Variance of Zein Protein and Starch Granule Morphology between Corn and Steam Flaked Products Determined Starch Ruminal Degradability Through Altering Starch Hydrolyzing Bacteria Attachment


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Chemical Profiles
2.2. Scanning Electron Microscope (SEM)
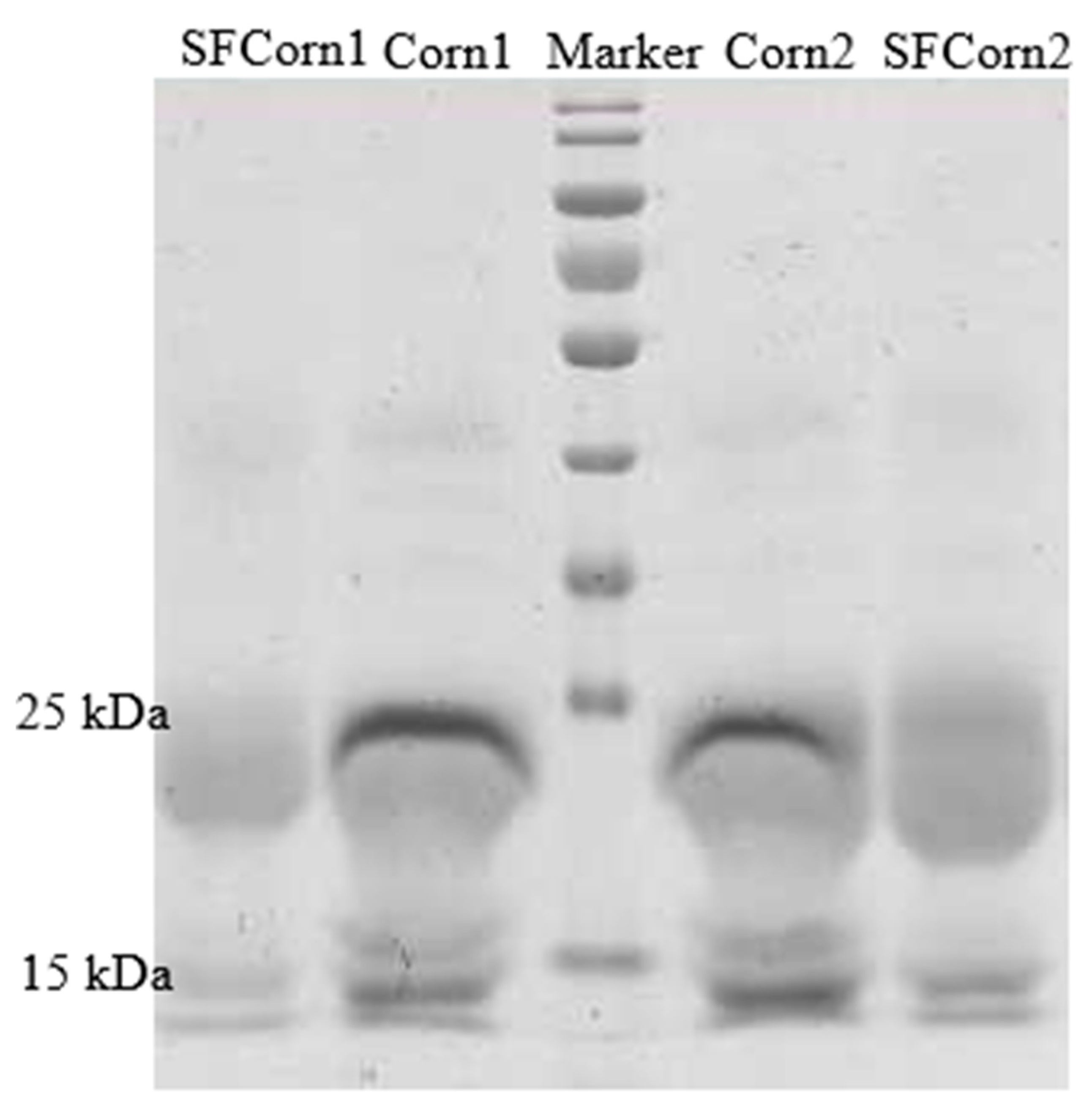
2.3. SDS-PAGE
2.4. Starch Granule Particle Size Analysis
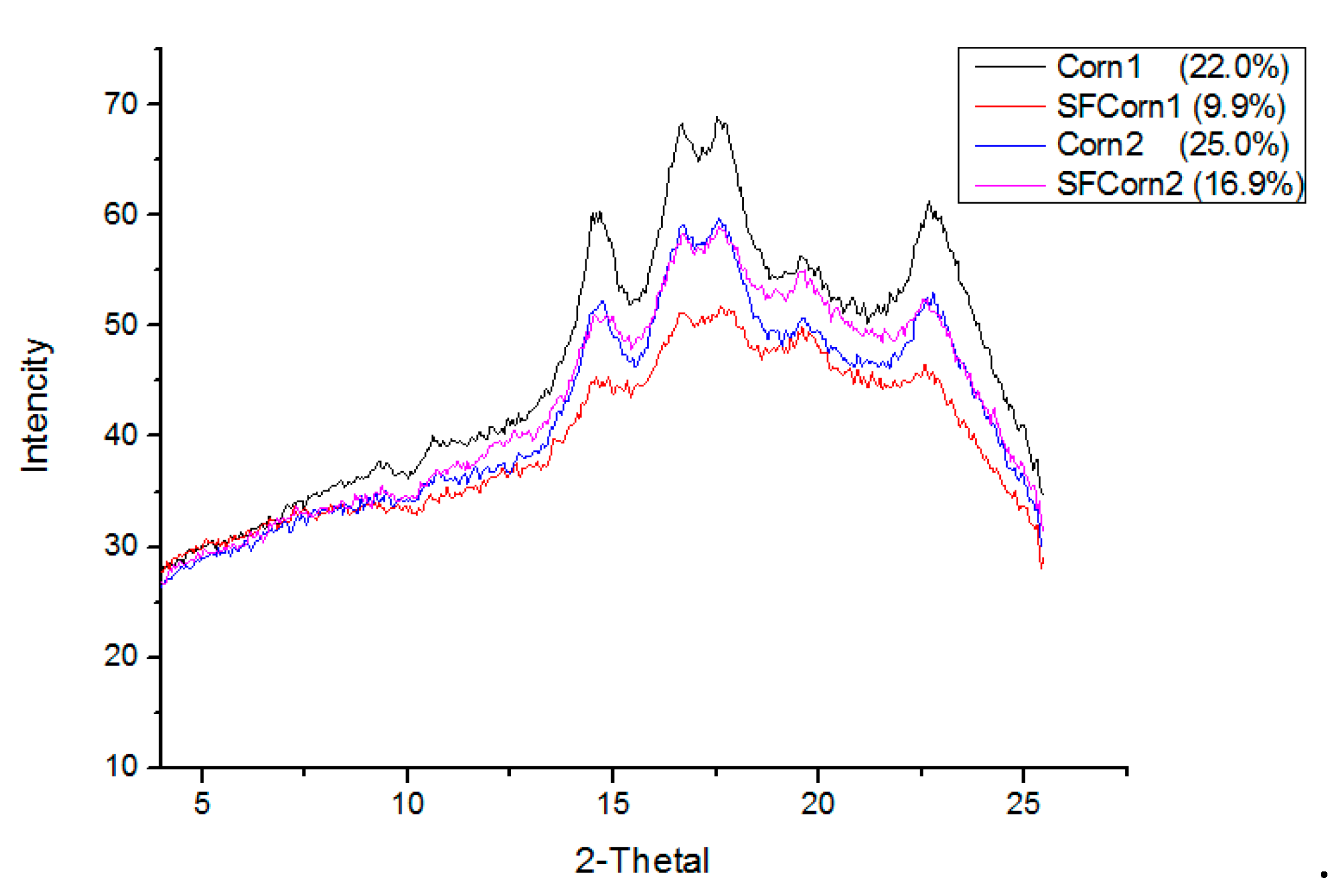
2.5. Small Angle X-Ray Scattering (SAXS)
2.6. In Situ Procedures and Starch Hydrolyzing Bacteria Collection
2.7. Quantification of Starch Hydrolyzing Bacteria
3. Results
3.1. Zein Protein Pattern and Starch–Protein Matrix
3.2. Starch Granule Morphology
3.3. Starch Granule Crystallinity
3.4. Starch Hydrolyzing Bacteria Attached to Incubation Residues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giuberti, G.; Gallo, A.; Masoero, F. Quantification of zeins from corn, high-moisture corn, and corn silage using a turbidimetric method: Comparative efficiencies of isopropyl and tert-butyl alcohols. J. Dairy Sci. 2012, 95, 3384–3389. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.; Esser, N.M.; Shaver, R.D.; Coblentz, W.K.; Scott, M.P.; Bodnar, A.L.; Schmidt, R.J.; Charley, R.C. Influence of ensiling time and inoculation on alteration of the starch-protein matrix in high-moisture corn. J. Dairy Sci. 2011, 94, 2465–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotarski, S.F.; Waniska, R.D.; Thurn, K.K. Starch hydrolyzing by the ruminal microflora. J. Nutr. 1992, 122, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Hamaker, B.; Mohamed, A.; Habben, J.; Huang, C.; Larkins, B. Efficient procedure for extracting maize and sorghum kernel proteins reveals higher prolamin contents than the conventional method. Cereal Chem. 1995, 72, 583–588. [Google Scholar]
- Tester, R.; Karkalas, J.; Qi, X. Starch structure and digestibility enzyme-substrate relationship. Worlds Poult. Sci. J. 2004, 60, 186–195. [Google Scholar] [CrossRef]
- Naguleswaran, S.; Li, J.; Vasanthan, T.; Bressler, D.; Hoover, R. Amylolysis of large and small granules of native triticale, wheat and corn starches using a mixture of α-amylase and glucoamylase. Carbohydr. Polym. 2012, 88, 864–874. [Google Scholar] [CrossRef]
