1. Introduction
Taste is well-known biological descriptor for sweet, bitter, sour, salty receptors and ion channels, which plays a critical role in the life and nutritional status of chickens and other organisms. Bitter taste perception provides animals with critical protection against the ingestion of poisonous and harmful toxic compounds [
1]. Taste signals have been associated to food avoidance and recognition, as well as feed or liquid intake in different species of animals [
2,
3,
4]. With the simultaneous inflation in the cost of animal feed and higher standards of livestock products, people endeavor to discover novel feed additives and effective alternatives to traditional antibiotics [
5,
6]. Efforts have been made to extract incredible numbers of potential additives from natural plants, and they often display bitter taste. However, bitter taste receptors are part of a superfamily which includes more than twenty members. This makes very difficult to do functional studies for each bitter taste receptor. In addition, bitter taste receptors expressions are not limited to taste buds but also exist in extra-gustatory organs, so it is critical to determine their extra-gustatory functions. Several studies have already revealed that bitter taste receptors exert a variety of functions in different cells and tissues [
7,
8]. Chickens have only three bitter taste receptors: ggTas2r1, ggTas2r2 and ggTas2r7 [
9]. Due to their low number of bitter taste receptors, chickens are a good minimalistic model for understanding the functions of bitter taste receptors in non-gustatory tissues [
9]. Denatonium benzoate as a bitter taste receptor stimulus can bind to the bitter taste receptors to activate bitter taste signaling. Furthermore, it has been reported that this receptor family plays a critical role in the heart [
10], thyroid [
11], and gastrointestinal muscle [
12]. However, their roles in the kidney has never been determined. Therefore, our present project aimed to examine the roles and underlying mechanisms of bitter taste transduction signaling associated with mRNA expression patterns of bitter taste receptors, apoptosis-, autophagy- and antioxidant-related genes in the hearts and kidneys of chickens.
Cysteine aspartate-specific proteases (CASPs or caspases) serve as intrinsic initiators of apoptosis by cleaving substrates at aspartate residues [
13]. In mammals and humans the caspase protein family currently consists of 13 and 11 isoenzymes, respectively [
14]. The function of caspases is closely associated to the initiation and execution of apoptosis, with caspases categorized as either initiator or effector caspases. CASP2, CASP8, CASP9 and CASP10 are initiator caspases, and CASP3, CASP6 and CASP7 are effector caspases [
15]. Known as interleukin 1β-converting enzyme, CASP1 plays an important role in both apoptosis and inflammation [
16]. Apoptosis is regulated by stepwise activation of caspases for the processing or cleaving of other caspases [
17].
Programmed cell death or apoptosis has a vital role in various biological events. This is a process of single cell death controlled by the activation of specific genes, the elimination of unwanted or damaged cells by apoptosis is an indispensable action that occurs via several mechanisms which maintain cellular homeostasis, and normal regulation of the immune system [
18]. An earlier study showed that denatonium benzoate enhanced intracellular Ca
2+, damaged mitochondria and induced apoptosis in airway epithelial cells, respectively [
19]. Programmed cell death is activated by intracellular stresses and developmental cues. Well-known representative intrinsic regulators, the extended BCL2 family proteins, play crucial roles in cell death regulation and are able to regulate several cell death mechanisms, including apoptosis, necrosis and autophagy [
20,
21,
22]. Caspase-independent mechanisms lead to the release of apoptosis-inducing factor (AIF) from the mitochondria, inducing large-scale DNA fragmentation in several cell types including heart and kidney cells to induce apoptosis caused by AIF [
22]. BCL2 family proteins contain at least one of four BCL2 homology (BH) domains (i.e., BH1, BH2, BH3 or BH4), and the number of BH domains included in proteins is associated with their apoptotic functions [
23].
Autophagy is the pathway involved in forming an organelle called an autophagosome. This pathway moves something from the cytoplasm of the cell into the lysosome for degradation. The term, which derives from the Greek words ‘auto’ meaning self and ‘phagos’ meaning to eat, is defined as a catabolic pathway involving the degradation of cellular components via the lysosomal machinery [
24,
25]. Autophagy is the natural process by which the cells can clear out damaged mitochondria, recycle proteins, and get rid of intracellular pathogens. However, all organisms need a balance of autophagy with anabolic processes. There are about 30+ different proteins involved in the formation of the autophagosome. Researchers are still actively figuring out how all of the bits and pieces of the process go together, but recent genetic studies have shed a lot of light on the pathway. A family of genes known as the autophagy-related genes, whose abbreviations start with A/TG, code for several of the proteins integral to autophagy. Several of these genes have variants that have been studied in reference to pathogen susceptibility, autoimmune diseases, cancer, and sepsis [
26,
27,
28].
There is a complex system containing natural enzymatic and non-enzymatic antioxidants that protect the body from oxidative damage. Briefly, the antioxidant enzymes (SOD, MnSOD, CAT, GSH, and GSH-Px) appear to be the first line of defense during oxidative stress, and exert beneficial effects preventing oxidative damage in poultry raising [
29,
30,
31]. Catalase (CAT) participates in defense mechanisms against oxidative stress by converting H
2O
2 into water and molecular oxygen [
32]. The antioxidant glutathione peroxidase enzymes (GPX) are implicated in the protection of cells against oxidative damage by reducing H
2O
2 and other organic peroxidases to water with reduced glutathione [
32].
Denatonium benzoate (DB), one of the most bitter-tasting substances known, is described as extremely unpleasant at a lower amount and can cause perceptible bitterness [
33]. DB has been demonstrated extensively as a bitter taste agonist and used to activate bitter taste receptors in many cell types, including taste cells [
34]. DB is intensely bitter and non-toxic, and can be detected by human taste receptors [
35]. For these reasons DB has been broadly added to liquid detergents, cosmetics, plastic toys and personal care products to avoid the consumption of harmful substances [
36]. Aside from its strong bitter taste, DB also exerts biologic effects on various physiological systems in different organisms. Exposure to DB quickly suppressed ongoing intake and delayed gastric emptying in rodents [
37,
38].
However, there is limited knowledge about the biologic effects of DB and its potential molecular mechanisms in chicken heart and kidneys and no studies have been conducted to date to investigate the relationship between bitter receptors, signaling effectors, and apoptosis-, autophagy- and antioxidant- related genes in the chicken heart and kidney. Therefore, the objective of this study was to investigate the mRNA expressions of bitter taste and its downstream signaling effectors, apoptosis-, autophagy-, and antioxidant-related genes and transduction signaling pathways in chicken heart and kidney to DB dietary exposure in a dose-response manner. The results were confirmed by RT-PCR, qRT-PCR, haemotoxylin and eosin, TUNEL assay, correlation of apoptosis, autophagy and bitter taste receptors and its associated downstream signaling pathway, complete amino acid sequence alignments of related genes, selected gene heat mapping and the potential molecular mechanisms of action of dietary exposure to denatonium benzoate on the heart and kidney cells of Chinese Fast Yellow chickens were determined.
4. Discussion
In the present study, we investigated the biological effects of denatonium benzoate (DB) on growth performance, mRNA expressions of bitter taste receptors, its downstream signaling effectors genes and related pathway, apoptosis, autophagy, antioxidant related genes, histological changes and correlations among genes expressions in the heart and kidney of Chinese Fast Yellow chickens on both day-07 and day-28 of the experiments using RT, qRT-PCR, Hematoxylin and Eosin and TUNNEL assays. We found that DB induced apoptosis, autophagy and increased the expressions of antioxidant-related genes in the heart and kidney of the chickens. However, the expressions of bitter taste receptors genes and its downstream signaling effectors were significantly higher on day-07 compared to day-28 for different DB doses, but the High-dose DB had more potential effects on apoptosis, autophagy, antioxidant, bitter taste receptors and its downstream signaling effectors gene expression than other doses, which significantly induced apoptosis and autophagy in the heart and kidney of chicken on both day-07 and day-28. Remarkably, we also found that bitter taste receptors and the associated signaling effectors, apoptosis, autophagy and antioxidant gene expressions were dependent in a dose-response manner. These findings suggest that the bitter taste receptors have a potential role among the extra-gustatory organs of the chicken, and high-dose DB causes severe necrosis via apoptosis and may result in autophagy in chicken heart and kidneys, while these symptoms were obviously observed on day-07, which proves that the chicken were more sensitive to DB exposure at the beginning of the experiments and later they adapted accordingly.
