
Neuropsychological Evidence Underlying Counterclockwise Bias in Running: Electroencephalography and Functional Magnetic Resonance Imaging Studies of Motor Imagery


Abstract
:1. Introduction
2. Methods
2.1. Participants
2.2. Instruments and Paradigm
2.2.1. Questionnaire upon Mental Imagery (QMI)
2.2.2. Experimental Paradigms for the EEG and ERP Studies
2.2.3. Experimental Paradigm for the fMRI Study
2.3. EEG Acquisition and Analysis
2.4. fMRI Acquisition and Analysis
2.5. Statistical Analysis
3. Results
3.1. EEG Hemispheric Asymmetry
3.2. ERPs
3.3. fMRI
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Toussaint, Y.; Fagard, J. A counterclockwise bias in running. Neurosci. Lett. 2008, 442, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Golomer, E.; Rosey, F.; Dizac, H.; Mertz, C.; Fagard, J. The influence of classical dance training on preferred supporting leg and whole body turning bias. Laterality 2009, 14, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Kim, Y.; Lee, J.; Jung, H.; Kim, J. An age related difference in turning bias. JKPESAGW 2018, 32, 101–111. [Google Scholar]
- Adámková, J.; Benediktová, K.; Svoboda, J.; Bartoš, L.; Vynikalová, L.; Nováková, P.; Burda, H. Turning preference in dogs: North attracts while south repels. PLoS ONE 2021, 16, e0245940. [Google Scholar] [CrossRef]
- Gordon, H.W.; Busdiecker, E.C.; Bracha, H.S. The relationship between leftward turning bias and visuospatial ability in humans. Int. J. Neurosci. 1992, 65, 29–36. [Google Scholar] [CrossRef]
- Streuli, J.C.; Obrist, G.; Brugger, P. Childrens’ left-turning preference is not modulated by magical ideation. Laterality 2017, 22, 90–104. [Google Scholar] [CrossRef] [Green Version]
- Scharine, A.A.; McBeath, M.K. Right-handers and Americans favor turning to the right. Hum. Factors 2002, 44, 248–256. [Google Scholar] [CrossRef]
- Taylor, M.J.D.; Strike, S.C.; Dabnichki, P. Turning bias and lateral dominance in a sample of able-bodied and amputee participants. Laterality 2007, 12, 50–63. [Google Scholar] [CrossRef]
- Tavakkoli, M.H.; Jose, T.P. The reason why do athletes run around the track counter-clockwise. Int. J. Educ. Dev. 2013, 2, 23–30. [Google Scholar]
- Brown, P. Why Do Athletes Have to Race around the Track in an Anti-Clockwise Direction? London. 2011. Available online: www.guardian.co.uk/notesandqueries/query/0,5753,-1416,00.html (accessed on 30 November 2019).
