Rapid and Positive Effect of Bicarbonate Addition on Growth and Photosynthetic Efficiency of the Green Microalgae Chlorella Sorokiniana (Chlorophyta, Trebouxiophyceae)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strains and Growth Conditions
2.2. Pigment Contents
2.3. Photosynthetic Efficiency
2.4. Evaluation of Microbial Load Reduction
2.5. Statistical Analysis
3. Results
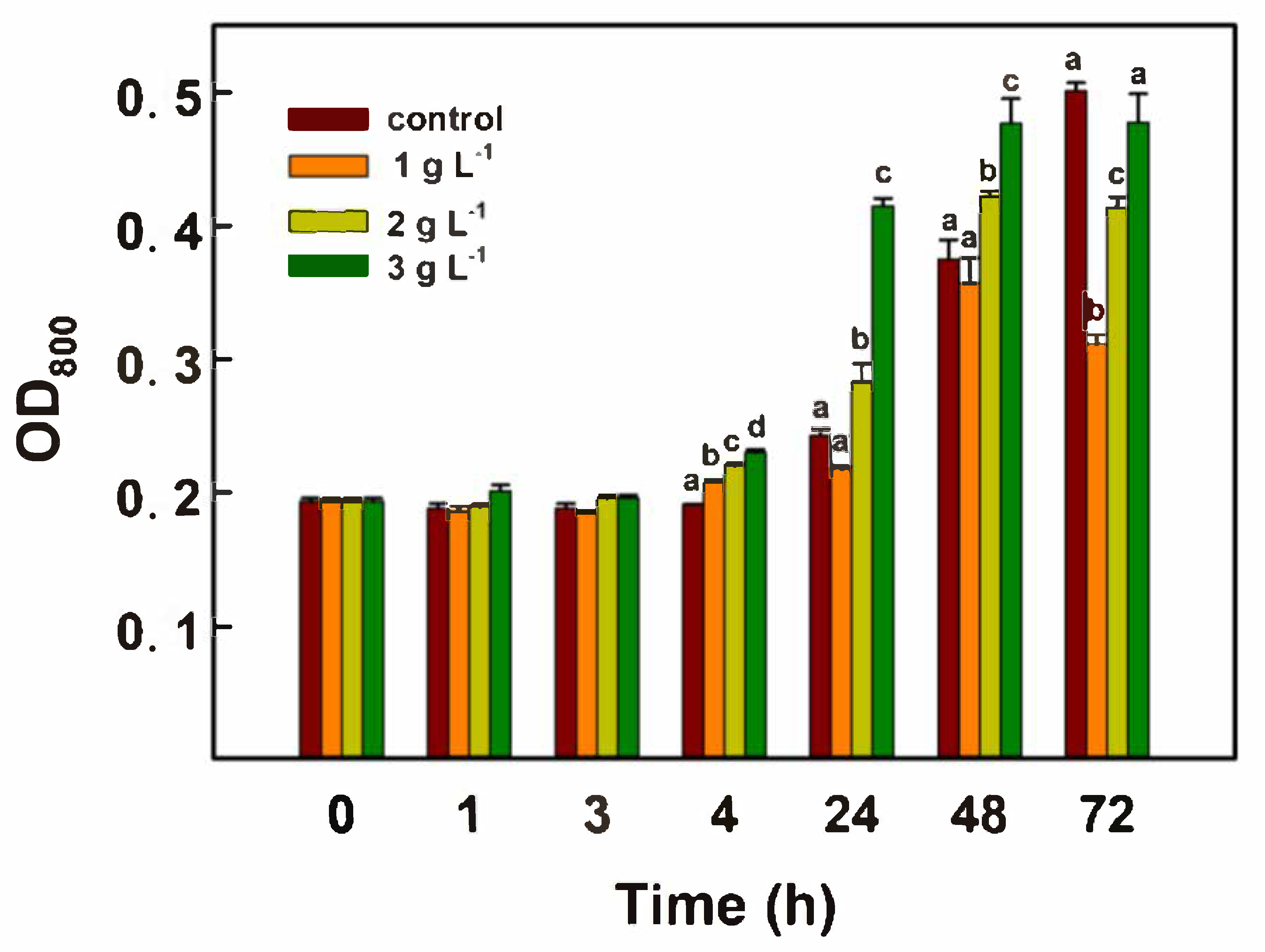
3.1. Impact of Sodium Bicarbonate Supplementation on Chlorella Growth
3.2. Impact of Sodium Bicarbonate Supplementation on Pigments
3.3. Impact of Sodium Bicarbonate Supplementation on Pigments Photosystem II Efficiency
3.4. Evaluation of Microbial Load Reduction
3.5. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal Biomass Utl. 2012, 3, 89–100. [Google Scholar]
- Barros, A.I.; Gonçalves, A.L.; Simões, M.; Pires, J.C.M. Harvesting techniques applied to microalgae: A review. Renew. Sustain. Energy Rev. 2015, 41, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Carfagna, S.; Salbitani, G.; Bottone, C.; Vona, V. Galdieria sulphuraria as a Possible Source of Food Colorant. J. Nutr. Ecol. 2016, 3, 78–81. [Google Scholar] [CrossRef] [Green Version]
- Vona, V.; Di Martino Rigano, V.; Andreoli, C.; Lobosco, O.; Caiazzo, M.; Martello, A.; Carfagna, S.; Salbitani, G.; Rigano, C. Comparative analysis of photosynthetic and respiratory parameters in the psychrophilic unicellular green alga Koliella antarctica, cultured in indoor and outdoor photo-bioreactors. Physiol. Mol. Biol. Plants 2018, 24, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Carfagna, S.; Landi, V.; Coraggio, F.; Salbitani, G.; Vona, V.; Pinto, G.; Pollio, A.; Ciniglia, C. Different characteristics of C-phycocyanin (C-PC) in two strains of the extremophilic Galdieria phlegrea. Algal. Res. 2018, 31, 406–412. [Google Scholar] [CrossRef]
- Bottone, C.; Camerlingo, R.; Miceli, R.; Salbitani, G.; Sessa, G.; Pirozzi, G.; Carfagna, S. Antioxidant and anti-proliferative properties of extracts from heterotrophic cultures of Galdieria sulphuraria. Nat. Prod. Res. 2019, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G.; Demurtas, O.; Ferrante, P. Le microalghe come bio-fabbriche per composti ad elevato valore aggiunto. ENEA Magazine, EAI Speciale. Biotecnologie per lo Sviluppo Sostenibile 2013, I, 61–66. [Google Scholar]
- Serive, B.; Kaas, R.; Bérard, J.B.; Pasquet, V.; Picot, L.; Cadoret, J.P. Selection and optimisation of a method for efficient metabolites extraction from microalgae. Bioresour. Technol. 2012, 124, 311–320. [Google Scholar] [CrossRef]
- Basu, S.; Mackey, K.R.M. Phytoplankton as Key Mediators of the Biological Carbon Pump: Their Responses to a Changing Climate. Sustainability 2018, 10, 869. [Google Scholar] [CrossRef] [Green Version]
