Induction of Proteasome Subunit Low Molecular Weight Protein (LMP)-2 Is Required to Induce Active Remodeling in Adult Rat Ventricular Cardiomyocytes

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Isolation and Cultivation
2.2. Spontaneously Hypertensive Rats
2.3. RT-PCR
2.4. Western Blot
2.5. Statistics
3. Results
3.1. Kinetics of Sarcomere Degradation
3.2. Cell Shape of Mono and Binucleated Cells and Adaptation to Culture Conditions
3.3. Molecular Adaptation of ARVC during Cultivation
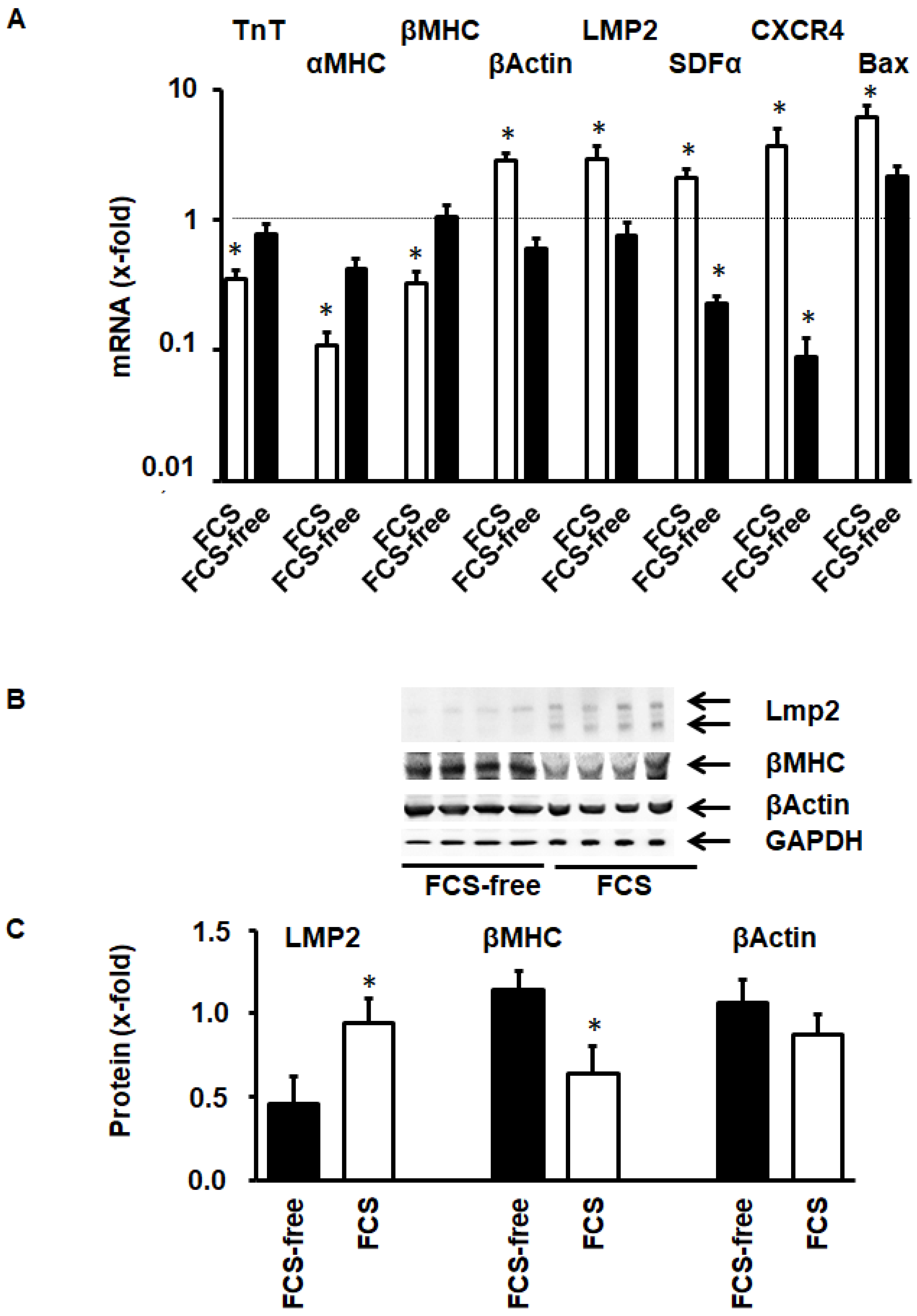
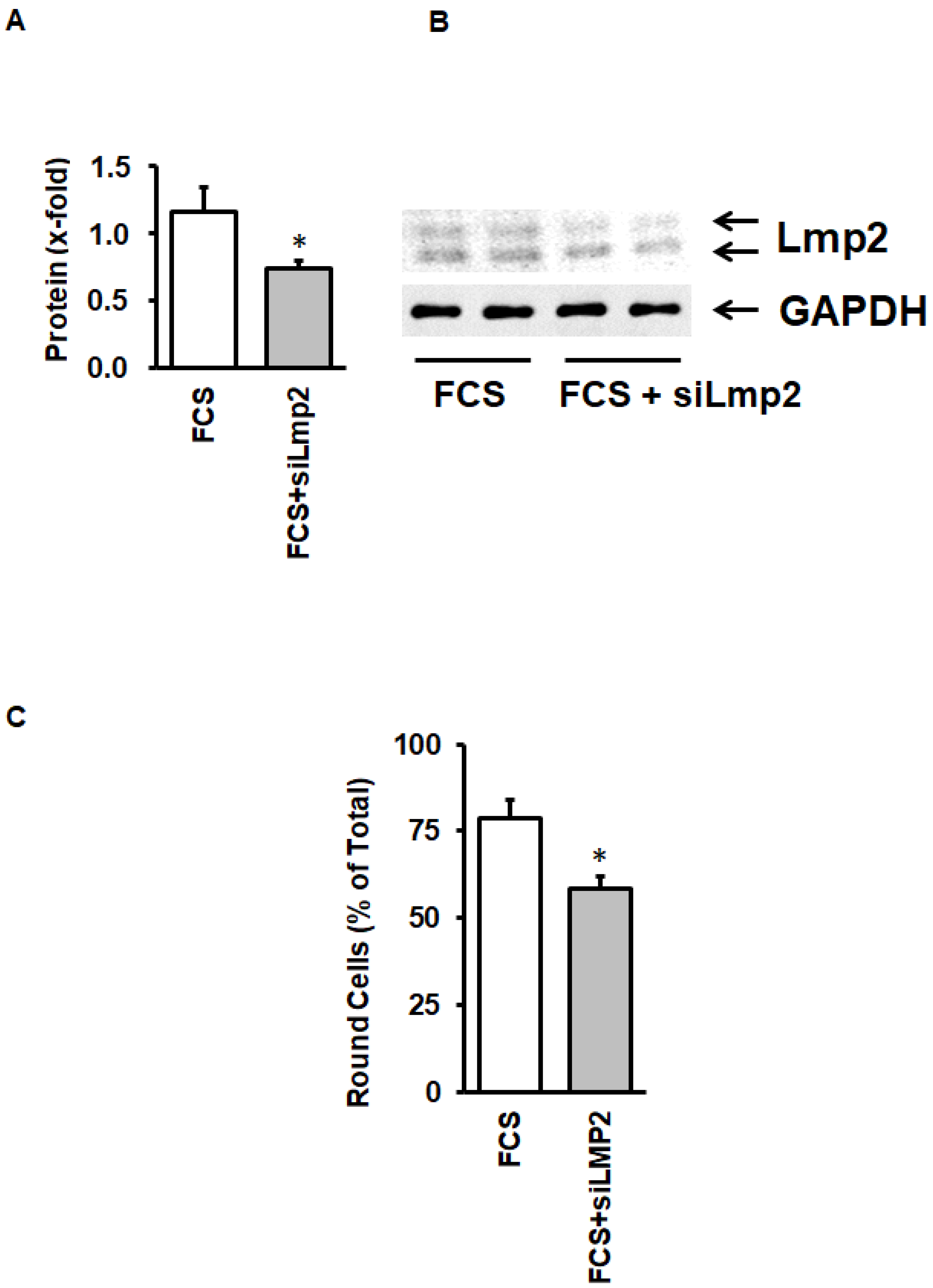
3.4. Activation of LMP-2 and Contribution to Cellular Adaptation to Culture Conditions
3.5. Induction of LMP-2 during the Adaptation of the Left Ventricle to Pressure Overload
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Li, T.-S.; Lee, S.-T.; Wawrowsky, K.A.; Cheng, K.; Galang, G.; Malliaras, K.; Roselle Abraham, M.; Wang, C.; Marban, E. Dedifferentiation and proliferation of mammalian cardiomyocytes. PLoS ONE 2010, 5, e12559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nippert, F.; Schreckenberg, R.; Hess, A.; Weber, M.; Schlüter, K.-D. The effects of swiprosin-1 on the formation of pseudopodia-like structures and β-adrenoceptor coupling in cultured adult rat ventricular cardiomyocytes. PLoS ONE 2016, 10, e0167655. [Google Scholar] [CrossRef] [PubMed]
- Kubin, T.; Pöling, J.; Kostin, S.; Gajawada, P.; Hein, S.; Rees, W.; Wietelmann, A.; Tanaka, M.; Lörchner, H.; Schimanski, S.; et al. Oncostatin M is a major mediator of cardiomyocyte differentiation and remodeling. Cell Stem Cell 2011, 9, 420–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, A.N.; Gay, M.S.; Zhang, L. Binucleation of cardiomyocytes: The transition from a proliferative to a terminally differentiated state. Drug Disc. Today 2014, 19, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Brodsky, W.Y.; Arefyeva, A.M.; Uryvaeva, I.V. Mitotic polyploidization of mouse heart myocytes during the first postnatal week. Cell Tissue Res. 1980, 210, 133–144. [Google Scholar] [CrossRef]
