Improving Terrestrial Squamate Surveys with Camera-Trap Programming and Hardware Modifications


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Time-Lapse and Passive Infrared Triggers
1.2. A More Sensitive PIR Sensor
1.3. In Focus
2. Materials and Methods
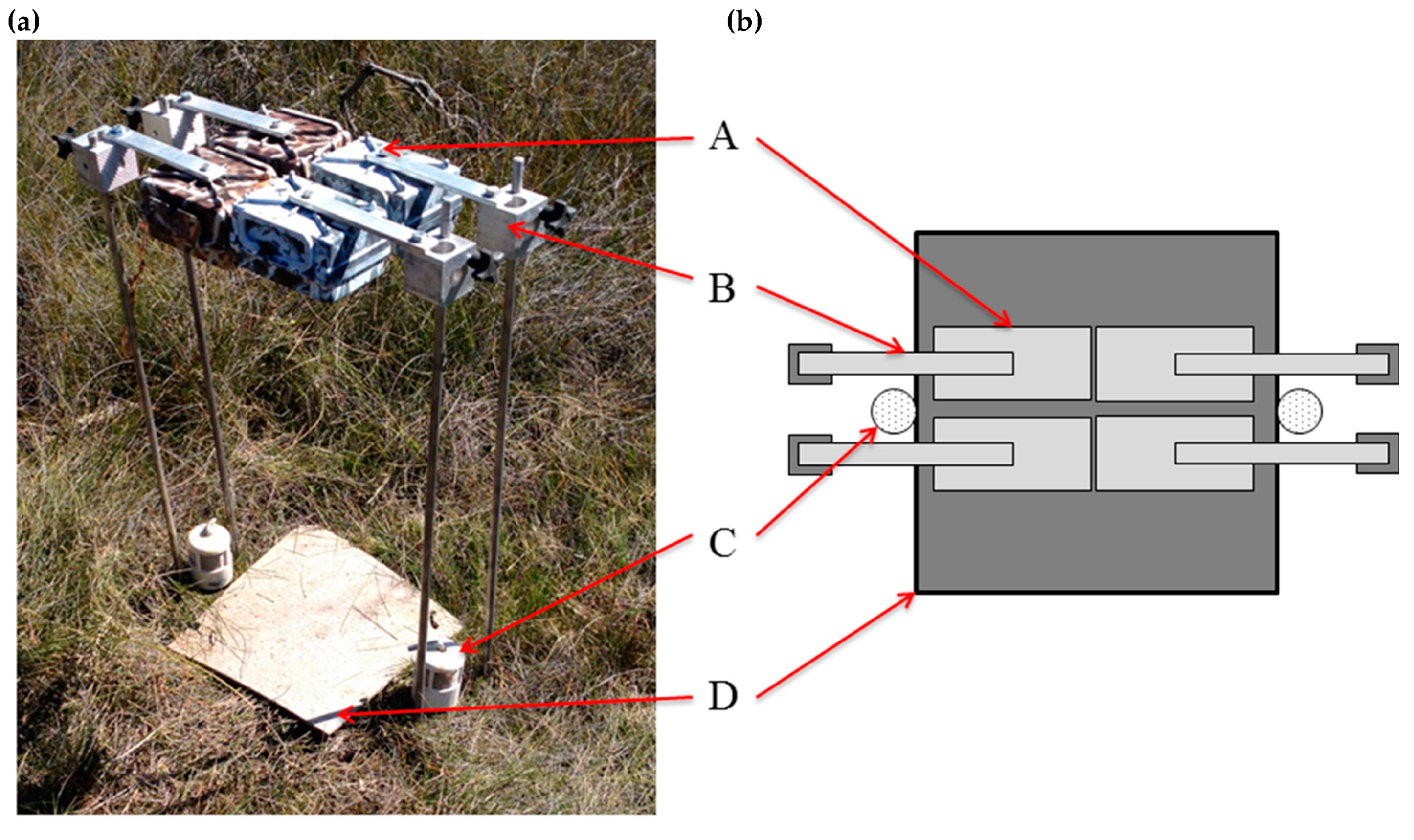
2.1. Study Design
2.2. Camera-Trap Programming
2.3. Camera-Trap Modification
3. Results
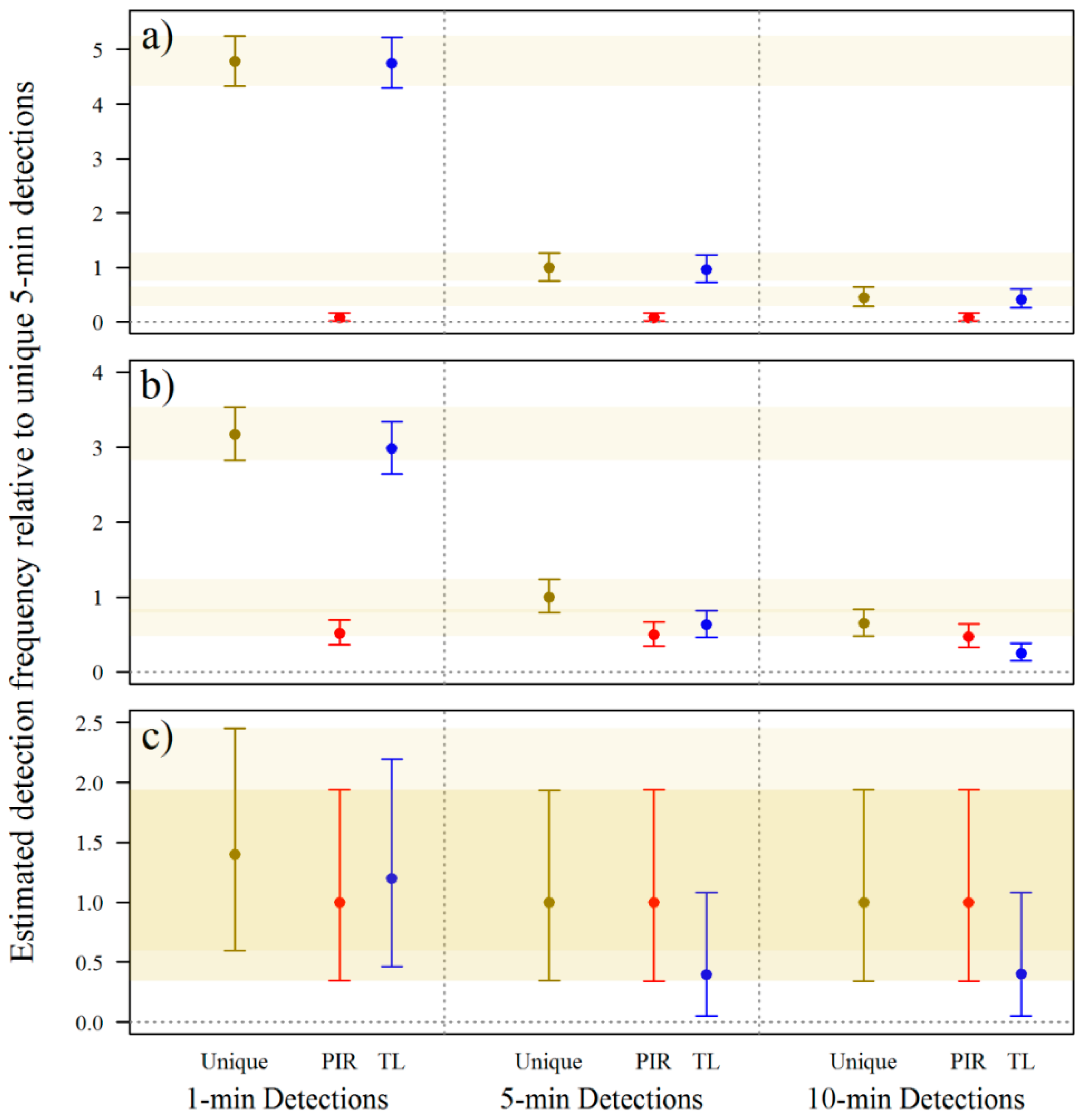
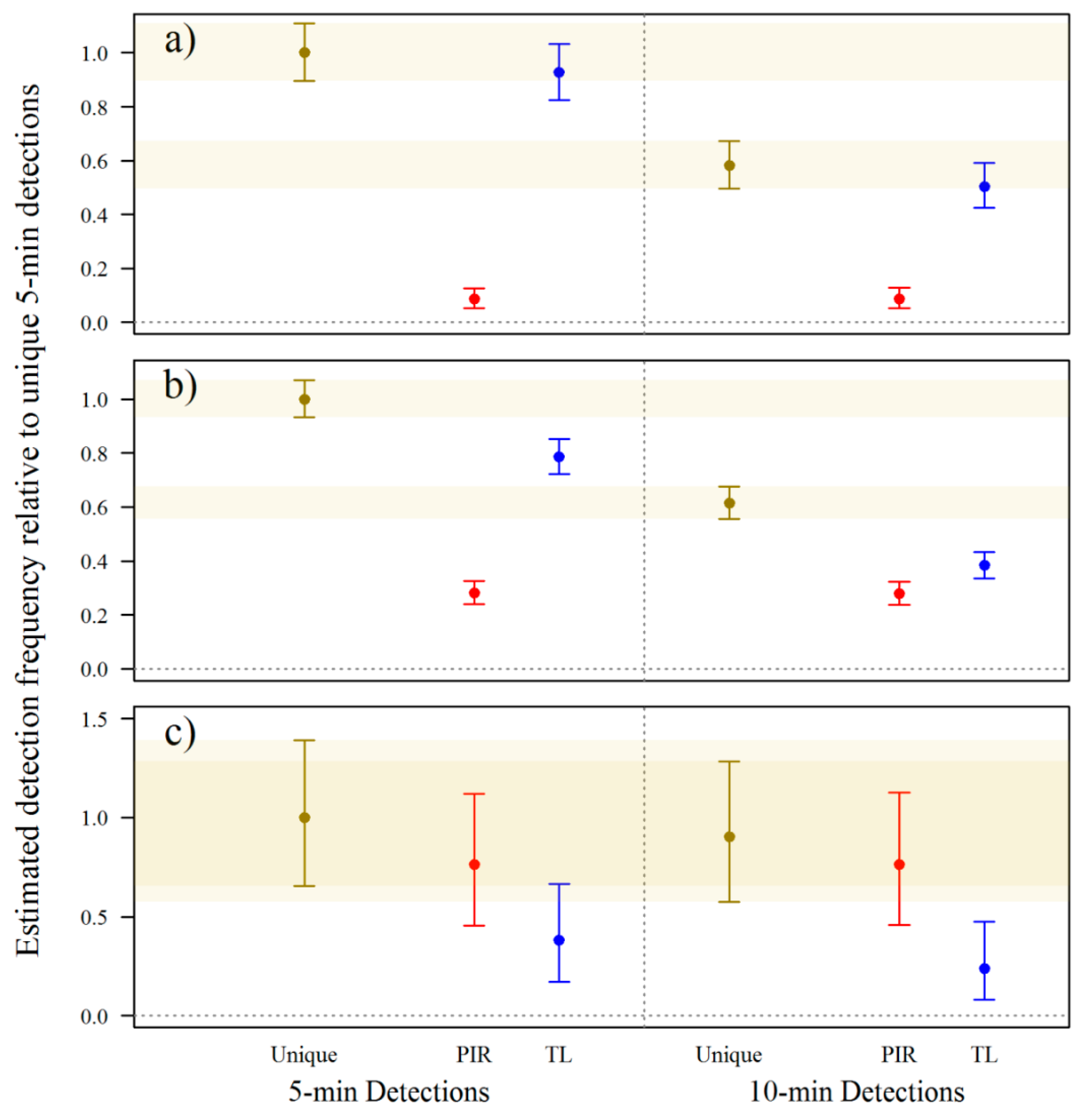
3.1. Camera-Trap Programming
3.2. Camera-Trap Modification
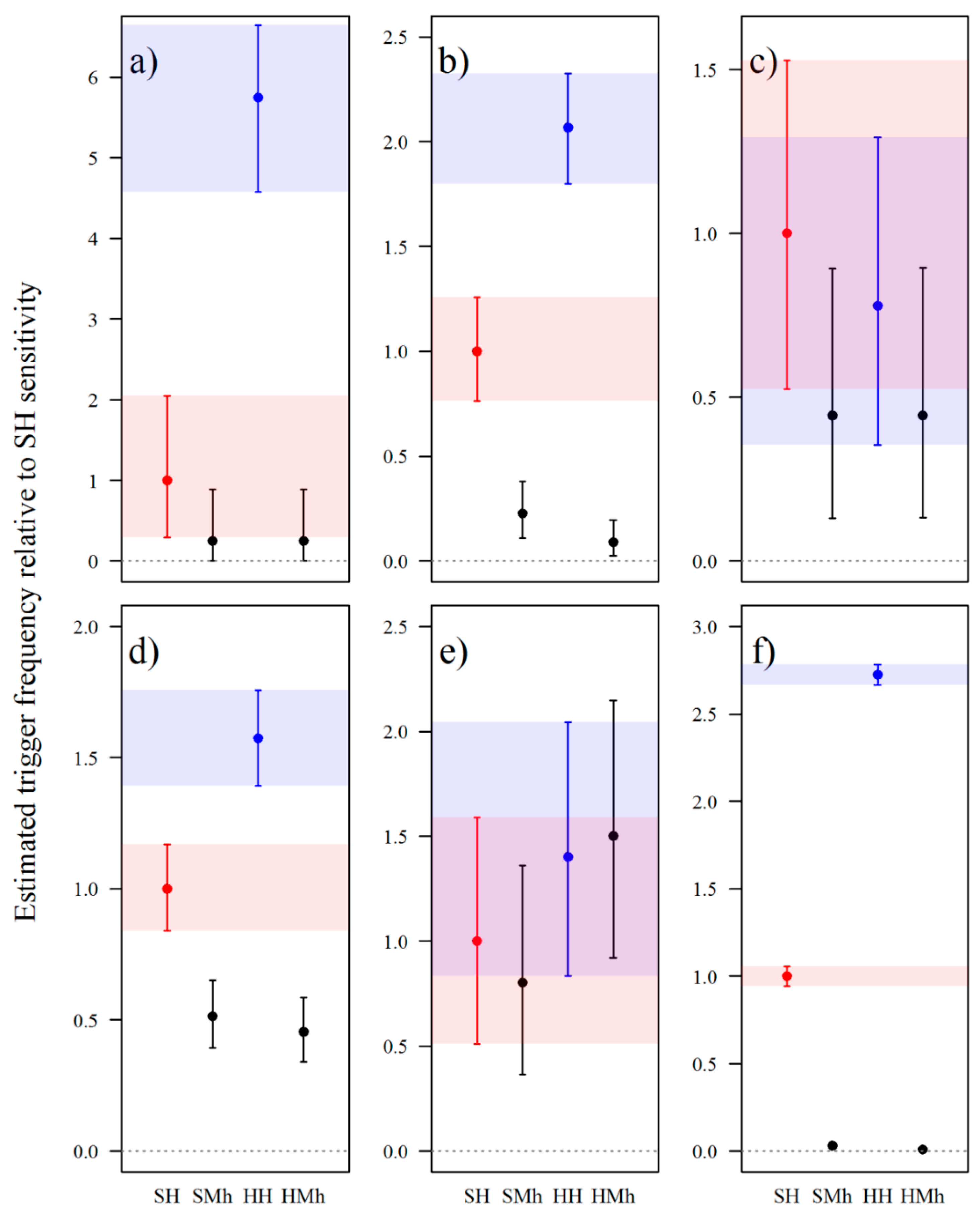
3.2.1. Passive Infrared Sensitivity
3.2.2. Focal Length
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meek, P.; Fleming, P.; Ballard, G.; Banks, P.; Claridge, A.; Sanderson, J.; Swann, D. Camera Trapping Wildlife Management and Research; CSIRO Publishing: Melbourne, Australia, 2014; p. 367. [Google Scholar]