- Poschenrieder, C.; Fernandez, J.A.; Rubio, L.; Perez, L.; Teres, J.; Barcelo, J. Transport and use of bicarbonate in plants: Current knowledge and challenges ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, L. Why are most aquatic plants widely distributed? Dispersal, clonal growth and small-scale heterogeneity in a stressful environment. Acta Oecol. 2002, 23, 137–154. [Google Scholar] [CrossRef]
- Hussner, A.; Mettler-Altmann, T.; Weber, A.P.M.; Sand-Jensen, K. Acclimation of photosynthesis to supersaturated CO2 in aquatic plant bicarbonate users. Freshw. Biol. 2016, 61, 1720–1732. [Google Scholar] [CrossRef]
- Cole, J.J.; Prairie, Y.T. Dissolved CO2. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2009; Volume 2, pp. 30–34. [Google Scholar]
- Huertas, I.E.; Colman, B.; Espie, G.; Lubian, L.M. Active transport of CO2 by three species of marine microalgae. J. Phycol. 2000, 36, 314–320. [Google Scholar] [CrossRef]
- White, D.A.; Pagarette, A.; Rooks, P.; Ali, S.T. The effect of sodium bicarbonate supplementation on growth and biochemical composition of marine microalgae cultures. J. Appl. Phycol. 2012, 25, 153–165. [Google Scholar] [CrossRef]
- Mondal, M.; Khanra, S.; Tiwari, O.N.; Gayen, K.; Halder, G.N. Role of carbonic anhydrase on the way to biological carbon capture through microalgae-A mini review. Environ. Prog. Sustain. Energy 2016. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant. Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [Green Version]
- Reinfelder, J.R. Carbon concentrating mechanisms in eukaryotic marine phytoplankton. Ann. Rev. Mar. Sci. 2011, 3, 291–315. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Wu, Y. Effects of Zn Deficiency and Bicarbonate on the Growth and Photosynthetic Characteristics of Four Plant Species. PLoS ONE 2017, 12, e0169812. [Google Scholar] [CrossRef]
- Acièn, F.G.; Gonz’alez, C.V.; Fern´andez, J.M.; Molina, E. Conversion of CO2 into biomass by microalgae: How realistic a contribution may it be to significant CO2 removal? Appl. Microbiol. Biotechnol. 2012, 96, 577–586. [Google Scholar] [CrossRef]
- Nunez, M.; Quigg, A. Changes in growth and composition of the marine microalgae Phaeodactylum tricornutum and Nannochloropsis salina in response to changing sodium bicarbonate concentrations. J. Appl. Phycol. 2015, 28, 2123–2138. [Google Scholar] [CrossRef]
- De Farias Silva, C.E.; Grisa, B.; Sforza, E.; La Rocca, N.; Bertucco, A. Effects of Sodium Bicarbonate on Biomass and Carbohydrate Production in Synechococcus PCC 7002. Chem. Eng. Trans. 2016, 49, 241–246. [Google Scholar]
- Kim, G.Y.; Roh, K.; Han, J.I. The use of bicarbonate for microalgae cultivation and its carbon footprint analysis. Green Chem. 2019, 21, 5053–5062. [Google Scholar] [CrossRef]