- Senyo, S.E.; Steinhauser, M.L.; Pizzimenti, C.L.; Yang, V.K.; Cai, L.; Wang, M.; Wu, T.D.; Guerguin-Kern, J.L.; Lechene, C.P.; Lee, R.T. Mammalian heart renewal by pre-existing cardiomyocytes. Nature 2013, 493, 433–437. [Google Scholar] [CrossRef] [Green Version]
- Brodsky, W.Y.; Tsirekidze, N.N.; Arefyeva, A.M. Mitotic-cyclic and cyclic-independent growth of cardiomyocytes. J. Mol. Cell. Cardiol. 1985, 17, 445–455. [Google Scholar] [CrossRef]
- Wohlschlaeger, J.; Levkau, B.; Brockhoff, G.; Schmitz, K.J.; von Winterfeld, M.; Takeda, A.; Takeda, N.; Stypmann, J.; Vahlhaus, C.; Schmid, C.; et al. Hemodynamic support by left ventricular assist device reduces cardiomyocytes DNA content in the failing human heart. Circulation 2010, 121, 989–996. [Google Scholar] [CrossRef]
- Engelmann, G.L.; Vitullo, J.C.; Gerrity, R.G. Age-related changes in ploidy levels and biochemical parameters in cardiac myocytes isolated from spontaneously hypertensive rats. Circ. Res. 1986, 58, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Kellermann, S.; Moore, J.A.; Zierhut, W.; Zimmer, H.-G.; Campbell, J.; Gerdes, A.M. Nuclear DNA content and nucleation patterns in rat cardiac myocytes from different models of cardiac hypertrophy. J. Mol. Cell. Cardiol. 1992, 24, 497–505. [Google Scholar] [CrossRef]
- Brodsky, W.Y.; Carlson, B.M.; Arefieva, A.M.; Vacilieva, I.A. Polyploidization of transplanted cardiac myocytes. Cell Differ. Dev. 1988, 25, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, B.; Maddox, A.S. Cytokinesis, ploidy and aneuploidy. J. Pathol. 2012, 226, 338–351. [Google Scholar] [CrossRef] [PubMed]
- Kajustra, J.; Urbanek, K.; Perl, S.; Hosoda, T.; Zheng, H.; Ogorek, B.; Ferreira-Martins, J.; Goichberg, P.; Rondon-Clavo, C.; Sanada, F.; et al. Cardiomyogensis in the adult human heart. Circ. Res. 2010, 107, 305–315. [Google Scholar]
- Huang, C.-F.; Chen, Y.-C.; Yeh, H.-I.; Chen, S.-A. Mononucleated and binucleated cardiomyocytes in left atrium and pulmonary vein have different electrical activity and calcium dynamics. Prog. Biophys. Mol. Biol. 2012, 108, 64–73. [Google Scholar] [CrossRef]