- Torretta, E.; Mosini, A.; Piana, M.; Tirozzi, P.; Serafini, M.; Puopolo, F.; Saino, N.; Balestrieri, A. Time partitioning in mesocarnivore communities from different habitats of NW Italy: Insights into martens’ competitive abilities. Behaviour 2017, 154, 241–266. [Google Scholar] [CrossRef]
- Welbourne, D.J.; Paull, D.J.; Claridge, A.W.; Ford, F. A frontier in the use of camera traps: Surveying terrestrial squamate assemblages. Remote Sens. Ecol. Conserv. 2017, 3, 133–145. [Google Scholar] [CrossRef]
- Welbourne, D. A method for surveying diurnal terrestrial reptiles with passive infrared automatically triggered cameras. Herpetol. Rev. 2013, 44, 247–250. [Google Scholar]
- Adams, C.S.; Ryberg, W.A.; Hibbitts, T.J.; Pierce, B.L.; Pierce, J.B.; Rudolph, D.C. Evaluating effectiveness and cost of time-lapse triggered camera trapping techniques to detect terrestrial squamate diversity. Herpetol. Rev. 2017, 48, 44–48. [Google Scholar]
- Dundas, S.J.; Ruthrof, K.X.; Hardy, G.E.S.; Fleming, P.A. Pits or pictures: a comparative study of camera traps and pitfall trapping to survey small mammals and reptiles. Wildlife Res. 2019, 46, 104–113. [Google Scholar] [CrossRef]
- Richardson, E.; Nimmo, D.G.; Avitable, S.; Tworkowski, L.; Watson, S.J.; Welbourne, D.; Leonard, S.W.J. Camera traps and pitfalls: An evaluation of two methods for surveying reptiles in a semiarid ecosystem. Wildlife Res. 2018, 44, 637–647. [Google Scholar] [CrossRef]
- Welbourne, D.J.; MacGregor, C.; Paull, D.; Lindenmayer, D.B. The effectiveness and cost of camera traps for surveying small reptiles and critical weight range mammals: a comparison with labour-intensive complementary methods. Wildlife Res. 2015, 42, 414–425. [Google Scholar] [CrossRef]
- Fenner, A.L.; Pavey, C.R.; Bull, C.M. Behavioural observations and use of burrow systems by an endangered Australian arid-zone lizard, Slater’s skink (Liopholis slateri). Aust. J. Zool. 2012, 60, 127–132. [Google Scholar] [CrossRef]