- Ferraretto, L.F.; Crump, P.M.; Shaver, R.D. Effect of cereal grain type and corn grain harvesting and processing methods on intake, digestion, and milk production by dairy cows through a meta-analysis. J. Dairy Sci. 2013, 96, 533–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.N.; Liu, J.X.; Yu, P.Q. Alteration of biomacromolecule in corn by steam flaking in relation to biodegradation kinetics in ruminant, revealed with vibrational molecular spectroscopy. Spectrochim. Acta, A. Mol. Biomol. Spectrosc. 2018, 191, 491–497. [Google Scholar] [CrossRef] [PubMed]
- McDonough, C.; Anderson, B.; Rooney, L. Structural characteristics of steam-flaked sorghum. Cereal Chem. 1997, 74, 542–547. [Google Scholar] [CrossRef]
- Kokić, B.; Lević, J.; Chrenková, M.; Formelová, Z.; Poláčikov, M.; Rajský, M.; Jovanović, R. Influence of thermal treatments on starch gelatinization and in vitro organic matter digestibility of corn. Food & Feed Res. 2013, 40, 93–99. [Google Scholar]
- Cotta, M.A. Amylolytic activity of selected species of ruminal bacteria. Appl. Environ. Microbiol. 1988, 54, 772–776. [Google Scholar] [PubMed]
- Xia, Y.; Kong, Y.; Seviour, R.; Yang, H.E.; Forster, R.; Vasanthan, T.; McAllister, T.A. In situ identification and quantification of starch-hydrolyzing bacteria attached to barley and corn grain in the rumen of cows fed barley-based diets. FEMS Microbiol. Ecol. 2015, 91, 8. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.S.; Singh, N.; Malhi, N.S. Physicochemical and thermal properties of starches separated from corn produced from crosses of two germ pools. Food Chem. 2005, 89, 541–548. [Google Scholar] [CrossRef]
- Liu, H.; Shi, J.P.; Sun, C.L.; Gong, H.; Fan, X.M.; Qiu, F.Z.; Huang, X.H.; Feng, Q.; Zheng, X.; Yuan, N.N.; et al. Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize. Proc. Natl Acad. Sci. 2016, 113, 4964–4969. [Google Scholar] [CrossRef] [PubMed]
- Dhital, S.; Shrestha, A.K.; Gidley, M.J. Relationship between granule size and in vitro digestibility of maize and potato starches. Carbohyd. Polym. 2010, 8, 480–488. [Google Scholar] [CrossRef]
- Shrestha, A.K.; Blazek, J.; Flanagana, B.M.; Dhitala, S.; Larroquec, O.; Morellc, M.K.; Gilbertb, E.P.; Gidleya, M.J. Molecular, mesoscopic and microscopic structure evolution during amylase digestion of extruded maize and high amylose maize starches. Carbohyd. Polym. 2015, 118, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Blazek, J.; Gilbert, E.P. Effect of enzymatic hydrolyzing on native starch granule structure. Biomacromolecules 2010, 11, 3275–3289. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Kong, Y.; Nielsen, P.H. In situ detection of starch-hydrolyzing microorganisms in activated sludge. FEMS Microbiol. Ecol. 2008, 66, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image processing with image. J. Biophotonics Int. 2004, 11, 36–41. [Google Scholar]