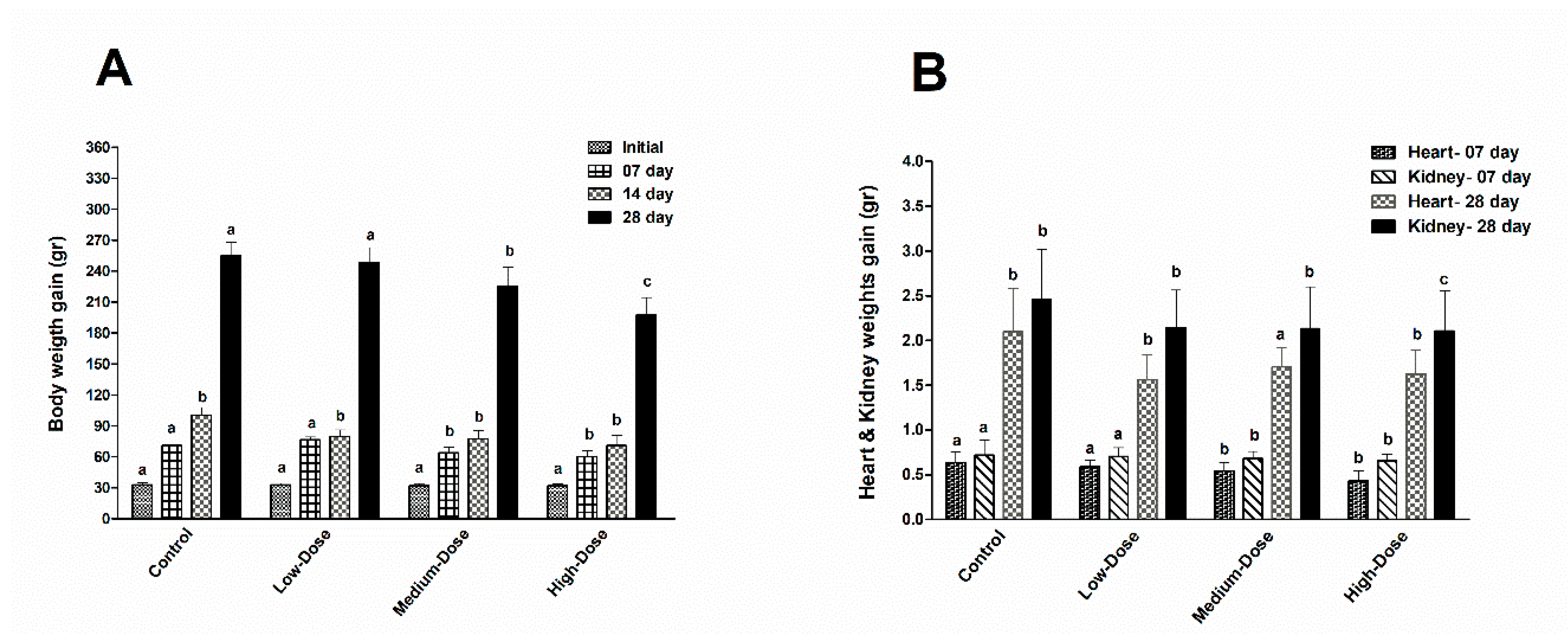
DB (485–740 mg/kg) exhibited a low toxicity rate in acute oral LD
50 tests in rats and rabbits, while chronic toxicity studies have indicated that gavage of 16 mg/kg/day resulted in no compound-related toxicity in monkeys and rats [
43,
44]. In our current study, the average exposure amount of DB in the Low-dose (5 mg/kg), Medium-dose (20 mg/kg) and High-dose (100 mg/kg) groups was calculated on a daily basis for the feed and was less than the doses above. Therefore, DB dietary exposure for 28 days significantly reduced the growth performance and organ (heart and kidney) weights of the chickens. In agreement with our results, four weeks of treatment with bitter agonists like DB or quinine resulted in decreases in body weight gain associated with decreased feed intake [
45]. Moreover, DB has been shown to influence ongoing interdigestive behavior, food intake and gut peptide secretion in healthy volunteers and DB may be able to suppress the contraction of smooth muscles, which inevitably affects the nutrient integration, palatability, digestibility and impairs body weight gain [
33,
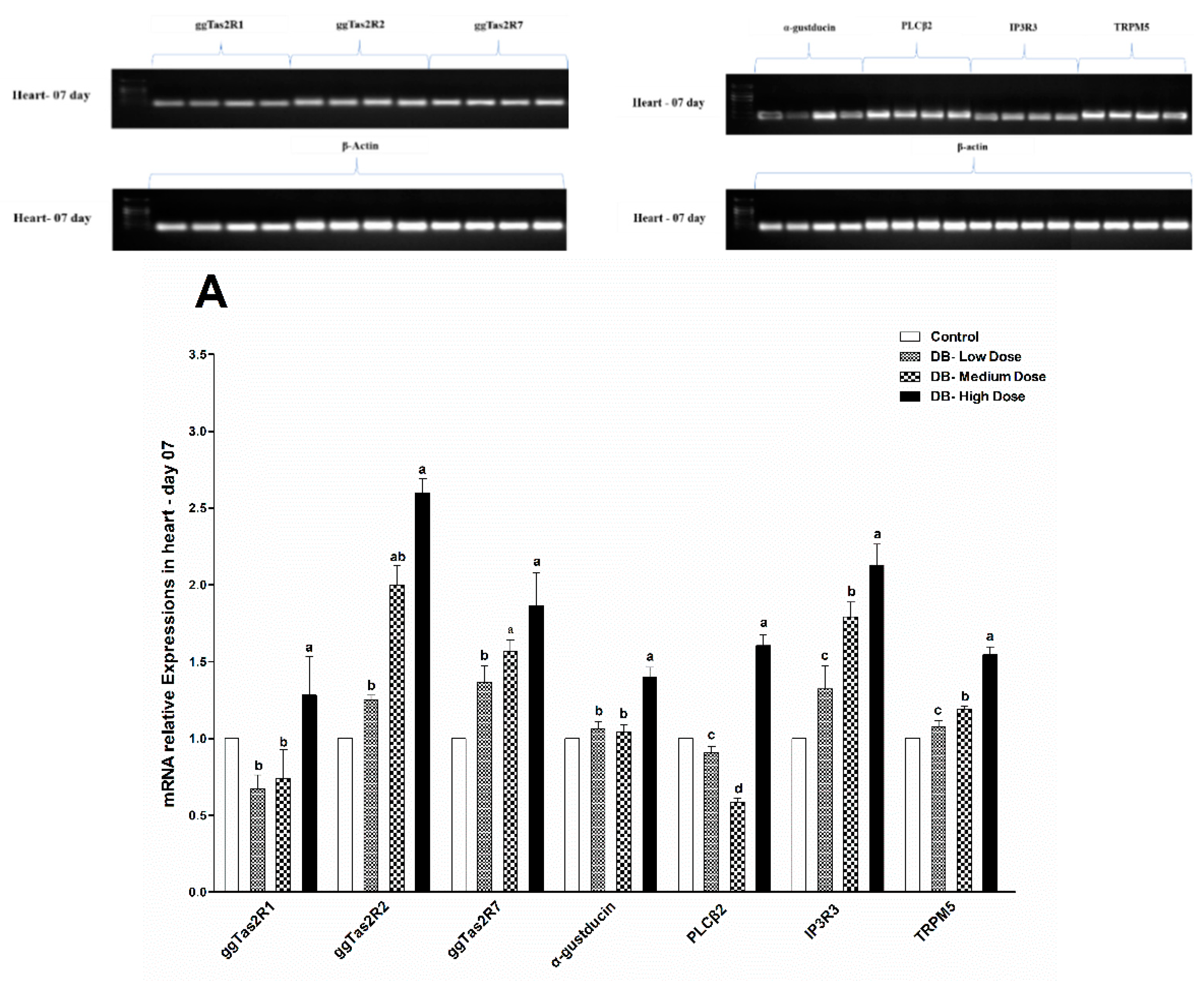
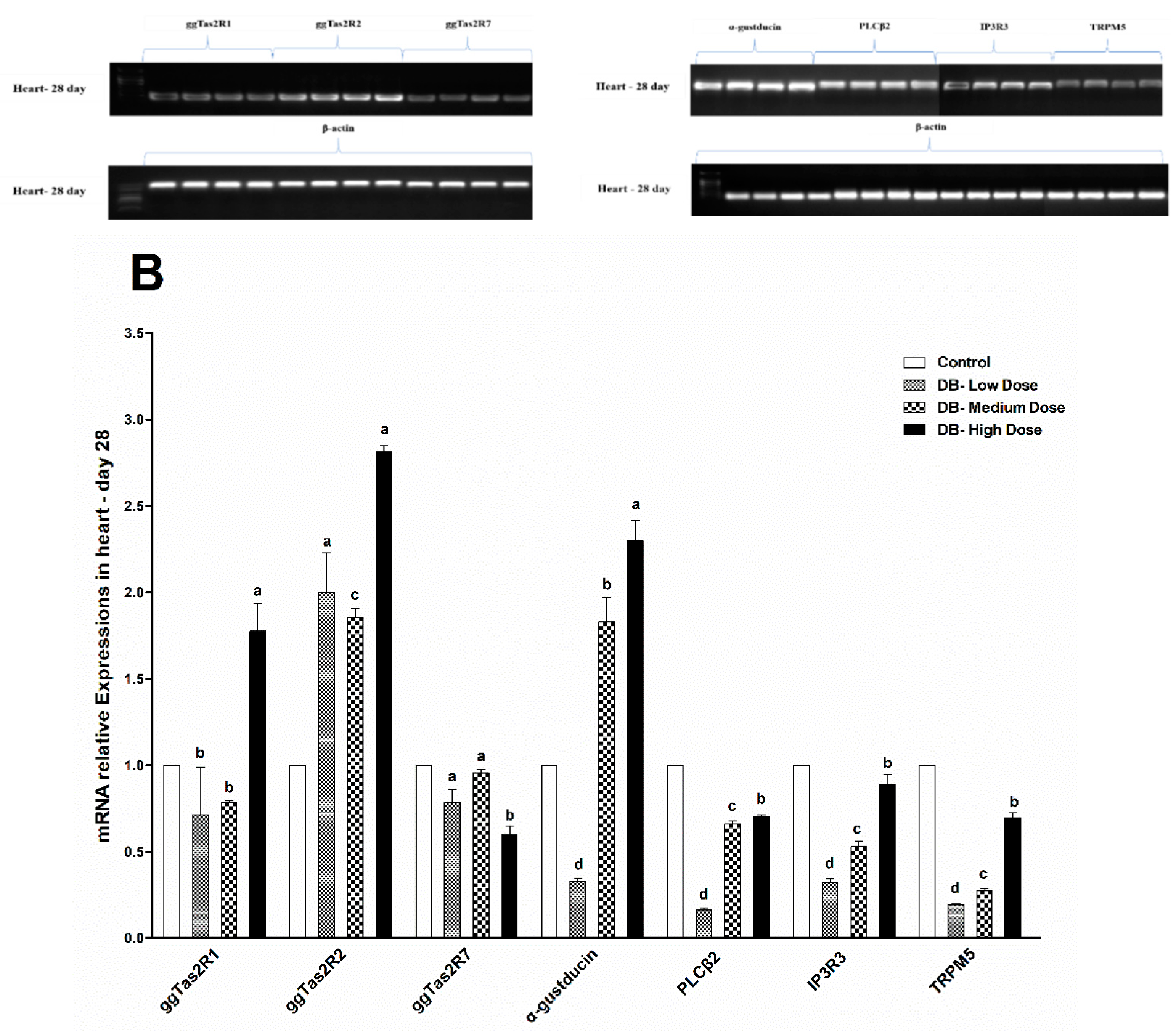
46]. Interestingly, our results indicated that at day-07 and day-28 the ggTas2r2 expressions were higher among three tested bitter taste receptors, and separately, the higher expressions were found in the DB High-dose (100 mg/kg) group, which indicate that the expressions were dose-dependent. Overall, the expressions were significantly decreased on day-28 in contrast do day-07. This finding indicates that chicken sensitivity to DB decreased consequently.
