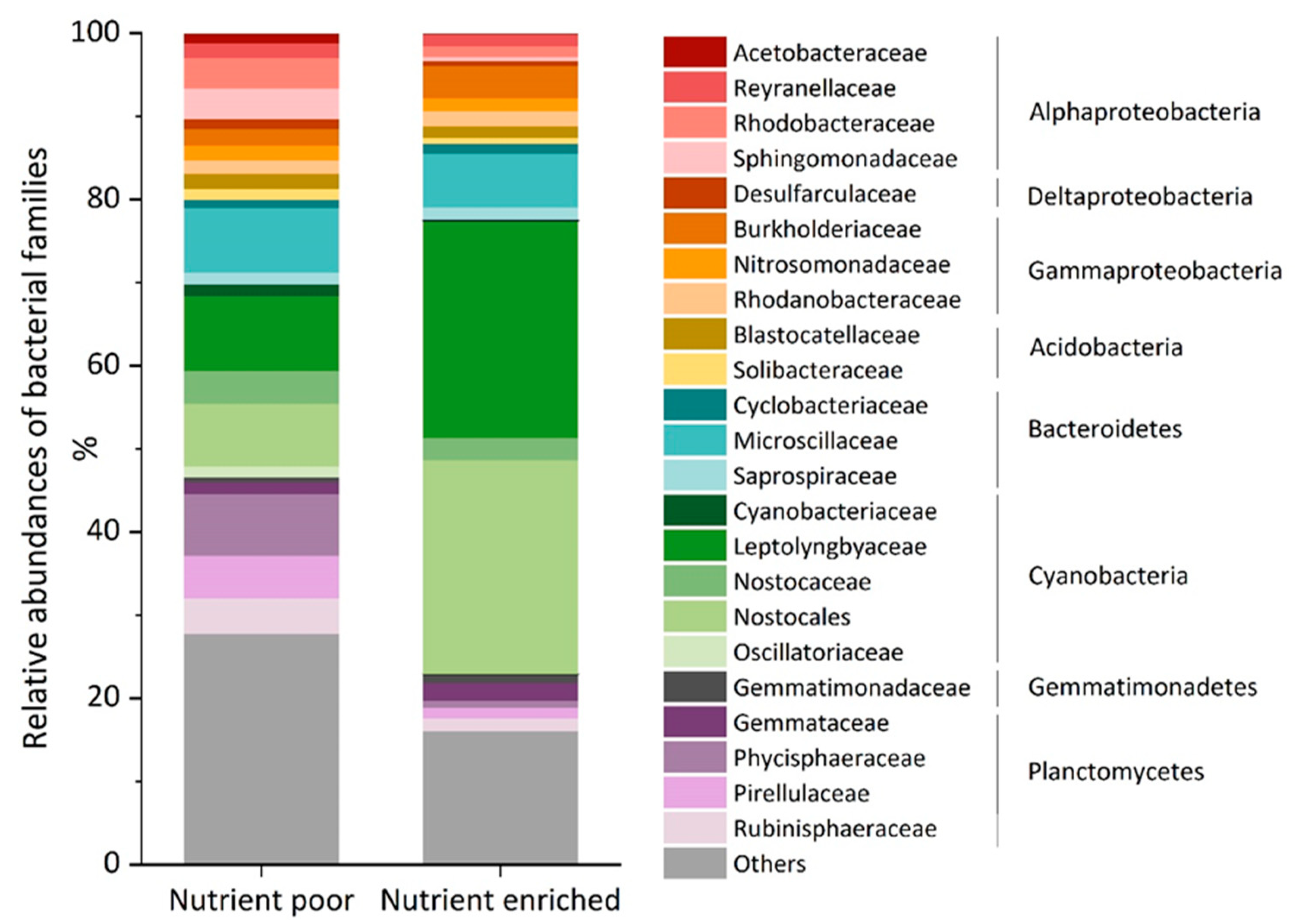
3.2. Nutrient Availability Did Not Significantly Influence the Production of Sialic Acids in the Environmental Biofilms
3.3. Relevance to Industrial Processes and Future Considerations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seviour, T.; Derlon, N.; Dueholm, M.S.; Flemming, H.-C.; Girbal-Neuhauser, E.; Horn, H.; Kjelleberg, S.; van Loosdrecht, M.C.M.; Lotti, T.; Malpei, M.F.; et al. Extracellular polymeric substances of biofilms: Suffering from an identity crisis. Water Res. 2019, 151, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Trappetti, C.; Kadioglu, A.; Carter, M.; Hayre, J.; Iannelli, F.; Pozzi, G.; Andrew, P.W.; Oggioni, M.R. Sialic acid: A preventable signal for pneumococcal biofilm formation, colonization, and invasion of the host. J. Infect. Dis. 2009, 199, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Stafford, G.; Roy, S.; Honma, K.; Sharma, A. Sialic acid, periodontal pathogens and tannerella forsythia: Stick around and enjoy the feast! Mol. Oral Microbiol. 2012, 27, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angata, T.; Varki, A. Chemical diversity in the sialic acids and related alpha-keto acids: An evolutionary perspective. Chem. Rev. 2002, 102, 439–469. [Google Scholar] [CrossRef]
- De Graaff, D.R.; Felz, S.; Neu, T.R.; Pronk, M.; van Loosdrecht, M.C.M.; Lin, Y. Sialic acids in the extracellular polymeric substances of seawater-adapted aerobic granular sludge. Water Res. 2019, 155, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Kleikamp, H.B.C.; Lin, Y.M.; McMillan, D.G.G.; Geelhoed, J.S.; Naus-Wiezer, S.N.H.; van Baarlen, P.; Saha, C.; Louwen, R.; Sorokin, D.Y.; van Loosdrecht, M.C.M.; et al. Tackling the chemical diversity of microbial nonulosonic acids—A universal large-scale survey approach. Chem. Sci. 2020, 11, 3074–3080. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.L.; Desa, N.; Hansen, E.E.; Knirel, Y.A.; Gordon, J.I.; Gagneux, P.; Nizet, V.; Varki, A. Innovations in host and microbial sialic acid biosynthesis revealed by phylogenomic prediction of nonulosonic acid structure. Proc. Natl. Acad. Sci. USA 2009, 106, 13552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinel, I.S.M.; Kim, L.H.; Proença Borges, V.R.; Farhat, N.M.; Witkamp, G.J.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Effect of phosphate availability on biofilm formation in cooling towers. Biofouling 2020, 1–16. [Google Scholar]
- Al-Bloushi, M.; Saththasivam, J.; Jeong, S.; Amy, G.L.; Leiknes, T. Effect of organic on chemical oxidation for biofouling control in pilot-scale seawater cooling towers. J. Water Process Eng. 2017, 20, 1–7. [Google Scholar] [CrossRef]
- Belila, A.; El-Chakhtoura, J.; Otaibi, N.; Muyzer, G.; Gonzalez-Gil, G.; Saikaly, P.E.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Bacterial community structure and variation in a full-scale seawater desalination plant for drinking water production. Water Res. 2016, 94, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbial. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenhofen, I.C.; Vinogradov, E.; Whitfield, D.M.; Brisson, J.-R.; Logan, S.M. The cmp-legionaminic acid pathway in campylobacter: Biosynthesis involving novel gdp-linked precursors. Glycobiology 2009, 19, 715–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lu, Z.; Allen, K.N.; Mariano, P.S.; Dunaway-Mariano, D. Human symbiont bacteroides thetaiotaomicron synthesizes 2-keto-3-deoxy-d-glycero-d- galacto-nononic acid (kdn). Chem. Biol. 2008, 15, 893–897. [Google Scholar] [CrossRef] [Green Version]