- French, D. Starch: Chemical Technology, 2nd ed.; Academic Press: New York, NY, USA, 1984; pp. 183–247. [Google Scholar]
- Owens, F.; Zinn, R.; Kim, Y. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 1986, 63, 1634–1648. [Google Scholar] [CrossRef]
- McAllister, T.; Phillippe, R.; Rode, L.; Cheng, K. Effect of the protein matrix on the digestion of cereal grains by ruminal microorganisms. J. Anim. Sci. 1993, 71, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Ngonyamo-Majee, D.; Shaver, R.; Coors, J.; Sapienza, D.; Lauer, J. Relationships between kernel vitreousness and dry matter degradability for diverse corn germplasm: II. Ruminal and post-ruminal degradabilities. Anim. Feed Sci. Tech. 2008, 142, 259–274. [Google Scholar] [CrossRef]
- Philippeau, C.; Landry, J.; Michalet-Doreau, B. Influence of the protein distribution of maize endosperm on ruminal starch degradability. J. Sci. Food Agr. 2000, 80, 404–408. [Google Scholar] [CrossRef]
- Hasjim, J.; Srichuwong, S.; Scott, M.P.; Jane, J.L. Kernel composition, starch structure, and enzyme digestibility of opaque-2 maize and quality protein maize. J. Agri. Food Chem. 2009, 57, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.C.; Shaver, R.D.; Hoffman, P.C.; Akins, M.S.; Bertics, S.J.; Gencoglu, H.; Coors, J.G. Type of corn endosperm influences nutrient digestibility in lactating dairy cows. J. Dairy Sci. 2009, 92, 4541–4548. [Google Scholar] [CrossRef]
- Sadeghi, A.A.; Shawrang, P. Effects of microwave irradiation on ruminal protein and starch degradation of corn grain. Anim. Feed Sci. Tech. 2006, 127, 113–123. [Google Scholar] [CrossRef]
- Carré, B. Causes for variation in digestibility of starch among feedstuffs. World Poultry Sci. J. 2004, 60, 76–89. [Google Scholar] [CrossRef]
- Bednar, G.E.; Patil, A.R.; Murray, S.M.; Grieshop, C.M.; Merchen, N.; Fahey, G.C. Starch and fiber fractions in selected food and feed ingredients affect their small intestinal digestibility and fermentability and their large bowel fermentability in vitro in a canine model. J. Nutr. 2001, 131, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Benmoussa, M.; Suhendra, B.; Aboubacar, A.; Hamaker, B.R. Distinctive sorghum starch granule morphologies appear to improve raw starch digestibility. Starch-Stärke 2006, 58, 92–99. [Google Scholar] [CrossRef]