- Nicholls, M.E.; Clode, D.; Wood, S.J.; Wood, A.G. Laterality of expression in portraiture: Putting your best cheek forward. Proceedings of the Royal Society of London. Ser. B Biol. Sci. 1999, 266, 1517–1522. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, M.; Van Overschelde, S.; De Rycke, M.; Musch, E. Intrinsic and extrinsic factors of turning preferences in humans. Neurosci. Lett. 2016, 393, 179–183. [Google Scholar] [CrossRef]
- Kim, Y.K.; Park, E.; Lee, A.; Im, C.H.; Kim, Y.H. Changes in network connectivity during motor imagery and execution. PLoS ONE 2018, 13, e0190715. [Google Scholar] [CrossRef]
- Kober, S.E.; Grössinger, D.; Wood, G. Effects of motor imagery and visual neurofeedback on activation in the swallowing network: A real-time fMRI study. Dysphagia 2019, 34, 879–895. [Google Scholar] [CrossRef]
- Hardwick, R.M.; Caspers, S.; Eickhoff, S.B.; Swinnen, S.P. Neural correlates of motor imagery, action observation, and movement execution: A comparison across quantitative meta-analyses. BioRxiv 2017, 198432. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, P.W. A shortened form of Betts’ questionnaire upon mental imagery. J. Clin. Psychol. 1967, 23, 386–389. [Google Scholar] [CrossRef]
- Park, M.J.; Park, S.H. Effect of Positive Mental Imagery Stimuli on Anhedonic Depressive Symptoms. Korean J. Clin. Psychol. 2022, 41, 1–10. [Google Scholar] [CrossRef]
- Qiu, Z.; Allison, B.Z.; Jin, J.; Zhang, Y.; Wang, X.; Li, W.; Cichocki, A. Optimized motor imagery paradigm based on imagining Chinese characters writing movement. IEEE Trans. Neural Syst. Rehabil. Eng. 2017, 25, 1009–1017. [Google Scholar] [CrossRef]
- Tang, Z.C.; Li, C.; Wu, J.F.; Liu, P.C.; Cheng, S.W. Classification of EEG-based single-trial motor imagery tasks using a B-CSP method for BCI. Front. Inf. Technol. Electron. Eng. 2019, 20, 1087–1098. [Google Scholar] [CrossRef]
- Dai, M.; Zheng, D.; Na, R.; Wang, S.; Zhang, S. EEG classification of motor imagery using a novel deep learning framework. Sensors 2019, 19, 551. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kwon, M.; Lee, J.; Jung, H.; Kim, J.G. The neural mechanism of exercise addiction as determined by functional magnetic resonance imaging (fMRI). JKPESAGW 2018, 32, 69–80. [Google Scholar]
- Jasper, H.H. The ten-twenty electrode system of the International Federation. Electroencephalogr. Clin. Neurophysiol. 1958, 10, 370–375. [Google Scholar]