- Gardner, R.D.; Lohman, E.; Gerlach, R.; Cooksey, K.E.; Peyton, B.M. Comparison of CO2 and bicarbonate as inorganic carbon sources for triacylglycerol and starch accumulation in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2012, 110, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pancha, I.; Chokshi, K.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Bicarbonate supplementation enhanced biofuel production potential as well as nutritional stress mitigation in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 193, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Mageswari, A.; Subramanian, P.; Suganthi, C.; Chaitanyakumar, A.; Aswini, V.; Gothandam, K.M. Bicarbonate supplementation enhances growth and biochemical composition of Dunaliella salina V-101 by reducing oxidative stress induced during macronutrient deficit conditions. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salbitani, G.; Barone, C.M.A.; Carfagna, S. Effect of bicarbonate on growth of the oleaginous microalga Botryococcus braunii. Int. J. Plant Biol. 2019, 10, 8273. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.L.; Wang, J.H.; Cheng, L.Y.; Tang, Y.J.; Chi, Z.Y. Selection of microalgae strains for bicarbonate based integrated carbon capture and algal production system to produce lipid. Int. J. Green Energy 2019, 16, 825–833. [Google Scholar] [CrossRef]
- Salbitani, G.; Del Prete, S.; Bolinesi, F.; Mangono, O.; De Luca, V.; Carginale, V.; Donald, W.A.; Supuran, C.T.; Carfagna, S.; Capasso, C. Use of an immobilized thermostable α-CA (SspCA) for enhancing the metabolic efficiency of the freshwater green microalga Chlorella sorokiniana. J. Enzyme Inhib. Med. Chem. 2020, 35, 913–920. [Google Scholar] [CrossRef] [Green Version]
- De Andrade, C.J.; De Andrade, L.M. An overview on the application of genus Chlorella in biotechnological processes. J. Adv. Res. Biotechnol. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Sampathkumar, S.J.; Gothandam, K.M. Sodium bicarbonate augmentation enhances lutein biosynthesis in green microalgae Chlorella pyrenoidosa. Biocatal. Agric. Biotechnol. 2019, 22, 101406. [Google Scholar] [CrossRef]
- Vadlamani, A.; Pendyala, B.; Viamajala, S.; Varanasi, S. High Productivity Cultivation of Microalgae without Concentrated CO2 Input. ACS Sustain. Chem. Eng. 2019, 7, 1933–1943. [Google Scholar] [CrossRef]
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M. Chlorella for protein and biofuels: From strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotech. Biofuels 2014, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Cazzaniga, S.; Dall’Osto, L.; Szaub, J.; Scibilia, L.; Ballottari, M.; Purton, S.; Bassi, R. Domestication of the green alga Chlorella sorokiniana: Reduction of antenna size improves light-use efficiency in a photobioreactor. Biotechnol. Biofuels 2014, 7, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Li, C.; Lan, C.Q.; Liao, D. Effects of sodium bicarbonate on cell growth, lipid accumulation, and morphology of Chlorella vulgaris. Microb. Cell Fact. 2018, 17, 111. [Google Scholar] [CrossRef] [PubMed]
