Isolation and Culture of Skin-Derived Differentiated and Stem-Like Cells Obtained from the Arabian Camel (Camelus dromedarius)

 ,
,  ,
,  ,
,  , and
, and 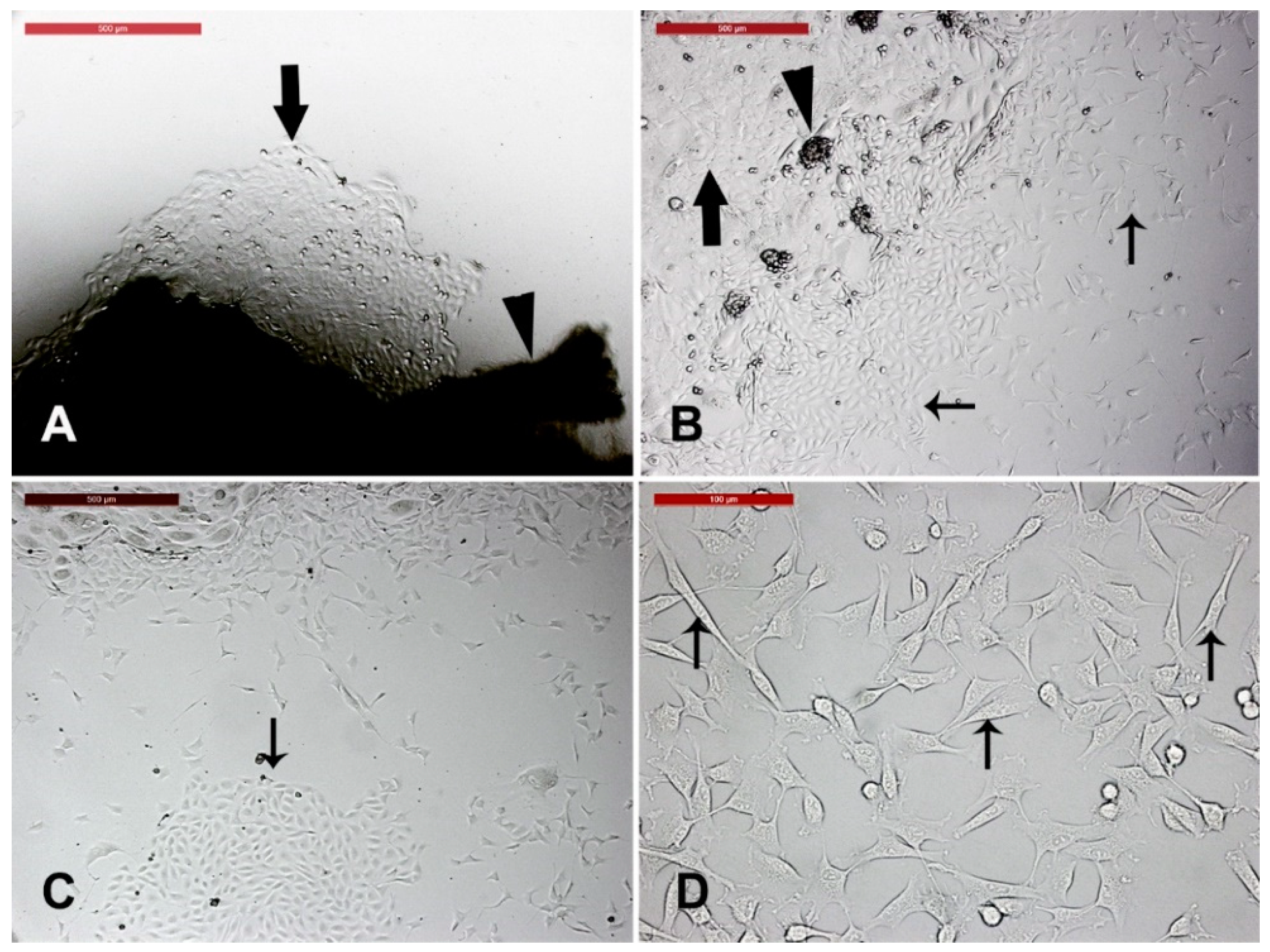{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Sample Collection