- Gibson, S.; Penniket, S.; Cree, A. Are viviparous lizards from cool climates ever exclusively nocturnal? Evidence for extensive basking in a New Zealand gecko. Biol. J. Linn. Soc. 2015, 115, 1–14. [Google Scholar] [CrossRef]
- Johnston, L. Trials and Evaluations of Monitoring Tools for New Zealand’s Alpine Skinks: The Barrier skink (Oligosoma judgei) and the Sinbad skink (O. pikitanga). Post-graduate Diploma in Wildlife Management, University of Otago, Dunedin, New Zealand, 2014. [Google Scholar]
- Milne, T.; Bull, C.M.; Hutchinson, M.N. Use of burrows by the endangered pygmy blue-tongue lizard, Tiliqua adelaidensis (Scincidae). Wildlife Res. 2003, 30, 523–528. [Google Scholar] [CrossRef]
- Pagnucco, K.; Paszkowski, C.A.; Scrimgeour, G.J. Using cameras to monitor tunnel use by long-toed salamanders (Ambystoma macrodactylum): An informative, cost-efficient technique. Herpetol. Conserv. Biol. 2011, 6, 277–286. [Google Scholar]
- Welbourne, D.J.; Claridge, A.W.; Paull, D.J.; Lambert, A. How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions. Remote Sens. Ecol. Conserv. 2016, 2, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Ballard, G.; Meek, P.D.; Doak, S.; Fleming, P.J.S.; Sparkes, J. Camera traps, sand plots and known events: What do camera traps miss? In Camera Trapping Wildlife Management and Research; Meek, P., Fleming, P., Ballard, G., Banks, P., Claridge, A., Sanderson, J., Swann, D., Eds.; CSIRO Publishing: Melbourne, Australia, 2014; pp. 189–202. [Google Scholar]
- Wellington, K.; Bottom, C.; Merrill, C.; Litvaitis, J.A. Identifying performance differences among trail cameras used to monitor forest mammals. Wildl. Soc. Bull. 2014, 38, 634–638. [Google Scholar] [CrossRef]
- Thinner, J.; Reconyx, USA. Personal communication, 2012.