- Lohman, E.; Gardner, R.D.; Pedersen, T.; Peyton, B.M.; Cooksey, K.E.; Gerlach, R. Optimized carbon regime for enhanced growth and lipid accumulation in Chlorella vulgaris. Biotech. Biofuel 2015, 8, 82. [Google Scholar] [CrossRef] [Green Version]
- Jayasankar, R.; Valsala, K.K. Influence of different concentrations of bicarbonate on growth rate and chlorophyll content of Chlorella salina. J. Mar. Biol. Assoc. India 2008, 50, 74–78. [Google Scholar]
- Salbitani, G.; Wirtz, M.; Hell, R.; Carfagna, S. Affinity purification of O-acetylserine (thiol) lyase from Chlorella sorokiniana by recombinant proteins from Arabidopsis thaliana. Metabolites 2014, 4, 629–639. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Lorenzen, C.J.; Holmes, R.W.; Strickland, J.D. Fluorometric determination of chlorophyll. J. Cons. Int. Explor. Mer. 1965, 30, 3–15. [Google Scholar] [CrossRef]
- Bolinesi, F.; Arienzo, M.; Donadio, C.; Ferrara, L.; Passarelli, A.; Saggiomo, M.; Saggiomo, V.; Stanislao, C.; Trifuoggi, M.; Mangoni, O. Spatial and temporal variation of phytoplankton community structure in a coastal marine system subjected to human pressure. Reg. Stud. Mar. Sci. 2020, 35, 101198. [Google Scholar] [CrossRef]
- Vidussi, F.; Claustre, H.; Bustillos-Guzman, J.; Cailliau, C.; Marty, J.C. Determination of chlorophylls and carotenoids of marine phytoplankton: Separation of chlorophyll a from divinylchlorophyll a and zeaxanthin from lutein. J. Plankton Res. 1996, 18, 2377–2382. [Google Scholar] [CrossRef] [Green Version]
- Brunet, C.; Mangoni, O. Determinazione quali-quantitativa dei pigmenti fitoplanctonici mediante HPLC. In Metodologie di Studio del Plancton Marino; Socal, G., Buttino, I., Cabrini, M., Mangoni, O., Penna, A., Totti, C., Eds.; Istituto Superiore per la Protezione e la Ricerca Ambientale: Roma, Italy, 2010; Volume 56, pp. 379–385. [Google Scholar]
- Mauzerall, D.; Greenbaum, N.L. The absolute size of a photosynthetic unit. Biochim. Biophys. Acta 1989, 974, 119–140. [Google Scholar] [CrossRef]
- Falkowski, P.; Raven, J. Aquatic Photosynthesis, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007; p. 448. [Google Scholar]
- Mangoni, O.; Saggiomo, V.; Bolinesi, F.; Margiotta, F.; Budillon, G.; Cotroneo, Y.; Misic, C.; Rivaro, P.; Saggiomo, M. Phytoplankton blooms during austral summer in the Ross Sea, Antarctica: Driving factors and trophic implications. PLoS ONE 2017, 12, e0176033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, J.J.; Davis, R.F. The blank can make a big difference in oceanographic measurements. Limnol. Oceanogr. Bull. 2003, 12, 29–355. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Schreiber, U.; Klughammer, C.; Kolbowski, J. Assessment of wavelength-dependent parameters of photosynthetic electron transport with a new type of multi-color PAM chlorophyll fluorometer. Photosynth. Res. 2012, 113, 127. [Google Scholar] [CrossRef] [Green Version]