- Knirel, Y.A.; Shashkov, A.S.; Tsvetkov, Y.E.; Jansson, P.E.; Zãhringer, U. 5,7-diamino-3,5,7,9-tetradeoxynon-2-ulosonic acids in bacterial glycopolymers: Chemistry and biochemistry. ADV Carbohyd. Chem. Biochem. 2003, 58, 371–417. [Google Scholar]
- Harduin-Lepers, A.; Mollicone, R.; Delannoy, P.; Oriol, R. The animal sialyltransferases and sialyltransferase-related genes: A phylogenetic approach. Glycobiology 2005, 15, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Boleij, M.; Kleikamp, H.; Pabst, M.; Neu, T.R.; van Loosdrecht, M.C.M.; Lin, Y. Decorating the anammox house: Sialic acids and sulfated glycosaminoglycans in the extracellular polymeric substances of anammox granular sludge. Environ. Sci. Technol. 2020, 54, 5218–5226. [Google Scholar] [CrossRef]
- Wagstaff, B.A.; Rejzek, M.; Field, R.A. Identification of a kdn biosynthesis pathway in the haptophyte prymnesium parvum suggests widespread sialic acid biosynthesis among microalgae. J. Biol. Chem. 2018, 293, 16277–16290. [Google Scholar] [CrossRef] [Green Version]
- Ganss, B.; Hoffmann, W. Calcium binding to sialic acids and its effect on the conformation of ependymins. J. Biochem. 1993, 217, 275–280. [Google Scholar] [CrossRef]
- Di Dato, V.; Di Costanzo, F.; Barbarinaldi, R.; Perna, A.; Ianora, A.; Romano, G. Unveiling the presence of biosynthetic pathways for bioactive compounds in the thalassiosira rotula transcriptome. Sci. Rep. 2019, 9, 9893. [Google Scholar] [CrossRef]
- Badr, H.A.; AlSadek, D.M.; Mathew, M.P.; Li, C.-Z.; Djansugurova, L.B.; Yarema, K.J.; Ahmed, H. Nutrient-deprived cancer cells preferentially use sialic acid to maintain cell surface glycosylation. Biomaterials 2015, 70, 23–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, A.; Rishi, V.; Soni, S.K.; Rishi, P. A novel multi-enzyme preparation produced from aspergillus niger using biodegradable waste: A possible option to combat heterogeneous biofilms. AMB Express 2020, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Jia, R.; Li, Y.; Gu, T. Advances in the treatment of problematic industrial biofilms. World J. Microb. Biotechnol. 2017, 33, 97. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, P.; Mauerhofer, S.; Schneider, J.; Maniura-Weber, K.; Rosenberg, U.; Ren, Q. Enzymes enhance biofilm removal efficiency of cleaners. Antimicrob. Agents Chem. 2016, 60, 3647–3652. [Google Scholar] [CrossRef] [Green Version]
- Saggu, S.K.; Jha, G.; Mishra, P.C. Enzymatic degradation of biofilm by metalloprotease from microbacterium sp. SKS10. Front. Bioeng. Biotechnol. 2019, 7, 192. [Google Scholar] [CrossRef] [Green Version]