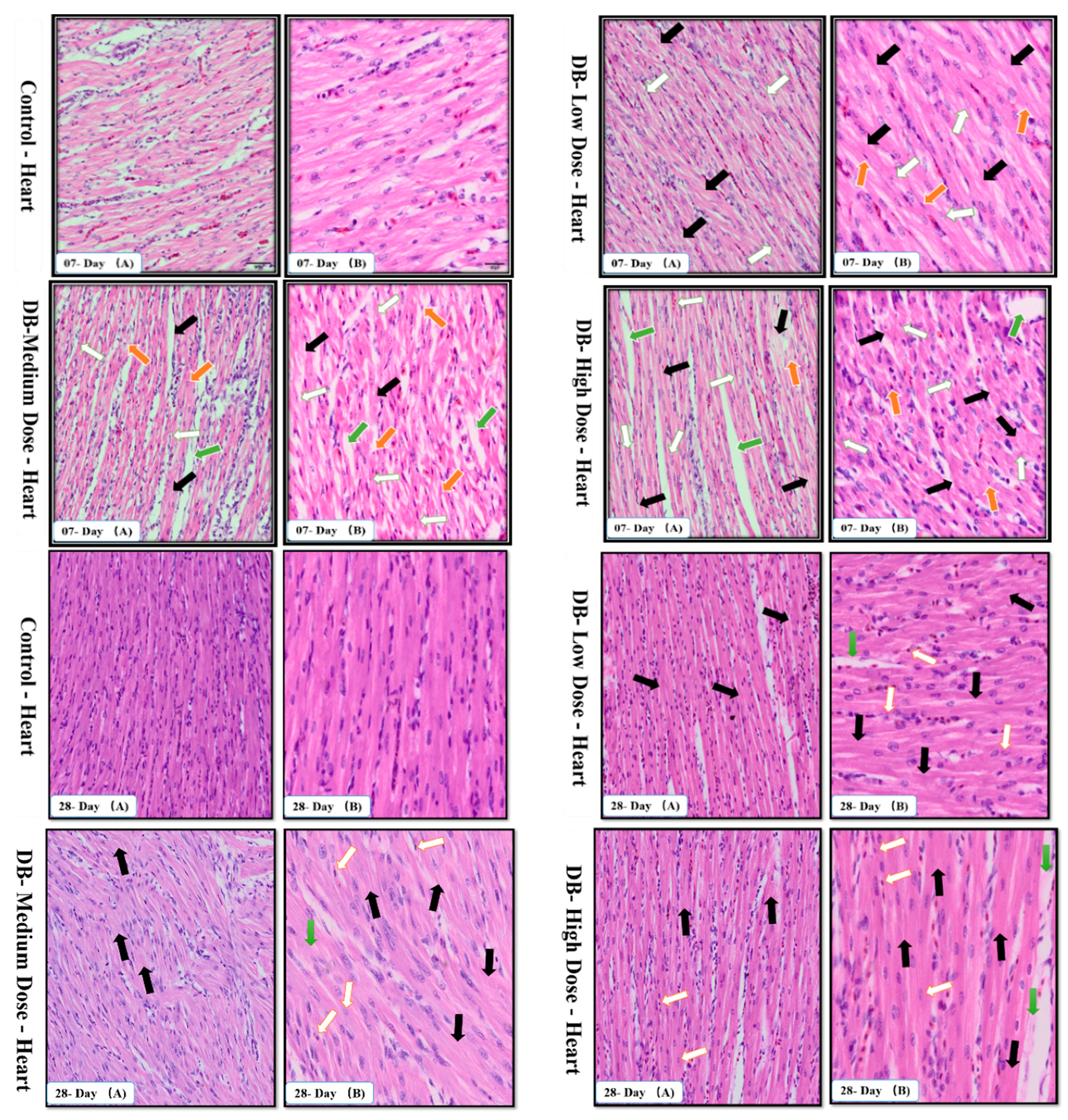
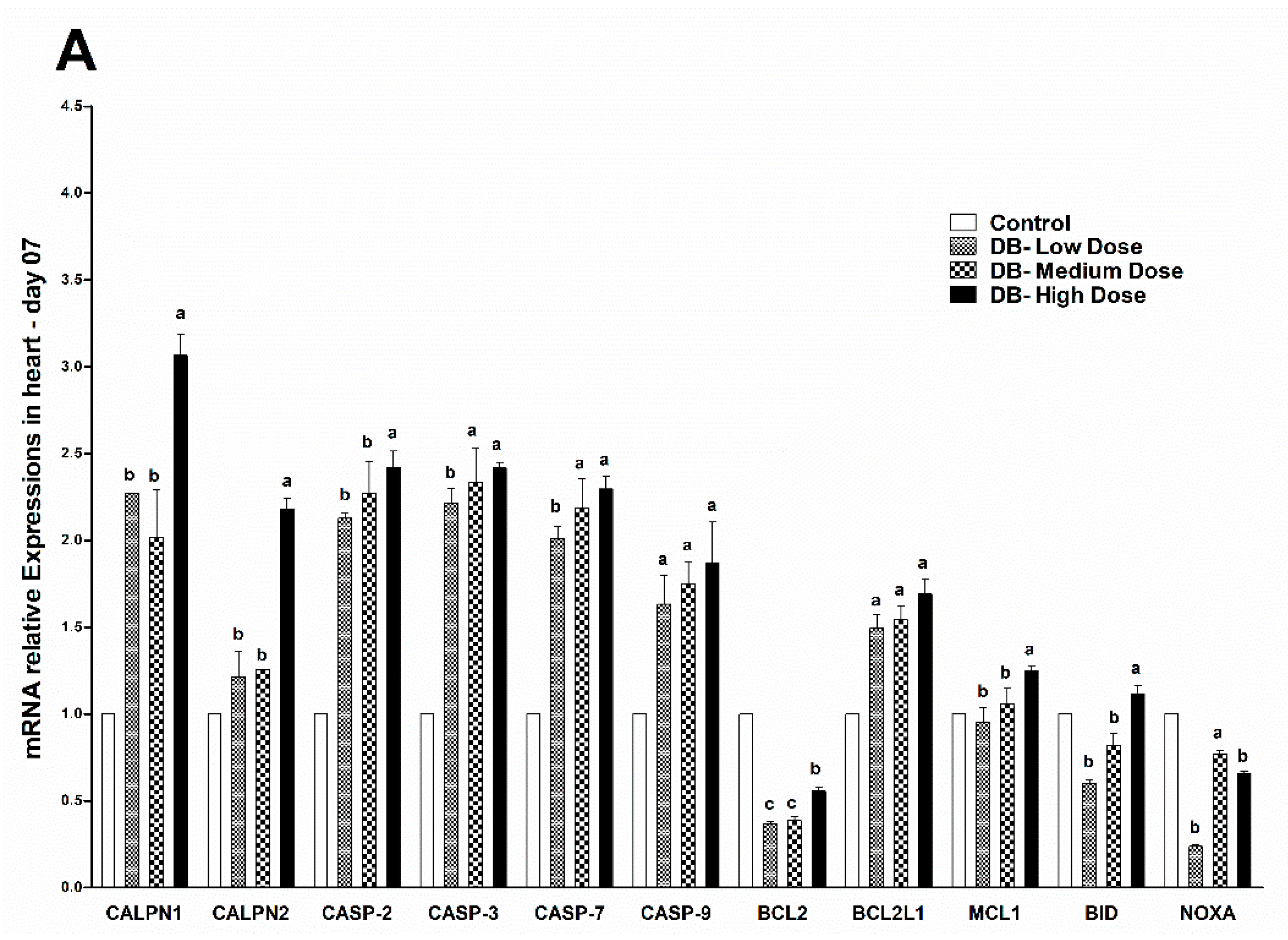
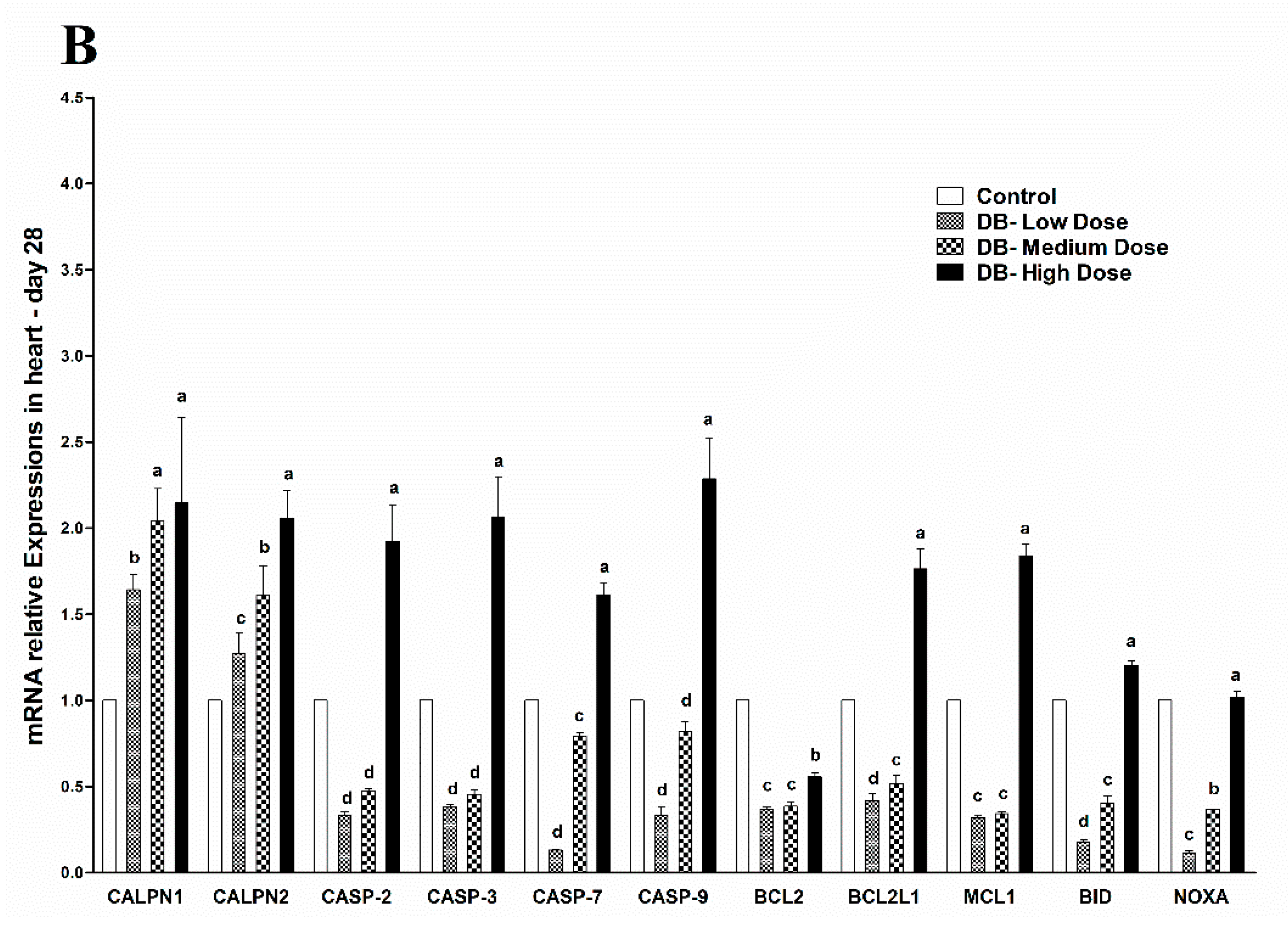
The avian circulatory system is the main transport system of the body. It is the means by which nutrients, enzymes and other important needs for the proper functioning of body systems, organs, tissues and cells as well as body defense components are transported to where they are required. The heart is the most significant and vital organ of the avian circulatory system and its main function is blood supply/pumping of the blood [
47]. To our knowledge this is the first time the potential mechanism underlying the heart pathological changes caused by different doses of DB has been determined. We performed haemotoxylin and eosin staining as well as TUNNEL assay examinations on day-07. In the haemotoxylin and eosin staining expreiments, we found particular pathological changes which alter cell necrosis, apoptosis, pyknotic cells, and distortion of morphological characteristics of the cell due to toxic effect of denatonium on the heart of chicken. Interestingly, these changes were more due to medium and high dose DB exposure. However, on day-28, we observed severe necrosis which caused apoptosis, autophagy and some shrinkage of the fibroblasts which can lead to condensation due to the effect of a high dose of BD (