- International Organization for Standardization. Microbiology of Food and Animal Feeding Stuffs—General Requirements and Guidance for Microbiological Examinations—Amendment 1; ISO 7218:2007/Amd 1:2013, Corrected Version 15 April 2014; International Organization for Standardization: Geneva, Switzerland, 2014. [Google Scholar]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Schumann, F.K.; Lorenzen, C.J. Quantitative degradation of chlorophyll by a marine herbivores. Limnol. Oceanogr. 1975, 2, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Collos, Y.; Husseini-Ratrema, J.; Bec, B.; Vaquer, A.; Hoai, T.L.; Rougier, C.; Pons, V.; Souchu, P. Pheopigment dynamics, zooplankton grazing rates and the autumnal ammonium peak in a Mediterranean lagoon. Hydrobiologia 2005, 550, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Richmond, A.; Hu, Q. Handbook of Microalgal Culture: Applied Phycology and Biotechnology; Wiley Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Chi, Z.; Elloy, F.; Xie, Y.; Hu, Y.; Chen, S. Selection of microalgae and cyanobacteria strains for bicarbonate-based integrated carbon capture and algae production system. Appl. Biochem. Biotechnol. 2014, 172, 447–457. [Google Scholar] [CrossRef]
- Russo, M.E.; Bareschino, P.; Olivieri, G.; Chirone, R.; Salatino, P.; Marzocchella, A. Modeling of slurry staged bubble column for biomimetic CO2 capture. Int. J. Greenh Gas Con. 2016, 47, 200–209. [Google Scholar] [CrossRef]
- Peirce, S.; Perfetto, R.; Russo, M.E.; Capasso, C.; Rossi, M.; Salatino, P.; Marzocchella, A. Characterization of technical grade carbonic anhydrase as biocatalyst for CO2 capture in potassium carbonate solutions. Green Gas Sci. Technol. 2018, 8, 279–291. [Google Scholar] [CrossRef]
- Peng, L.; Lan, C.Q.; Zhang, Z. Cultivation of Freshwater Green Alga Neochloris oleoabundans. Can. J. Chem. Eng. 2016, 94, 439–445. [Google Scholar] [CrossRef]
- Work, V.H.; Randor Radakovits, R.; Jinkerson, R.E.; Meuser, J.E.; Elliott, L.J.; Vinyard, D.J.; Laurens, L.M.; Dismukes, G.C.; Posewitz, M.C. Increased Lipid Accumulation in the Chlamydomonas reinhardtii sta7-10 Starchless Isoamylase Mutant and Increased Carbohydrate Synthesis in Complemented Strains. Eukaryot. Cell. 2010, 9, 1251–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Jiang, J.G. Osmotic responses of Dunaliella to the changes of salinity. J. Cell Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Chlorophyll degradation during senescence. Annu. Rev. Plant Biol. 2006, 57, 55–77. [Google Scholar] [CrossRef]
- Borghini, F.; Colacevich, A.; Bergamino, N.; Micarelli, P.; Dattilo, A.M.; Focardi, S.; Loiselle, S. The microalgae Tetraselmis suecica in mesocosms under different light regimes. Chem. Ecol. 2009, 25, 345–357. [Google Scholar] [CrossRef]
- Sade, N.; Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-induced senescence and plant tolerance to abiotic stress. J. Exp. Bot. 2018, 69, 845–853. [Google Scholar] [CrossRef]