- Damm, P.E.; Grand, J.B.; Barnett, S.W. Variation in detection among passive infrared triggered-cameras used in wildlife research. In Proceedings of the Annual Conference Southeast Association Fish and Wildlife Agencies, Biloxi, MI, USA, 17–20 October 2010; pp. 125–130. [Google Scholar]
- Kelly, M.J.; Holub, E.L. Camera trapping of carnivores: Trap success among camera types and across species, and habitat selection by species, on Salt Pond Mountain, Giles County, Virginia. Northeast Nat. 2008, 15, 249–262. [Google Scholar] [CrossRef]
- Swan, M.; Di Stefano, J.; Christie, F. Comparing the effectiveness of two types of camera trap for surveying ground-dwelling mammals. In Camera Trapping Wildlife Management and Research; Meek, P., Fleming, P., Ballard, G., Banks, P., Claridge, A., Sanderson, J., Swann, D., Eds.; CSIRO Publishing: Melbourne, Australia, 2014; pp. 123–130. [Google Scholar]
- Swann, D.E.; Hass, C.C.; Dalton, D.C.; Wolf, S.A. Infrared-triggered cameras for detecting wildlife: An evaluation and review. Wildl. Soc. Bull. 2004, 32, 357–365. [Google Scholar] [CrossRef]
- Urlus, J.; McCutcheon, C.; Gilmore, D.; McMahon, J. The effect of camera trap type on the probability of detecting different size classes of Australian mammals. In Camera Trapping Wildlife Management and Research; Meek, P., Fleming, P., Ballard, G., Banks, P., Claridge, A., Sanderson, J., Swann, D., Eds.; CSIRO Publishing: Melbourne, Australia, 2014; pp. 111–121. [Google Scholar]
- Meek, P.D.; Ballard, G.; Fleming, P. An Introduction to Camera Trapping for Wildlife Surveys in Australia. Unit, V.P.R., Ed. Department of Primary Industries: Orange, Australia, 2012; p 85.
- Reconyx. Hyperfire High Performance Cameras Instruction Manual; Reconyx: Holmen, Wisconsin, USA, 2010; p. 31. [Google Scholar]
- Wilson, S.; Swan, G. Complete Guide to Reptiles of Australia, 3rd ed.; New Holland Publishers: Sydney, Australia, 2010; p. 558. [Google Scholar]
- McCarthy, M.A. Bayesian Methods for Ecology; Cambridge University Press: New York, NY, USA, 2012; p. 296. [Google Scholar]
- Conti, C. lens_test_chart. Available online: http://chriscontiphotography.com/stuff/lens_test_chart.jpg (accessed on 11 March 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; 3.4.2; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Thomas, N. R2OpenBUGS, R Core Team: R CRAN, 2015.
- Spiegelhalter, D.; Thomas, A.; Best, N.; Lunn, D. OpenBUGS User Manual; 3.2.3; MRC Biostatistics Unit: Cambridge, UK, 2014. [Google Scholar]
- Plummer, M. coda, R Core Team: R CRAN, 2015.
- Yau, C. R Tutorial with Bayesian Statistics Using OpenBUGS; Amazon Kindle: Seattle, WA, USA, 2015; p. 554. [Google Scholar]
- Uetz, P. Species Numbers (as of March 2018). Available online: http://www.reptile-database.org/db-info/SpeciesStat.html (accessed on 20 April 2019).
- IUCN. The IUCN Red List of Threatened Species. Available online: http://cmsdocs.s3.amazonaws.com/summarystats/2017-3_Summary_Stats_Page_Documents/2017_3_RL_Stats_Table_1.pdf (accessed on 10 April 2019).
- Trailcampro. Cellular Trail Cameras. Available online: http://www.trailcampro.com/collections/cellular-trail-cameras (accessed on 31 May 2019).
- Soanes, K.; Vesk, P.A.; van der Ree, R. Monitoring the use of road-crossing structures by arboreal marsupials: Insights gained from motion-triggered cameras and passive integrated transponder (PIT) tags. Wildlife Res. 2015, 42, 241–256. [Google Scholar] [CrossRef]
- Villa, A.G.; Salazar, A.; Vargas, F. Towards automatic wild animal monitoring: Identification of animal species in camera-trap images using very deep convolutional neural networks. Ecol. Inform. 2017, 41, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Mlsna, T.; Reconyx, USA. Personal communication, 2016.