- Yekelchyk, M.; Guenther, S.; Preussner, J.; Braun, T. Mono- and multi-nucleated ventricular cardiomyocytes constitute a transcriptionally homogenous cell population. Basic Res. Cardiol. 2019, 114, 36. [Google Scholar] [CrossRef] [Green Version]
- Diaz, F.M.; Gilar, M.B.; Sauri, A.R.; Bosh, A.L.; Luna, A. Usefulness of DNA quantification in diagnosis of hypertrophic cardiomyopathies: A preliminary study. Forensic Sci. Int. 2006, 157, 40–45. [Google Scholar] [CrossRef]
- Eppenberger, M.E.; Hauser, I.; Baechi, T.; Schaub, M.C.; Brunner, U.T.; Dechesne, C.A.; Eppenberger, H.M. Immunocytochemical analysis of the regeneration of myofibrils in long-term cultures of adult cardiomyocytes of the rat. Dev. Biol. 1988, 130, 1–15. [Google Scholar] [CrossRef]
- Bugaisky, L.B.; Zak, R. Differentiation and adult rat cardiomyocytes in cell culture. Circ. Res. 1989, 64, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Decker, M.L.; Behnke-Barclay, M.; Cook, M.G.; Lesch, M.; Decker, R.S. Morphometric evaluation of the contractie apparatus in primary cultures of rabbit cardiac myocytes. Circ. Res. 1991, 69, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Joshi-Mukherjee, R.; Dick, I.E.; Liu, T.; O’Rourke, B.; Yue, D.T.; Tung, L. Structural and functional plasticity in long-term cultures of adult ventricular myocytes. J. Mol. Cell. Cardiol. 2013, 65, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Szalay, G.; Meiners, S.; Voigt, A.; Lauber, J.; Spieth, C.; Speer, N.; Sauter, M.; Kuckelkorn, U.; Zell, A.; Klingel, K.; et al. Ongoing coxsackievirus myocarditis is associated with increased formation and activity of myocardial immunoproteasome. Am. J. Pathol. 2006, 168, 1542–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zu, L.; Bedja, D.; Fox-Talbot, K.; Gabrielson, K.L.; van Kaer, L.; Becker, L.C.; Cai, Z.P. Evidence for a role of immunoproteasomes in regulating cardiac muscle mass in diabetic mice. J. Mol. Cell. Cardiol. 2010, 49, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basler, M.; Lauer, C.; Moebius, J.; Weber, R.; Przybylski, M.; Kisselev, A.F.; Tsu, C.; Groettrup, M. Why the structure but not the activity of the immunoproteasome subunit low molecular mass polypedtide 2 rescues antigen presentation. J. Immunol. 