Figure 2A,B).
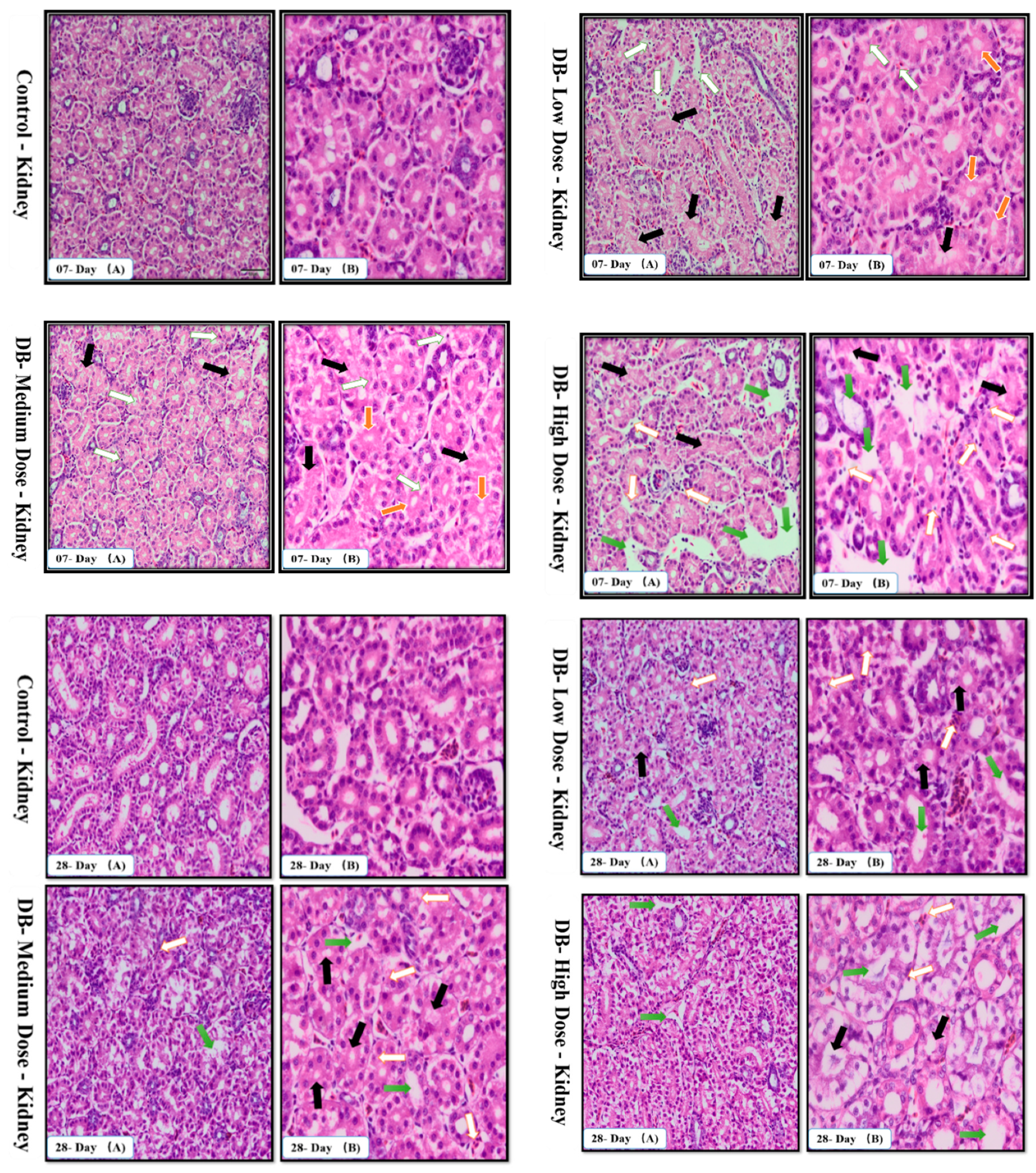
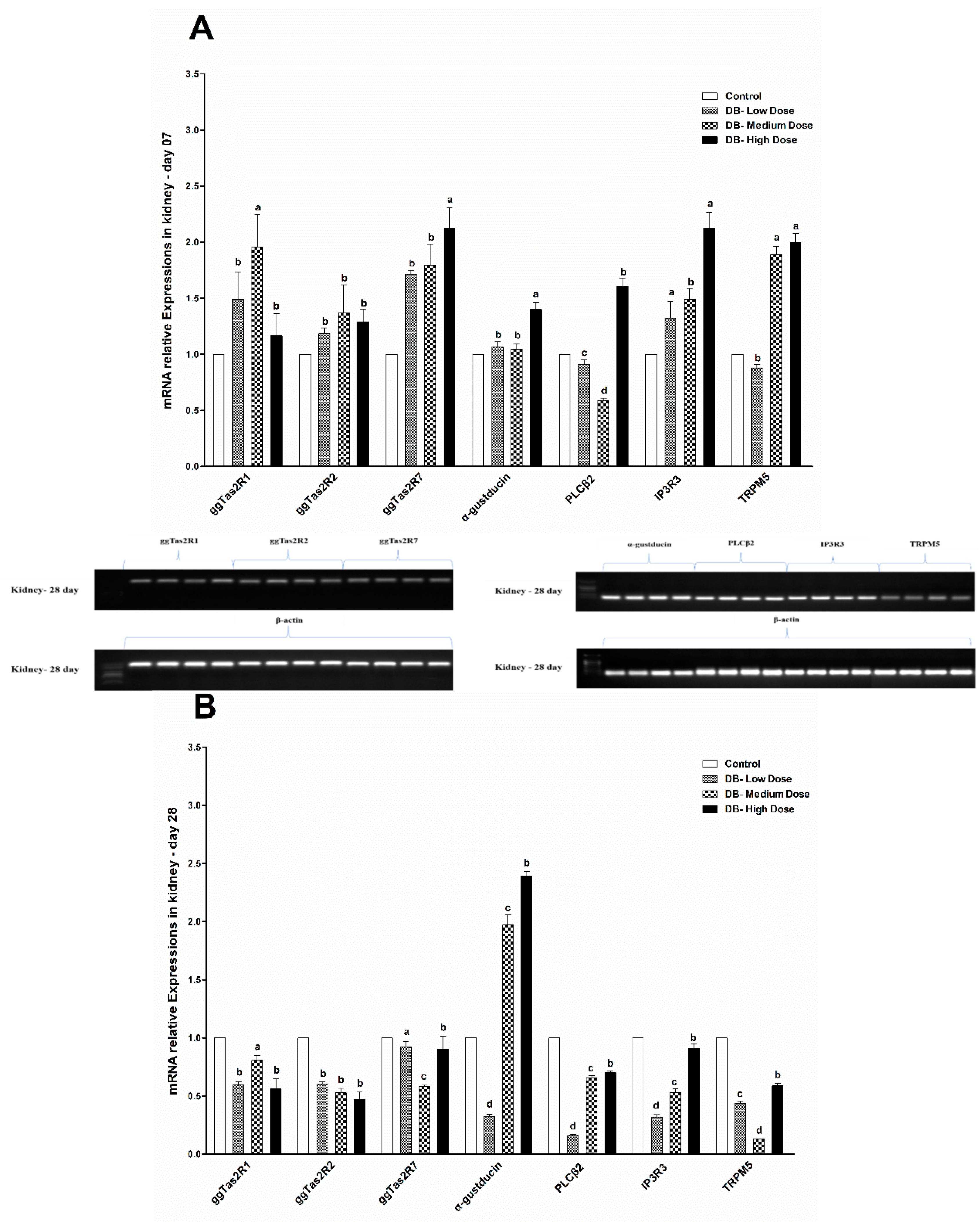
The urinary system is very complex because of its function. The kidneys maintain the water balance by removing excess water from the blood stream. Additionally, the kidneys maintain the electrolyte balance, and eliminate metabolic wastes, particularly nitrogen products. In addition, they need their own supply of nutrients for the maintenance of their own tissues and cells. When the kidneys are diseased or damaged and unable to carry out their functions efficiently, the animal becomes debilitated and death often occurs quickly [
48]. The present study showed that, on day-07 and day-28, after exposure to different doses to DB the PCTs swell due to hydrophiid degeneration, the basement membrane of DCTs becomes separated and karyolysis occurred in the kidney of chickens due to the toxic effects of medium and high dose DB, respectively (
Figure 3A,B). These findings were obviously more severe on day-28 compared to day-07 and suggest that after long term treatment with DB, the chicken kidney may suffer dysfunction.