- Henriques, J.B.; Davidson, R.J. Brain electrical asymmetries during cognitive task performance in depressed and nondepressed subjects. Biol. Psychiatry 1997, 42, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Coan, J.A.; Allen, J.J.; Harmon-Jones, E. Voluntary facial expression and hemispheric asymmetry over the frontal cortex. Psychophysiology 2001, 38, 912–925. [Google Scholar] [CrossRef]
- Wyczesany, M.; Capotosto, P.; Zappasodi, F.; Prete, G. Hemispheric asymmetries and emotions: Evidence from effective connectivity. Neuropsychologia 2018, 121, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Reznik, S.J.; Allen, J.J. Frontal asymmetry as a mediator and moderator of emotion: An updated review. Psychophysiology 2018, 55, e12965. [Google Scholar] [CrossRef]
- Yang, M.; Deng, X.; An, S. The relationship between habitual use and real-time emotion regulation strategies in adolescents: Evidence from frontal EEG asymmetry. Neuropsychologia 2021, 162, 108056. [Google Scholar] [CrossRef]
- Tandle, A.L.; Joshi, M.S.; Dharmadhikari, A.S.; Jaiswal, S.V. Mental state and emotion detection from musically stimulated EEG. Brain Inform. 2018, 5, 14. [Google Scholar] [CrossRef]
- Hawkins, K.A.; Fox, E.J.; Daly, J.J.; Rose, D.K.; Christou, E.A.; McGuirk, T.E.; Otzel, D.M.; Butera, K.A.; Chatterjee, S.A.; Clark, D.J. Prefrontal over-activation during walking in people with mobility deficits: Interpretation and functional implications. Hum. Mov. Sci. 2018, 59, 46–55. [Google Scholar] [CrossRef]
- Mizuguchi, N.; Kanosue, K. Changes in brain activity during action observation and motor imagery: Their relationship with motor learning. Prog. Brain Res. 2017, 234, 189–204. [Google Scholar]
- Thobois, S.; Dominey, P.F.; Decety, J.; Pollak, P.; Gregoire, M.C.; Le Bars, D.; Broussolle, E. Motor imagery in normal subjects and in asymmetrical Parkinson’s disease: A PET study. Neurology 2000, 55, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kirino, E. Dynamic reconfiguration of the supplementary motor area network during imagined music performance. Front. Hum. Neurosci. 2017, 11, 606. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Yoon, J.G.; Lee, S.W. Predicting motor imagery performance from resting-state EEG using dynamic causal modeling. Front. Hum. Neurosci. 2020, 14, 321. [Google Scholar] [CrossRef]
- Mehler, D.M.; Williams, A.N.; Krause, F.; Lührs, M.; Wise, R.G.; Turner, D.L.; Linden, D.E.J.; Whittaker, J.R. The BOLD response in primary motor cortex and supplementary motor area during kinesthetic motor imagery based graded fMRI neurofeedback. Neuroimage 2019, 184, 36–44. [Google Scholar] [CrossRef]
- Savaki, H.E.; Raos, V. Action perception and motor imagery: Mental practice of action. Prog. Neurobiol. 2019, 175, 107–125. [Google Scholar] [CrossRef]