2012, 189, 1868–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Lee, C.-M.; Tripathi, S.; Kim, K.-B.; Morgan, E.T. Nitric oxide-dependent CYP2B degradation is potentiated by a cytokine-regulated pathway and utilizes the immunoproteasome subunit LMP2. Biochem. J. 2012, 445, 377–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bøtker, H.E.; Hausenloy, D.; Andreadou, I.; Antonucci, S.; Boengler, K.; Davidson, S.M.; Deshwal, S.; Devaux, Y.; Di Lisa, F.; Di Sante, M.; et al. Practical guidelines for rigor and reproducibility in preclinical and clinical studies on cardioprotection. Basic Res. Cardiol. 2018, 113, 39. [Google Scholar] [CrossRef] [Green Version]
- Nippert, F.; Schreckenberg, R.; Schlüter, K.-D. Isolation and cultivation of adult rat cardiomyocytes. J. Vis. Exp. 2017, e56634. [Google Scholar] [CrossRef]
- Schreckenberg, R.; Horn, A.-M.; da Costa Rebelo, R.M.; Simsekyilmaz, S.; Niemann, B.; Li, L.; Rohrbach, S.; Schlüter, K.D. Effects of 6-months‘ exercise on cardiac function, structure and metabolism in female hypertensive rats–the decisive role of lysyl oxidase and collagen III. Front. Physiol. 2017, 8, 556. [Google Scholar] [CrossRef] [Green Version]
- Anwar, A.; Schlüter, K.-D.; Heger, J.; Piper, H.M.; Euler, G. Enhanced SERCA2A expression improves contractile performance of ventricular cardiomyocytes of rat under adrenergic stimulation. Pflügers Arch. Eur. J. Physiol. 2008, 457, 485–491. [Google Scholar] [CrossRef]
- Ren, X.; Li, Y.; Ma, X.; Zheng, L.; Xu, Y.; Wang, J. Activation of p38/MEF2C pathways by all-trans retinoic acid in cardiac myoblasts. Life Sci. 2007, 81, 89–96. [Google Scholar] [CrossRef]
- Leri, A.; Rota, M.; Pasqualini, F.S.; Goichberg, P.; Anversa, P. Origin of cardiomyocytes in the adult heart. Circ. Res. 2015, 116, 150–166. [Google Scholar] [CrossRef]