- Harvey, E.; Fletcher, D.; Shortis, M. Estimation of reef fish length by divers and by stereo-video—A first comparison of the accuracy and precision in the field on living fish under operational conditions. Fish Res. 2002, 57, 255–265. [Google Scholar] [CrossRef]
- Trailcampro. Bushnell Natureview Cam HD Live View. Available online: http://www.trailcampro.com/products/bushnell-natureview-cam-hd-liveview (accessed on 31 May 2019).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trigger Mechanism | Detection Description |
|---|---|
| Experiment 1 (Dec 2013–Feb 2014) | |
| Uni 1 | Unique detections with 1-min TL intervals. |
| Uni 5 | Unique detections with 5-min TL intervals. |
| Uni 10 | Unique detections with 10-min TL intervals. |
| TL 1 | TL detections only with 1-min TL intervals. |
| TL 5 | TL detections only with 5-min TL intervals. |
| TL 10 | TL detections only with 10-min TL intervals. |
| PIR | PIR detections only matched to interval |
| Experiment 2 (Feb–Mar 2014) | |
| Uni 5 | Unique detections with 5-min TL intervals. |
| Uni 10 | Unique detections with 10-min TL intervals. |
| TL 5 | TL detections only with 5-min TL intervals. |
| TL 10 | TL detections only with 10-min TL intervals. |
| PIR | PIR detections only matched to interval |
| Scientific Name | Common Name | PIR | Time-Lapse | ||
|---|---|---|---|---|---|
| 1-min | 5-min | 10-min | |||
| Small lizards (SVL ≤ 50 mm) | |||||
| Lampropholis spp.a | Delicate and grass skink | * | * | * | * |
| Saproscincus mustelinus | Weasel skink | * | * | * | |
| Medium–large lizards (SVL > 50 mm) | |||||
| Acritoscincus platynota | Red-throated skink | * | |||
| Amphibolurus muricatus | Jacky dragon | * | * | * | * |
| Ctenotus taeniolatus | Copper-tailed skink | * | * | * | * |
| Cyclodomorphus michaeli | Mainland she-oak skink | * | |||
| Tiliqua scincoides | Blue-tongue skink | * | |||
| Snakes | |||||
| Hemiaspis signata | Marsh snake | * | * | * | |
| Pseudechis porphyriacus | Red-bellied black snake | * | * | * | |
| Pseudonaja textilis | Eastern brown snake | * | * | * | * |
| Group/Scientific Name | Common Name | Reconyx HC600 | Reconyx PC900 | ||
|---|---|---|---|---|---|
| SH | SMh | HH | HMh | ||
| Small lizards (SVL ≤ 50 mm) | |||||
| Lampropholis spp.a | * | ||||
| Lampropholis delicata | Garden skink | NA | NA | * | |
| Lampropholis guichenoti | Grass skink | NA | NA | * | |
| Medium–large lizards (SVL > 50 mm) | |||||
| Amphibolurus muricatus | Jacky dragon | * | * | * | * |
| Ctenotus taeniolatus | Copper-tailed skink | * | * | ||
| Cyclodomorphus michaeli | Mainland she-oak skink | * | |||
| Snakes | |||||
| Pseudechis porphyriacus | Red-bellied black snake | * | * | * | * |
| Pseudonaja textilis | Eastern brown snake | * | * | * | * |
| Small mammals (<500 g) | |||||
| Antechinus stuartii | Brown antechinus | * | * | * | * |
| Cercartetus nanus | Eastern pygmy possum | * | |||
| Rattus fuscipes | Bush rat | * | * | * | * |
| Rattus | Black rat | * | * | * | * |
| Medium–large mammals (≥500 g) | |||||
| Perameles nasuta | Long-nosed bandicoot | * | * | * | * |
| Macropus giganteus | Eastern grey Kangaroo | * | * | * | * |
| Tachyglossus aculeatus | Short-beaked echidna | * | * | * | * |
| Wallabia bicolor | Black wallaby | * | * | * | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Welbourne, D.J.; Claridge, A.W.; Paull, D.J.; Ford, F. Improving Terrestrial Squamate Surveys with Camera-Trap Programming and Hardware Modifications. Animals 2019, 9, 388. https://doi.org/10.3390/ani9060388
Welbourne DJ, Claridge AW, Paull DJ, Ford F. Improving Terrestrial Squamate Surveys with Camera-Trap Programming and Hardware Modifications. Animals. 2019; 9(6):388. https://doi.org/10.3390/ani9060388
Chicago/Turabian StyleWelbourne, D. J., A. W. Claridge, D. J. Paull, and F. Ford. 2019. "Improving Terrestrial Squamate Surveys with Camera-Trap Programming and Hardware Modifications" Animals 9, no. 6: 388. https://doi.org/10.3390/ani9060388