- Kolber, Z.; Wyman, K.D.; Falkowsky, P.G. Natural variability in photosynthetic energy conversion efficiency: A field study in the Gulf of Maine. Limnol. Oceanogr. 1990, 35, 72–79. [Google Scholar] [CrossRef]
- Geider, R.J.; Greene, R.M.; Kolber, Z.; MacIntyre, H.L. Falkowski, P.G. Fluorescence assessment of the maximum quantum efficiency of photosynthesis in the western North Atlantic. Deep-Sea Res. 1993, 1, 1205–1224. [Google Scholar] [CrossRef]
- Greene, R.I.M.; Kolber, Z.S.; Swift, D.G.; Tindale, N.W.; Falkowski, P.G. Physiological limitation of phytoplankton photosynthesis in the Equatorial Pacific determined from natural variability In the quantum yield of fluorescence. Limnol. Oceanogr. 1994, 39, 1061–1064. [Google Scholar] [CrossRef]
- Sugget, D.J.; MacIntyre, H.L.; Kana, T.M.; Geider, R.J. Comparing electron transport with gas exchange: Parameterising exchange rates between alternative photosynthetic currencies for eukaryotic phytoplankton. Aquat. Microb. Ecol. 2009, 56, 147–162. [Google Scholar] [CrossRef] [Green Version]
- Caffarri, S.; Tibiletti, T.; Jennings, R.C.; Santabarbara, S.A. Comparison Between Plant Photosystem I and Photosystem II Architecture and Functioning. Curr. Protein Pept. Sci. 2014, 15, 296–331. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | |||||||
|---|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 4 | 24 | 48 | 72 | |
| Control | 0.99 ± 0.08 Aa | 1.01 ± 0.01 Aa | 1.00 ± 0.02 Aa | 1.02 ± 0.05 Aa | 0.96 ± 0.06 Aa | 0.97 ± 0.02 Aa | 1.00 ± 0.04 Aa |
| 1 g L−1 | 1.00 ± 0.01 Aa | 0.98 ± 0.07 Aa | 1.31 ± 0.07 Bb | 0.94 ± 0.03 Aa | 0.83 ± 0.07 Cc | 0.88 ± 0.03 Aab | 0.98 ± 0.02 Aa |
| 2 g L−1 | 0.99 ± 0.07 Aa | 1.04 ± 0.04 Aa | 1.28 ± 0.08 Bb | 0.96 ± 0.04 Aa | 0.81 ± 0.08 Cac | 0.79 ± 0.02 Cc | 0.90 ± 0.05 Cc |
| 3 g L−1 | 0.99 ± 0.08 Aa | 0.99 ± 0.02 Aa | 1.22 ± 0.06 Bb | 1.00 ± 0.02 Aa | 0.63 ± 0.02 Dd | 0.76 ± 0.01 Cc | 0.78 ± 0.02 Cc |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salbitani, G.; Bolinesi, F.; Affuso, M.; Carraturo, F.; Mangoni, O.; Carfagna, S. Rapid and Positive Effect of Bicarbonate Addition on Growth and Photosynthetic Efficiency of the Green Microalgae Chlorella Sorokiniana (Chlorophyta, Trebouxiophyceae). Appl. Sci. 2020, 10, 4515. https://doi.org/10.3390/app10134515
Salbitani G, Bolinesi F, Affuso M, Carraturo F, Mangoni O, Carfagna S. Rapid and Positive Effect of Bicarbonate Addition on Growth and Photosynthetic Efficiency of the Green Microalgae Chlorella Sorokiniana (Chlorophyta, Trebouxiophyceae). Applied Sciences. 2020; 10(13):4515. https://doi.org/10.3390/app10134515
Chicago/Turabian StyleSalbitani, Giovanna, Francesco Bolinesi, Mario Affuso, Federica Carraturo, Olga Mangoni, and Simona Carfagna. 2020. "Rapid and Positive Effect of Bicarbonate Addition on Growth and Photosynthetic Efficiency of the Green Microalgae Chlorella Sorokiniana (Chlorophyta, Trebouxiophyceae)" Applied Sciences 10, no. 13: 4515. https://doi.org/10.3390/app10134515