- Tayah, T.; Savard, M.; Desbiens, R.; Nguyen, D.K. Ictal bradycardia and asystole in an adult with a focal left insular lesion. Clin. Neurol. Neurosurg. 2013, 115, 1885–1887. [Google Scholar] [CrossRef]
- Sridharan, D.; Levitin, D.J.; Menon, V. A critical role for the right fronto-insular cortex in switching between central-executive and default-mode networks. Proc. Natl. Acad. Sci. USA 2008, 105, 12569–12574. [Google Scholar] [CrossRef] [Green Version]
- Ghaem, O.; Mellet, E.; Crivello, F.; Tzourio, N.; Mazoyer, B.; Berthoz, A.; Denis, M. Mental navigation along memorized routes activates the hippocampus, precuneus, and insula. Neuroreport 1997, 8, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Brodsky, M.C.; Fray, K.J.; Glasier, C.M. Perinatal cortical and subcortical visual loss: Mechanisms of injury and associated ophthalmologic signs. Ophthalmology 2022, 109, 85–94. [Google Scholar] [CrossRef]
- Jackson, P.L.; Lafleur, M.F.; Malouin, F.; Richards, C.; Doyon, J. Potential role of mental practice using motor imagery in neurologic rehabilitation. Arch. Phys. Med. Rehabil. 2001, 82, 1133–1141. [Google Scholar] [CrossRef]
- Sauvage, C.; Jissendi, P.; Seignan, S.; Manto, M.; Habas, C. Brain areas involved in the control of speed during a motor sequence of the foot: Real movement versus mental imagery. J. Neuroradiol. 2013, 40, 267–280. [Google Scholar] [CrossRef]
- Wagner, J.; Stephan, T.; Kalla, R.; Brückmann, H.; Strupp, M.; Brandt, T.; Jahn, K. Mind the bend: Cerebral activations associated with mental imagery of walking along a curved path. Exp. Brain Res. 2008, 191, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Fleck, M.S.; Daselaar, S.M.; Dobbins, I.G.; Cabeza, R. Role of prefrontal and anterior cingulate regions in decision-making processes shared by memory and nonmemory tasks. Cereb. Cortex 2006, 16, 1623–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huijbers, W.; Pennartz, C.M.; Rubin, D.C.; Daselaar, S.M. Imagery and retrieval of auditory and visual information: Neural correlates of successful and unsuccessful performance. Neuropsychologia 2011, 49, 1730–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Kim, J.; Ryu, K. Comparison the Brain Activation on Choice Reaction Between the Expert and the Novice: A fMRI study. Korean, J. Sport Psychol. 2012, 23, 103–115. [Google Scholar]
- Behrmann, M.; Geng, J.J.; Shomstein, S. Parietal cortex and attention. Curr. Opin. Neurobiol. 2004, 14, 212–217. [Google Scholar] [CrossRef]
- Husain, M.; Nachev, P. Space and the parietal cortex. TiCS 2007, 11, 30–36. [Google Scholar] [CrossRef]