- Wenzel, S.; Soltanpour, G.; Schlüter, K.-D. No correlation between p38 MAPK pathway and the contractile dysfunction in diabetic cardiomyocytes. Pflügers Arch. Eur. J. Physiol. 2005, 451, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kubota, J.; Hirayama, J.; Nishina, S.; Yokoi, T.; Asaoka, Y.; Seo, J.; Shimizu, N.; Kajiho, H.; Watanabe, T.; et al. p38 Mitogen-activated protein kinase controls a switch between cardiomyocytes and neuronal commitment of murine embraonic stem cells by activating myocyte enhancer factor 2C-dependent bone morphogenetic protein 2 transcription. Stem Cells Develop. 2010, 19, 1723–1734. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Torres, F.; Martinez-Fernandez, S.; Zuluaga, S.; Nebreda, A.; Porras, A.; Aranega, A.E.; Navarro, F. A role for p38α mitogen-activated protein kinase in embryonic cardiac differentiation. FEBS Lett. 2008, 582, 1025–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, H.; Li, J.; Zhang, H.; Ma, W.; Wie, N.; Liu, J.; Wang, X. COP9 signalosome controls the degradation of cytosolic misfolded proteins and protects against cardiac proteotoxicity. Circ. Res. 2015, 117, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Bitan, G.; Schrader, T.; Klärner, F.-G.; Osinska, H.; Robbins, J. Inhibition of mutant αB crystallin-induced protein aggreagtion by a molecular tweezer. J. Am. Heart Assoc. 2017, 6, e006182. [Google Scholar] [CrossRef]
- Campos, J.C.; Fernandes, T.; Bechara, L.R.G.; da Paixao, N.A.; Brum, P.C.; de Oliveira, E.M.; Ferreira, J.C. Increased clearance of reactive aldehydes and damaged proteins in hypertension-induced compensated cardiac hypertrophy: Impact of exercise training. Oxidative Med. Cell. Longev. 2015, 2015, 464195. [Google Scholar] [CrossRef] [Green Version]
- Glembotsky, C.C. Breaking down the COP9 signalsome in the heart. Circ. Res. 2015, 117, 914–916. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Horak, K.M.; Su, H.; Sanbe, A.; Robbins, J.; Wang, X. Enhancement of proteosomal function protects against cardiac proteinopathy and ischemia/reperfusion injury in mice. J. Clin. Invest. 2011, 121, 3689–3700. [Google Scholar] [CrossRef]
- Gilda, J.E.; Gomes, A.V. Proteasome dysfunction in cardiomyopathies. J. Physiol. 2017, 595, 4051–4071. [Google Scholar] [CrossRef]
- Ebstein, F.; Kloetzel, P.-M.; Krüger, E.; Seifert, U. Emerging roles of immunoproteasome beyond MHC class I antigen processing. Cell Mol. Life Sci. 2012, 69, 2543–2558. [Google Scholar] [CrossRef]