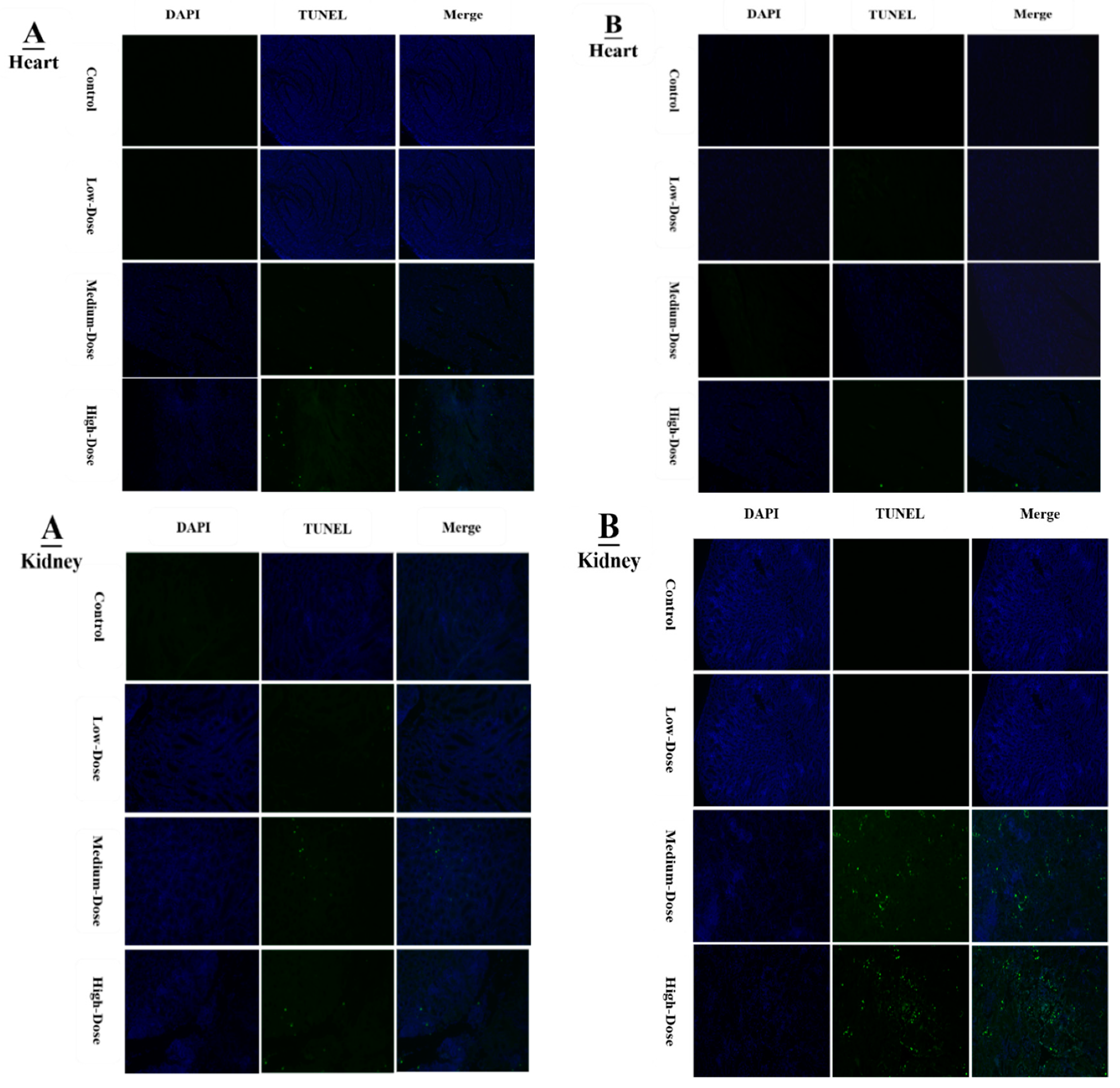
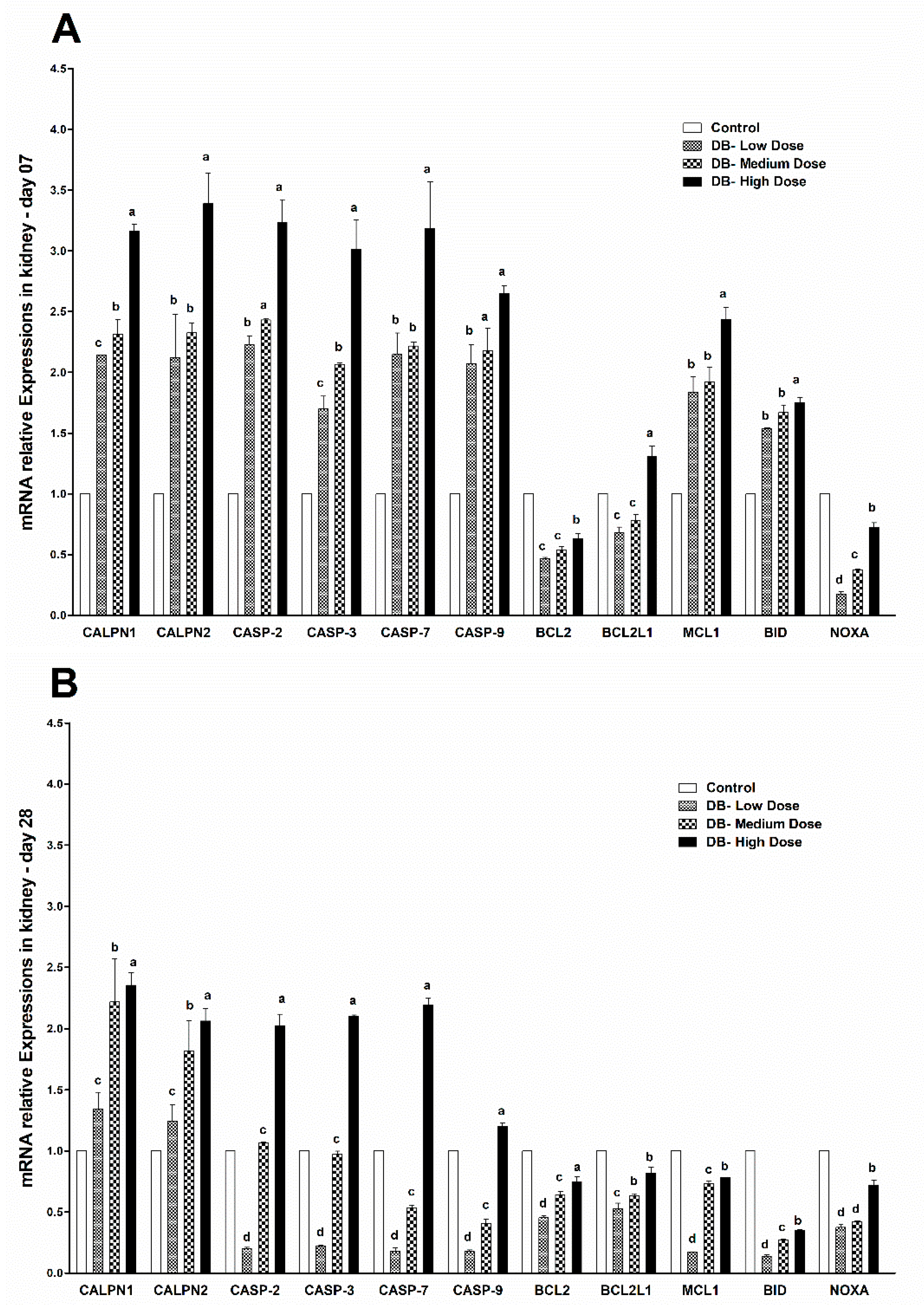
Apoptosis is involved in cellular growth and development, and is important for the turnover of heart and kidney epithelial cells and tissue homeostasis [
49]. Severe apoptosis is harmful for the heart and kidney, and can lead to cellular dysfunction [
49,
50]. It is reported that BD inhibits airway epithelial cell proliferation, decreases the number of cells and promotes cell apoptosis in a dose-dependent manner via a mitochondrial signaling pathway [
19]. We performed TUNNEL assays to confirm pathological changes caused by denatonium benzoate in the heart and kidneys of chicken. As described previously, the apoptosis-related genes showed higher expressions in the heart and kidney of chicken, pathological changes that were also confirmed by haemotoxylin and eosin staining. In the present study, we detected more serious apoptosis in the heart and kidney epithelial cells of the medium and high dose DB groups. In addition, we found greater number of apoptotic cells in the high dose DB group than in the medium-dose group at 28 days, these findings revealed that denatonium benzoate amplified apoptosis in a dose-effect manner. Consistent with our results, a previous study indicated that DB inhibited airway epithelial cell proliferation, and increased cell apoptosis in a dose-effect manner [
19]. Other studies also demonstrated that bitter-tasting compounds induced apoptosis in cancer cells [
51]. In our study, high-dose DB exerted seriously negative effects on the heart and kidney of Chinese Fast Yellow chickens. Interestingly, low-dose DB reduce the body weight without affecting the heart and kidney epithelium after long-term adaptation. We hypothesize that low-dose DB could be added into the feed for obese layers to control the body weight due to obesity-induced dysfunctions in layers [
52]. This hypothesis requires further investigations to evaluate.