- Cai, H.; Dave, V.; Gross, R.A.; McCarthy, S.P. Effects of physical aging, crystallinity, and orientation on the enzymatic degradation of poly (lactic acid). J. Polym. Sci. Pol. Phys. 1996, 34, 2701–2708. [Google Scholar] [CrossRef]
- McAllister, T.A.; Bae, H.D.; Jones, G.A.; Cheng, K.J. Microbial attachment and feed digestion in the rumen. J. Anim. Sci. 1994, 72, 3004–3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, T.A.; Cheng, K.J. Microbial strategies in the ruminal digestion of cereal grains. Anim. Feed Sci. Tech. 1996, 62, 29–36. [Google Scholar] [CrossRef]
- Leitch, E.C.M.W.; Walker, A.W.; Duncan, S.H.; Holtrop, G.; Flint, H.J. Selective colonization of insoluble substrates by human faecal bacteria. Environ. Microbiol. 2007, 9, 667–679. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Mastersizer Data | ||||
|---|---|---|---|---|---|
| D10 | D25 | D50 | D75 | D90 | |
| Corn1 | 11.15 | 14.29 | 17.77 | 21.70 | 26.97 |
| SFCorn1 | 11.29 | 20.02 | 30.83 | 50.27 | 96.34 |
| Corn2 | 11.24 | 14.45 | 17.98 | 24.44 | 27.15 |
| SFCorn2 | 9.99 | 17.86 | 26.03 | 45.20 | 54.60 |
| Sample | Incubation Time (h) | SEM | P | |||
|---|---|---|---|---|---|---|
| 4 | 12 | 24 | 48 | |||
| Corn 1, % | 43.9aA | 28.8b | 20.1bB | 31.5b | 3.73 | 0.01 |
| SFCorn 1, % | 34.6bB | 23.3b | 44.9aA | 35.7a | 4.06 | 0.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, N.; Wang, D.; Liu, J. Variance of Zein Protein and Starch Granule Morphology between Corn and Steam Flaked Products Determined Starch Ruminal Degradability Through Altering Starch Hydrolyzing Bacteria Attachment. Animals 2019, 9, 626. https://doi.org/10.3390/ani9090626
Xu N, Wang D, Liu J. Variance of Zein Protein and Starch Granule Morphology between Corn and Steam Flaked Products Determined Starch Ruminal Degradability Through Altering Starch Hydrolyzing Bacteria Attachment. Animals. 2019; 9(9):626. https://doi.org/10.3390/ani9090626
Chicago/Turabian StyleXu, NingNing, DiMing Wang, and JianXin Liu. 2019. "Variance of Zein Protein and Starch Granule Morphology between Corn and Steam Flaked Products Determined Starch Ruminal Degradability Through Altering Starch Hydrolyzing Bacteria Attachment" Animals 9, no. 9: 626. https://doi.org/10.3390/ani9090626