- Nahar, S.; Mizan, M.F.R.; Ha, A.J.-W.; Ha, S.-D. Advances and future prospects of enzyme-based biofilm prevention approaches in the food industry. Compr. Rev. Food. Sci. Food Saf. 2018, 17, 1484–1502. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.H.; Kim, S.-J.; Kim, C.-M.; Shin, M.S.; Kook, S.; Kim, I.S. Effects of enzymatic treatment on the reduction of extracellular polymeric substances (eps) from biofouled membranes. Desalin. Water Treat. 2013, 51, 6355–6361. [Google Scholar] [CrossRef]
- Li, J.; Nickel, R.; Wu, J.; Lin, F.; van Lierop, J.; Liu, S. A new tool to attack biofilms: Driving magnetic iron-oxide nanoparticles to disrupt the matrix. Nanoscale 2019, 11, 6905–6915. [Google Scholar] [CrossRef]
- Hotrum, N.E.; de Jong, P.; Akkerman, J.C.; Fox, M.B. Pilot scale ultrasound enabled plate heat exchanger–its design and potential to prevent biofouling. J. Food Eng. 2015, 153, 81–88. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | CMP-Pse Synthetase | CMP-Leg Synthetase | CMP-KDN Synthetase |
|---|---|---|---|---|
| Acetobacteraceae | ||||
| Roseomonas | ||||
| Blastocatellaceae | ||||
| Burkholderiaceae | ||||
| Hydrogenophaga | ||||
| Candidimonas | ||||
| Aquabacterium | ||||
| Limnobacter | ||||
| Noviherbaspirillum | ||||
| Ideonella | ||||
| Azohydromonas | ||||
| Cyanobacteriaceae | ||||
| Annamia HOs24 | ||||
| Geminocystis | ||||
| Cyclobacteriaceae | ||||
| Marinoscillum | ||||
| Ekhidna | ||||
| Algoriphagus | ||||
| Imperialibacter | ||||
| Desulfarculaceae | ||||
| Gemmataceae | ||||
| Gemmata | ||||
| Fimbriiglobus | ||||
| Gemmatimonadaceae | ||||
| Gemmatimonas | ||||
| Leptolyngbyaceae | ||||
| Leptolyngbya | ||||
| Microscillaceae | ||||
| Nitrosomonadaceae | ||||
| Nostocaceae | ||||
| Stigonema | ||||
| Nostoc | ||||
| Chlorogloeopsis | ||||
| Nostocales (undefined) | ||||
| Microseira wollei | ||||
| Oscillatoriaceae | ||||
| Oscillatoria | ||||
| Phycisphaeraceae | ||||
| Pirellulaceae | ||||
| Blastopirellula | ||||
| Bythopirellula | ||||
| Reyranellaceae | ||||
| Rhodanobacteraceae | ||||
| Aquimonas | ||||
| Rhodobacteraceae | ||||
| Rhodobacter | ||||
| Rubinisphaeraceae | ||||
| Planctomicrobium | ||||
| SH-PL14 | ||||
| Saprospiraceae | ||||
| Phaeodactylibacter | ||||
| Solibacteraceae | ||||
| Sphingomonadaceae | ||||
| Porphyrobacter | ||||
| Sphingopyxis | ||||
| Blastomonas | ||||
| Erythrobacter | ||||
| Sphingobium | ||||
| Altererythrobacter | ||||
| Sphingomonas |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinel, I.S.M.; Kleikamp, H.B.C.; Pabst, M.; Vrouwenvelder, J.S.; van Loosdrecht, M.C.M.; Lin, Y. Sialic Acids: An Important Family of Carbohydrates Overlooked in Environmental Biofilms. Appl. Sci. 2020, 10, 7694. https://doi.org/10.3390/app10217694
Pinel ISM, Kleikamp HBC, Pabst M, Vrouwenvelder JS, van Loosdrecht MCM, Lin Y. Sialic Acids: An Important Family of Carbohydrates Overlooked in Environmental Biofilms. Applied Sciences. 2020; 10(21):7694. https://doi.org/10.3390/app10217694
Chicago/Turabian StylePinel, Ingrid S.M., Hugo B.C. Kleikamp, Martin Pabst, Johannes S. Vrouwenvelder, Mark C.M. van Loosdrecht, and Yuemei Lin. 2020. "Sialic Acids: An Important Family of Carbohydrates Overlooked in Environmental Biofilms" Applied Sciences 10, no. 21: 7694. https://doi.org/10.3390/app10217694