- Gerdes, A.M.; Morales, M.C.; Handa, V.; Moore, J.A.; Alvarez, M.R. Nuclear size and DNA content in rat cardiac myocytes during growth, maturation and aging. J. Mol. Cell. Cardiol. 1991, 23, 833–839. [Google Scholar] [CrossRef]
- Hein, S.; Aron, E.; Kostin, S.; Schonburg, M.; Elsasser, A.; Polyakova, V.; Bauer, E.P.; Klovekorn, W.P.; Schaper, J. Progression from compensated hypertrophy to failure in the pressure-overloaded human heart: Structural deterioration and compensatory mechanism. Circulation 2003, 107, 984–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stansfield, W.E.; Tang, R.H.; Moss, N.C.; Baldwin, A.S.; Willis, M.S.; Selzman, C.H. Proteasome inhibition promotes regression of left ventricular hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H645–H650. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cheng, Y.; Dou, Y.; Goonesekara, C.; David, J.-P.; Steele, D.F.; Huang, C.; Fedida, D. Trafficking of an endogenous potassium channel in adult ventricular myocytes. Am. J. Physiol. Cell. Physiol. 2012, 303, C963–C976. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petersen, A.; Kutsche, H.S.; Nippert, F.; Schreckenberg, R.; Schulz, R.; Schlüter, K.-D. Induction of Proteasome Subunit Low Molecular Weight Protein (LMP)-2 Is Required to Induce Active Remodeling in Adult Rat Ventricular Cardiomyocytes. Med. Sci. 2020, 8, 21. https://doi.org/10.3390/medsci8020021
Petersen A, Kutsche HS, Nippert F, Schreckenberg R, Schulz R, Schlüter K-D. Induction of Proteasome Subunit Low Molecular Weight Protein (LMP)-2 Is Required to Induce Active Remodeling in Adult Rat Ventricular Cardiomyocytes. Medical Sciences. 2020; 8(2):21. https://doi.org/10.3390/medsci8020021
Chicago/Turabian StylePetersen, Antonia, Hanna Sarah Kutsche, Franziska Nippert, Rolf Schreckenberg, Rainer Schulz, and Klaus-Dieter Schlüter. 2020. "Induction of Proteasome Subunit Low Molecular Weight Protein (LMP)-2 Is Required to Induce Active Remodeling in Adult Rat Ventricular Cardiomyocytes" Medical Sciences 8, no. 2: 21. https://doi.org/10.3390/medsci8020021