The downstream signaling effectors genes (α-gustducin, PLCβ2, IP3R3 and TRPM5) of bitter taste receptors displayed similar expression patterns as the bitter taste receptors. However, these expressions were higher in day-07, while the age of chicken increased the amplified genes’ expressions (bitter and downstream signaling) in the heart and kidney of chicken were attenuated in both low-dose and medium-dose groups apparently. The results indicate that heart and kidney have a better tolerance to bitter stimuli after long-term of exposure to low and medium doses of DB. Taste transduction gene mRNA expression showed variations in the heart and kidney, through administration of DB, which suggests possible extra-gustatory effects for these genes on heart and kidney cell function of the chicken which require further investigations.
The transduction of taste is a fundamental process that allows animals to discriminate nutritious from noxious substances. Three taste modalities, bitter, sweet, and amino acid, are mediated by G protein-coupled receptors that signal through a common transduction cascade: activation of phospholipase Cβ2, leading to a breakdown of phosphatidylinositol-4,5-bisphosphate (PIP2) into diacylglycerol and inositol 1,4,5-trisphosphate, which causes release of Ca2+ from intracellular stores. The ion channel, TRPM5, is an essential component of this cascade; however, the mechanism by which it is activated is unknown. The bitter taste signaling transduction requires the involvement of Ca
2+ influx [
53,
54]. It is clarified that increased cytosolic [Ca
2+] is reversed by Ca**-ATPase [
55,
56]. Ca
2+-ATPase is responsible for actively maintaining the balance of Ca
2+ concentration within the cytoplasm and cellular organelles [
57]. In the present study, the reduced activity of Ca
2+-ATPase revealed that the function of the Ca
2+ pump was affected in the heart and kidney. In addition, in humans and rodents, mitochondrial dysfunction and oxidative damage could cause Ca
2+-ATPase damage [
58,
59]. Hence, in order to further understand the exact mechanism of Ca
2+-ATPase damage in chicken further studies are required. Likewise, in agreement with our results in this experiment, the Ca
2+-ATPase activity in low-dose DB and medium-dose DB groups were recovered with an adaptation to denatonium after 28 consecutive days of exposure. Moreover, excessive Ca
2+ concentration is able to activate the Ca
2+-dependent cysteine proteases (calpain family) [
60]. The major calpain isoforms are calpain 1 and calpain 2, which are expressed in different tissues including heart and kidney of the chicken [
61]. Calpain is demonstrated to be capable of inducing the activation of caspase family, which results in apoptosis [
56,
62]. The activation of calpain could cause tissue damage, apoptosis and autophagy [
62,
63]. Calpain 1 (u-calpain) and calpain 2 (m-calpain) require micromolar [Ca
2+] and millimolar [Ca
2+] to activate, respectively [
60]. We speculate that long-term of bitter taste receptor agonist caused [Ca**]
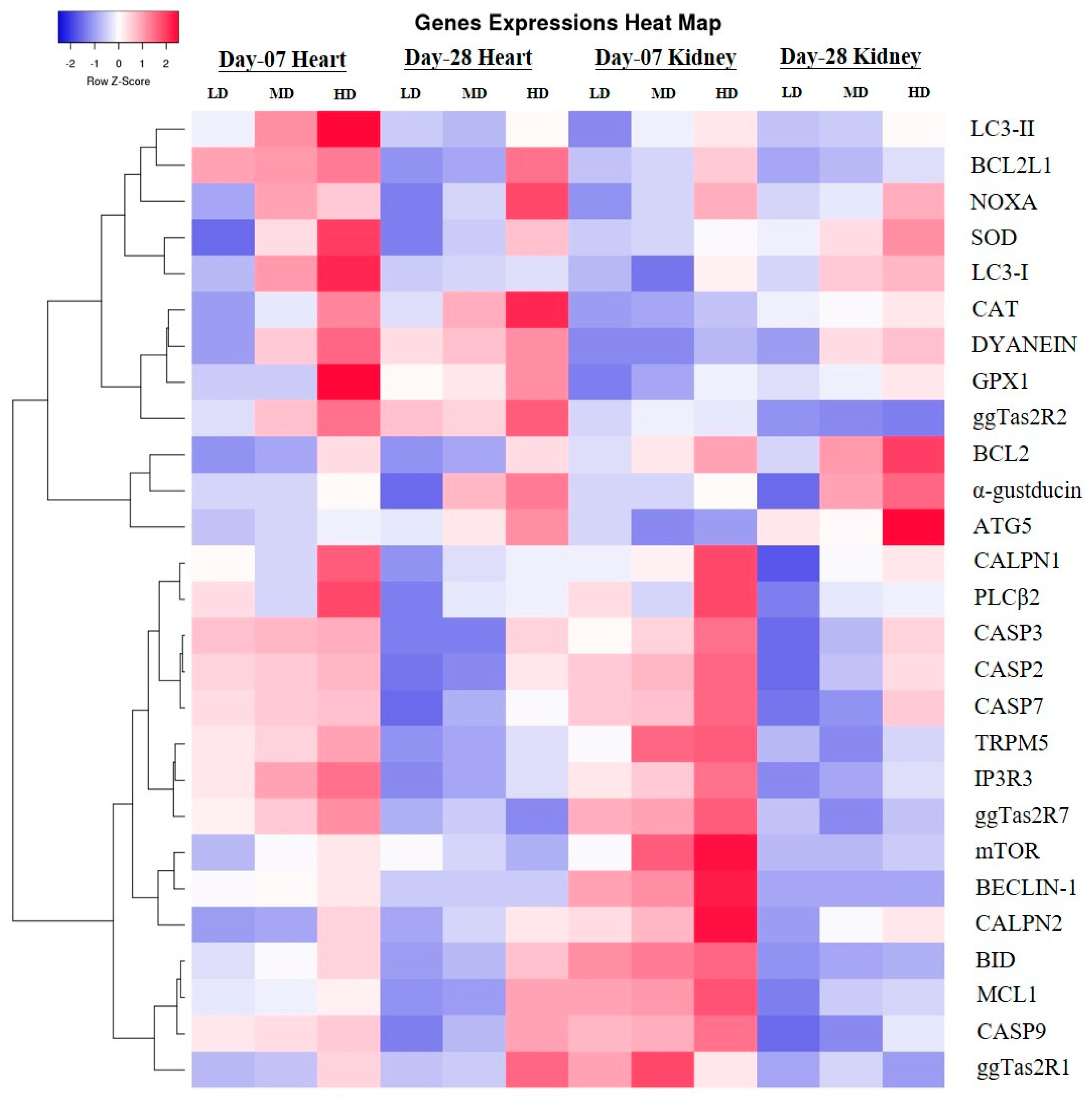
c to increase in heart and kidney epithelial cells in micromolar degree according to the result of enhanced CAPN1 expression and invariant expression of CAPN2. Elevated gene expressions of CAPN1 and apoptosis executioners (BCL2, BCL2L1, Caspase 2, Caspase 3, Caspase 7, Caspase 9, MCL1, BID, NOXA) in high-dose group indicated that high-dose DB induced more apoptosis in the heart and kidney of chicken. The apoptosis result was validated by a TUNEL assays. These data increase the possibility that after administration of DB, bitter taste receptors expressed in the heart and kidney of chicken are involved in the process of apoptosis via a calpain/caspase-dependent mechanism.