- Thompson, C.; Sabik, M. Allocation of attention in familiar and unfamiliar traffic scenarios. Transp. Res. F Traffic Psychol. Behav. 2018, 55, 188–198. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | EEG Alpha Asymmetry Score | |||
|---|---|---|---|---|
| Clockwise | Counterclockwise | |||
| M (SD) | Range | M (SD) | Range | |
| Log Fp1 | 5.09 (1.97) | 1.88–6.62 | 5.29 (1.53) | −0.64–6.85 |
| Log Fp2 | 5.20 (1.99) | 0.18–6.62 | 5.32 (1.76) | −1.56–7.00 |
| Log Fp2-Fp1 | 0.11 (0.19) | −0.18–0.66 | 0.27 (0.27) | −0.91–0.31 |
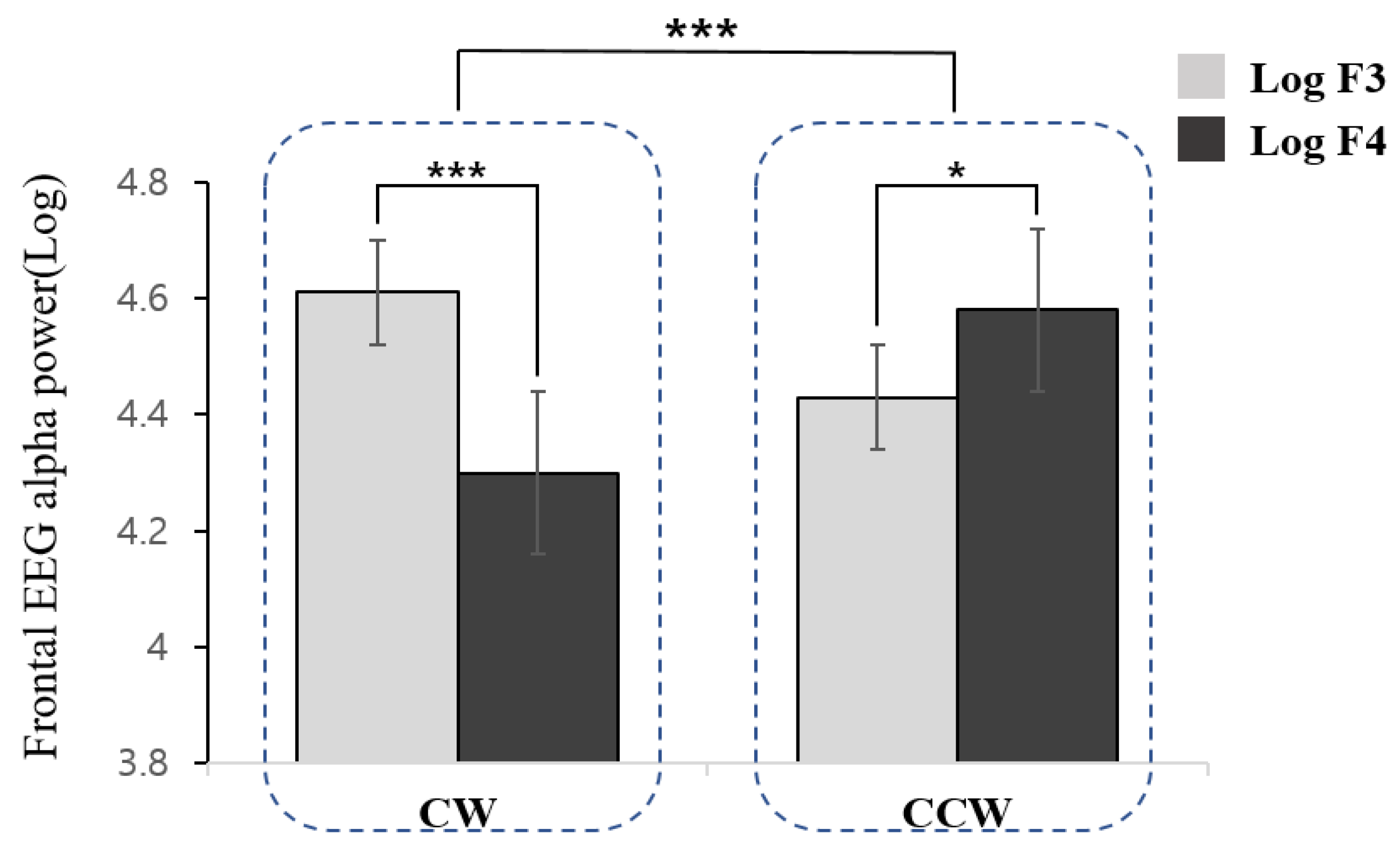
| Log F3 | 4.60 (1.66) | 2.14–4.92 | 4.43 (1.31) | 1.40–5.53 |
| Log F4 | 4.30 (1.60) | 2.27–4.68 | 4.57 (1.25) | 1.84–5.85 |
| Log F4-F3 | −0.31 (0.20) | −0.74–0.56 | 0.14 (0.24) | −0.21–0.61 |
| Log C3 | 3.48 (1.24) | 1.64–5.97 | 3.06 (1.56) | −1.59–5.32 |
| Log C4 | 3.87 (1.40) | 0.41–6.04 | 4.25 (1.47) | −1.43–5.37 |
| Log C4-C3 | 0.38 (0.90) | −1.23–1.96 | 1.20 (1.73) | −3.21–6.14 |
| Components | Event-Related Potential (ERP) | ||||||
|---|---|---|---|---|---|---|---|
| Clockwise M (SD) n = 33 | Counterclockwise M (SD) n = 33 | ||||||
| Fz | Cz | Pz | Fz | Cz | Pz | ||
| N200 | amplitude (mV) | −22.75 | −52.75 | −62.42 | −30.45 | −57.79 | −65.74 |
| (21.20) | (23.56) | (29.21) | (35.10) | (33.65) | (35.10) | ||
| latency (ms) | 269.65 | 277.99 | 288.65 | 264.83 | 278.75 | 279.32 | |
| (31.93) | (31.89) | (32.01) | (37.94) | (31.89) | (39.83) | ||
| P300 | amplitude (mV) | 35.16 | 40.95 | 49.76 | 38.68 | 42.31 | 50.95 |
| (37.71) | (48.97) | (43.49) | (33.77) | (35.82) | (37.94) | ||
| latency (ms) | 424.70 | 358.96 | 380.69 | 475.11 | 420.87 | 409.46 | |
| (131.62) | (98.23) | (86.71) | (145.01) | (100.20) | (111.07) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.; Kim, J.; Kwon, S. Neuropsychological Evidence Underlying Counterclockwise Bias in Running: Electroencephalography and Functional Magnetic Resonance Imaging Studies of Motor Imagery. Behav. Sci. 2023, 13, 173. https://doi.org/10.3390/bs13020173
Kim T, Kim J, Kwon S. Neuropsychological Evidence Underlying Counterclockwise Bias in Running: Electroencephalography and Functional Magnetic Resonance Imaging Studies of Motor Imagery. Behavioral Sciences. 2023; 13(2):173. https://doi.org/10.3390/bs13020173
Chicago/Turabian StyleKim, Teri, Jingu Kim, and Sechang Kwon. 2023. "Neuropsychological Evidence Underlying Counterclockwise Bias in Running: Electroencephalography and Functional Magnetic Resonance Imaging Studies of Motor Imagery" Behavioral Sciences 13, no. 2: 173. https://doi.org/10.3390/bs13020173