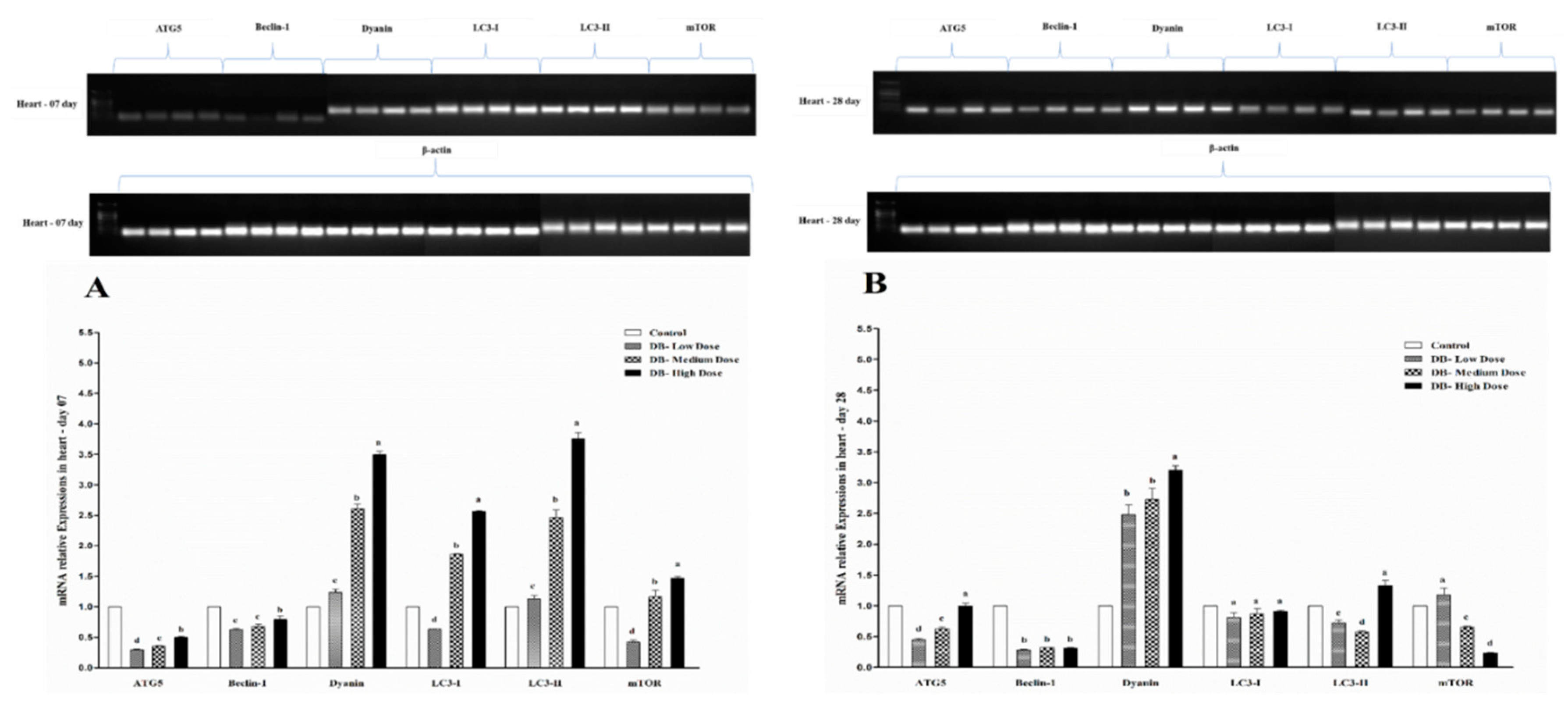
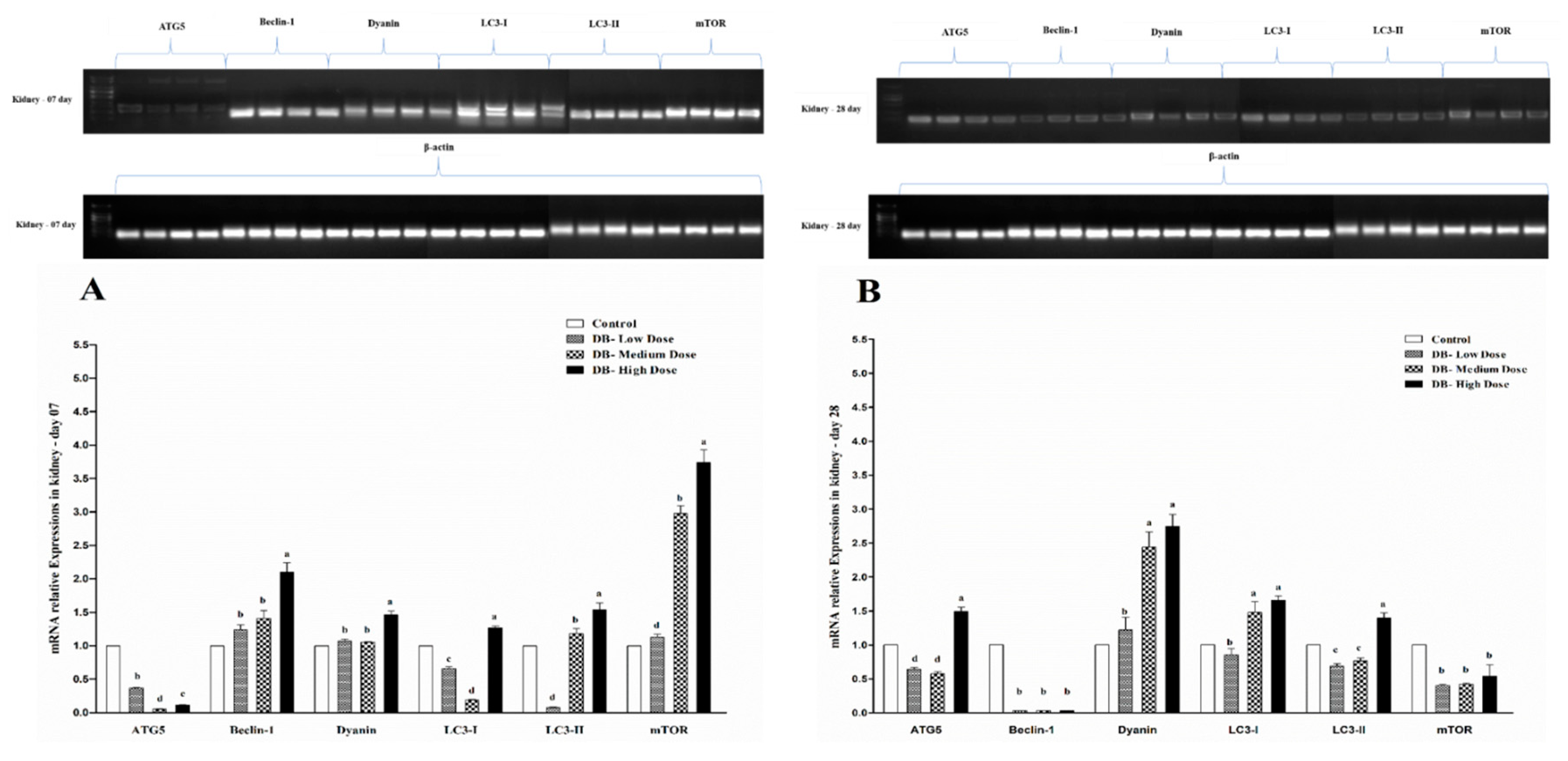
Autophagy is an evolutionary conserved catabolic process that includes different forms of digestive pathways, namely macro-autophagy, micro-autophagy, chaperone-mediated autophagy and non-canonical autophagy, regulating the degradation of a cell’s own components through the lysosomal machinery [
64]. Dramatically it plays a key homeostatic role in every cell type to maintain the balance between the synthesis, degradation, and consequent recycling of cellular components [
65]. Currently, more than thirty different autophagy-related genes have been identified by genetic screening in yeast, and many of these genes are conserved in plants, flies and mammals, respectively [
21,
66]. Particularly, Bcl-2, a major apoptosis inhibitor, binds Beclin-1 to prevent its interaction with AGT5, thus resulting in the inhibition of autophagic initiation [
67]. Conversely, when cleaved by caspase-3, Beclin-1 loses its ability to promote autophagy but renders cells sensitive to apoptosis [
68]. Some reports indicate that autophagic degradation prevents apoptosis by eliminating harmful cellular wreckages [
69,
70], whereas others suggest that boosted autophagy results in increased apoptotic vulnerability [
68,
71]. However, data in birds are rare. Here, we report our results for the first time to indicate the autophagy-related genes expressions in the heart and kidney of chicken exposed to dietary DB treatment for 28 consecutive days. All selected autophagy-related genes in this experiment (ATG5, Beclin-1, Dyanin, LC3-I, LC3-II and mTOR) had high basal expression levels in the two examined tissues from chicken both at day-07 and day-28, respectively. However, the expressions of autophagy related genes were confirmed by RT and qRT-PCR analysis. Moreover, the expressions level of ATG5, Beclin-1 and mTOR were significantly lower in day-07 and day-28, while, we visualized higher significant expressions for Dyanin, LC3-I and LC3-II in both experimental stages in the heart of chicken. Interestingly, these expressions were in contradiction with the kidney of chicken data, where we observed higher significant expressions of Beclin-1, Dyanin and mTOR at both day-07 and day-28 among other selected genes, respectively. These results suggest that autophagy may play a crucial role in regulating many toxicity- and apoptotic-related complications which may be due to exposute to BD. On the other hand, limited knowledge is available on the role of the effect of the modulation of the autophagy process in the DB exposure context in chickens.
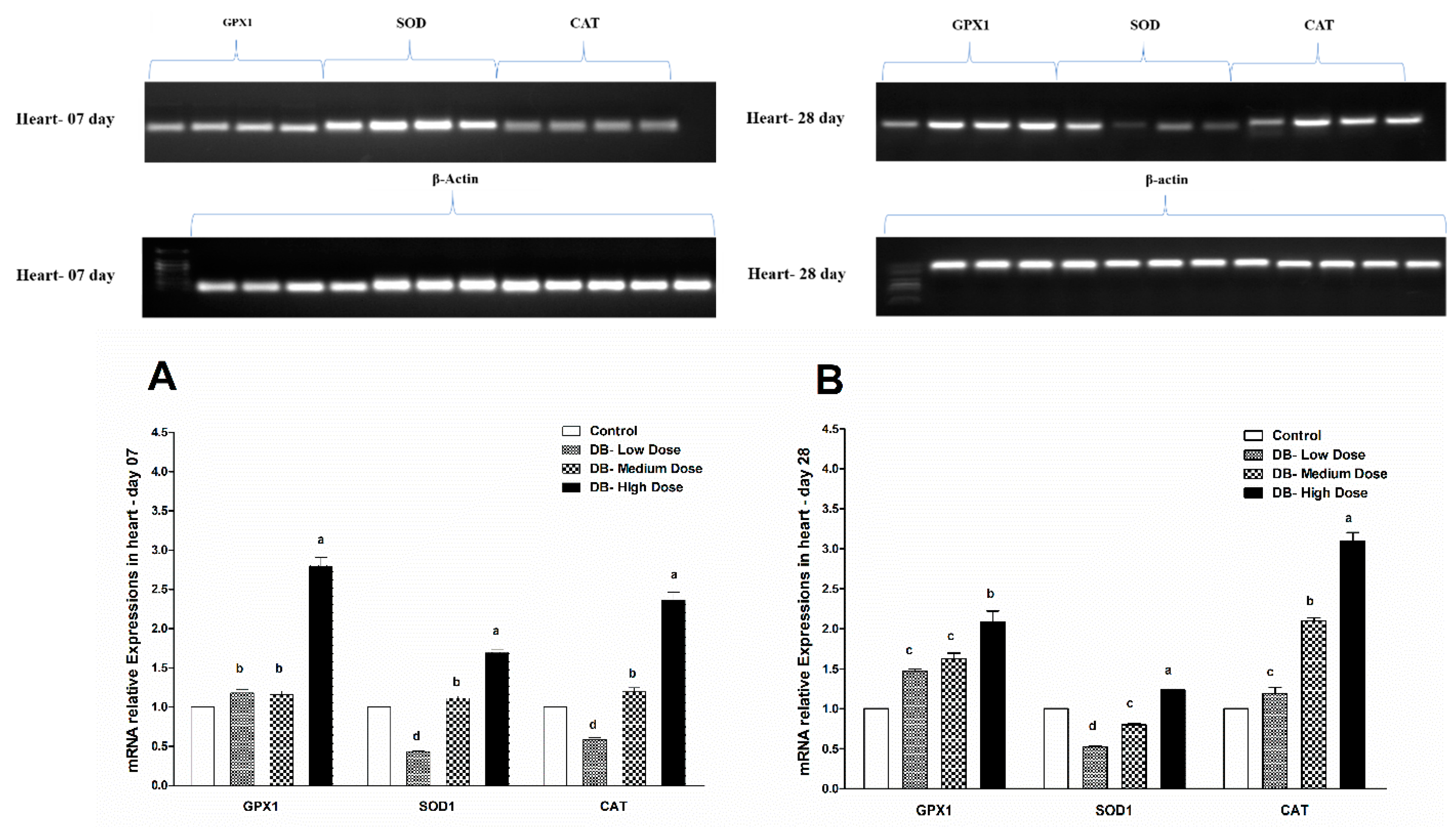
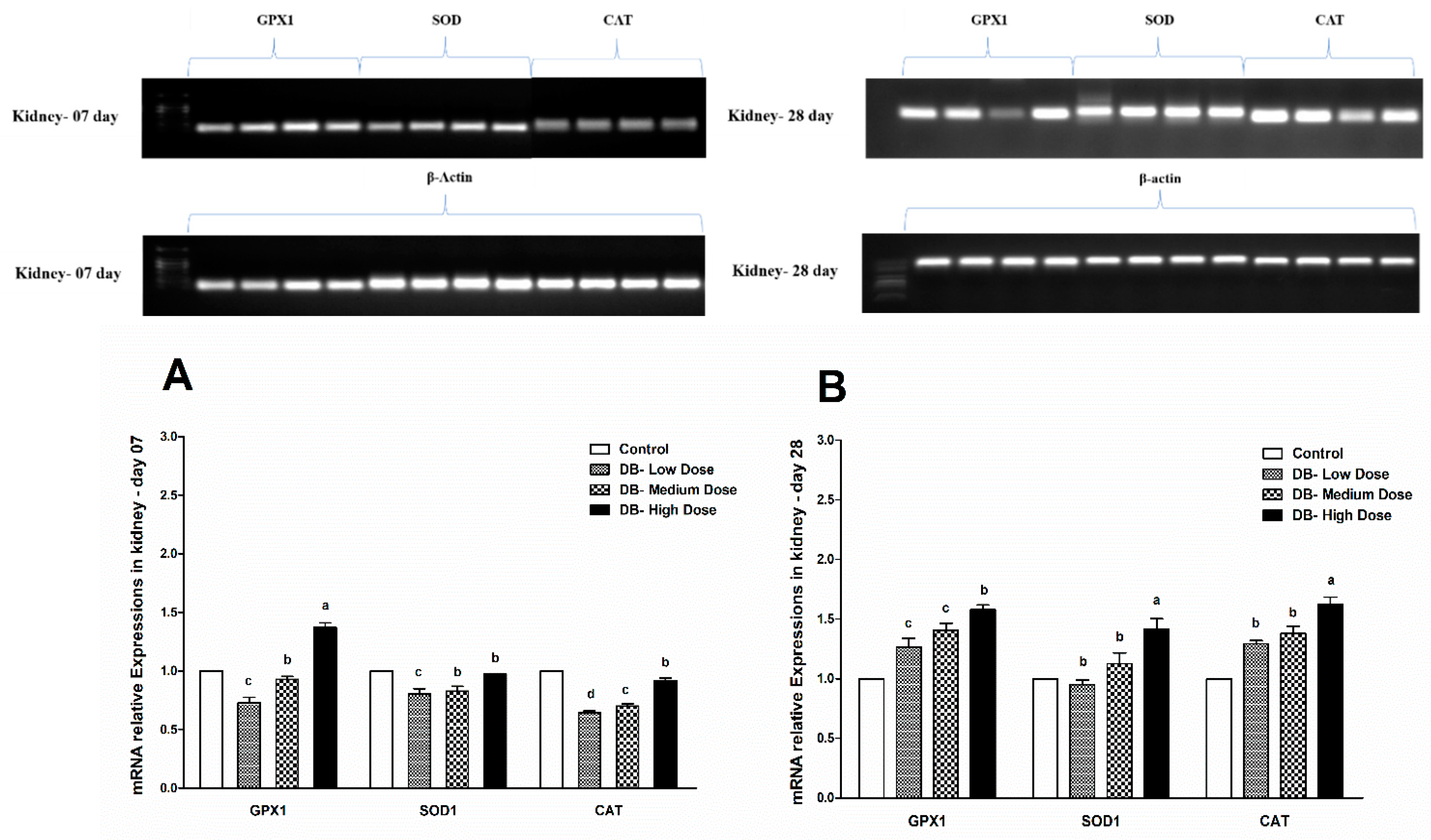
The endogenous cellular defense system consists of a number of antioxidant enzymes and proteins that maintain the cellular redox status, which is critical for various biological processes and functions. Most enzymatic components of this antioxidant defense system are commonly known as “antioxidant enzymes” (e.g., catalase, superoxide dismutase, glutathione peroxidase). Additionally, several experimental works evaluate the activity and expression of such antioxidant enzymes in different physiological conditions as a parameter to assess oxidative stress in a given system. As reported by Yuzhalin and Kutikhin [
72]. Long-term accumulation of ROS and high levels of reactive oxygen species (ROS) may enhance oxidative damage at the DNA level. This process may affect several genes responsible for the regulation of proliferation, growth, survival, apoptosis, autophagy, invasion, leading to genomic instability and deregulation of several pathways [
72]. Several enzymes, such as super oxide dismutase (SOD), glutathione peroxidase (GPX), catalase (CAT), nitric oxide synthase (NOS), and paraoxon’s (PON), function to prevent damage caused by ROS [
73].
Therefore, in the present study, our results indicated different expressions level in RT and qRT-PCR analysis for the confirmation of oxidative stress in the heart and kidney of chicken due to exposure to different doses of denatonium benzoate for 28 consecutive days. However, the expressions of GPX1, SOD and CAT were almost similar on both day-07 and day-28 in the heart of chicken but we observed higher significant (p < 0.005) expressions among them in the BD high-dose treatment groups. This indicates that the oxidative genes expressions are dose-responsive, and it confirms our previous apoptosis and autophagy results, while similar expression patterns were observed in the kidney of chicken in day-07 and day-28 of the experiment. Remarkably, GPX1, SOD and CAT expressions were significantly (p < 0.005) similar, while there were slightly higher expressions in the DB high-dose treatment groups, respectively. These results suggest that oxidative stress damage is correlated with apoptotic and autophagic changes in the heart and kidney of chicken in a dose-responsive manner.

 ,
, 
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}