2.2. Explant Preparation and Culture
2.3. Monitoring for Outgrowths of Primary Cell Cultures and Different Cell Types
2.4. Characterization of the Observed Cells (Morphology, Trypsinization, Heat Shock, and Osmotic Challenge)
2.5. Cell Differentiation in Dermal Spheroids
2.6. Establishing Secondary Cultures
2.7. Cryopreservation and Resuscitation of Fibroblasts
2.8. Measuring Cell Viability
3. Results
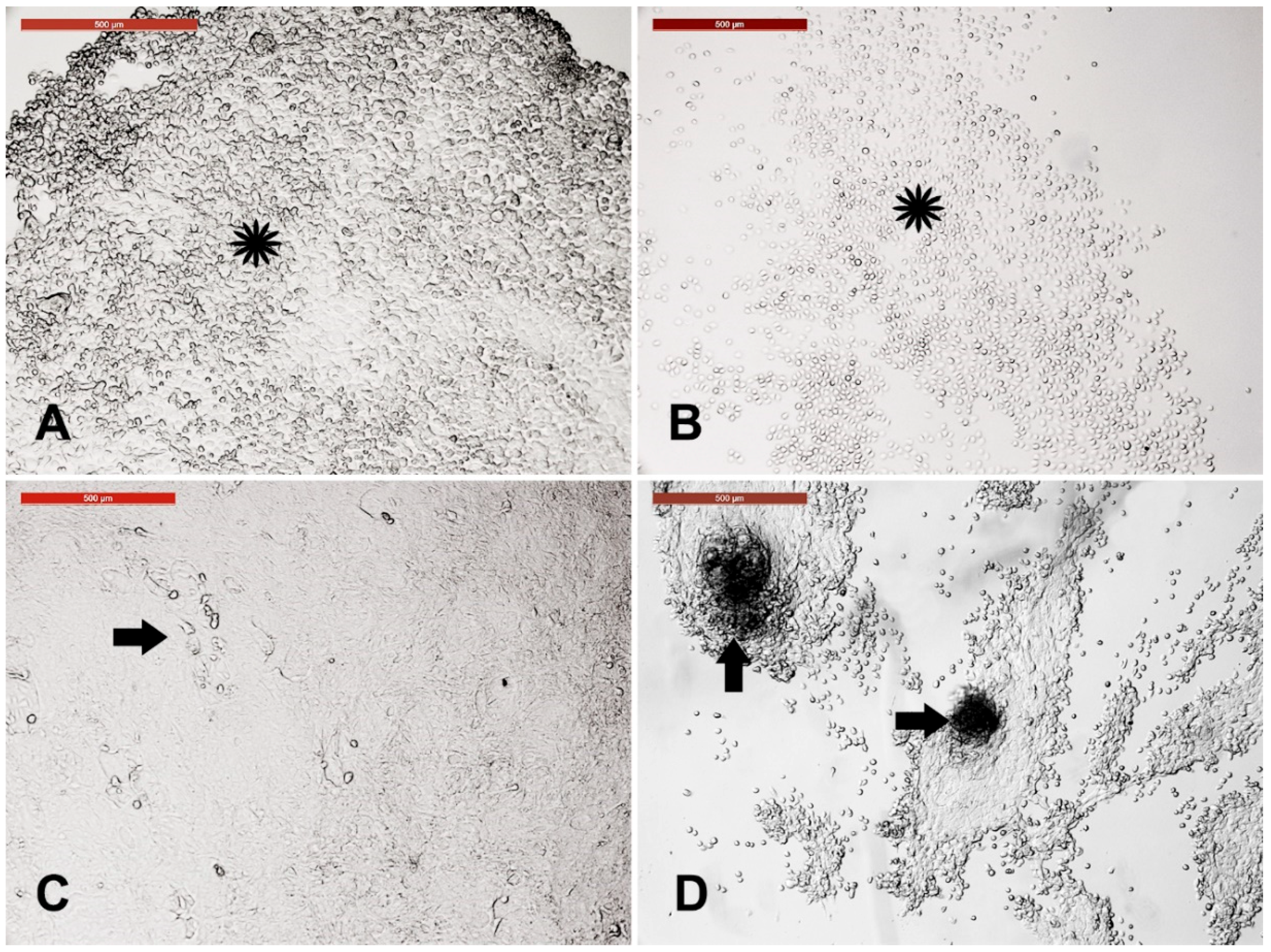
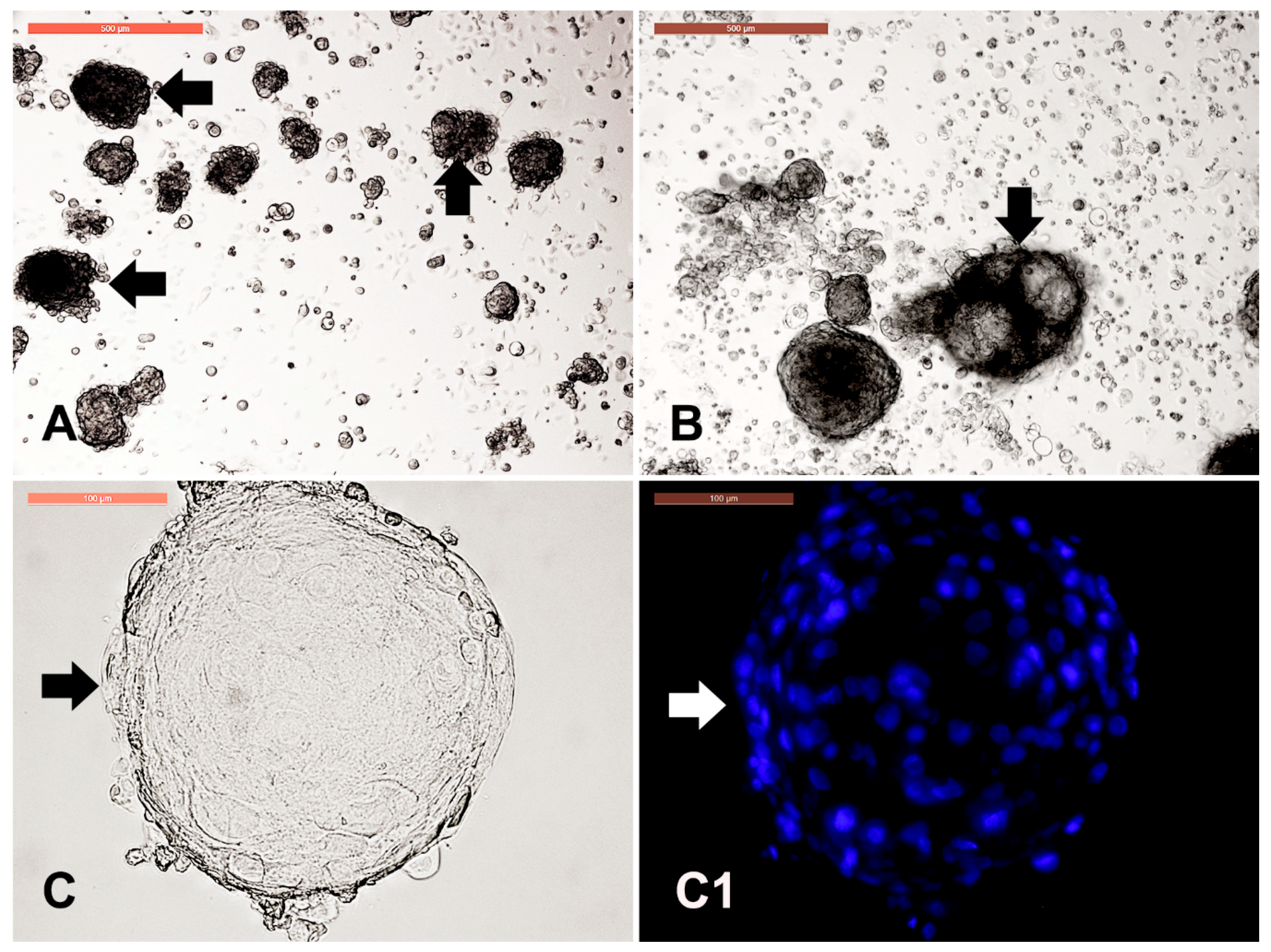
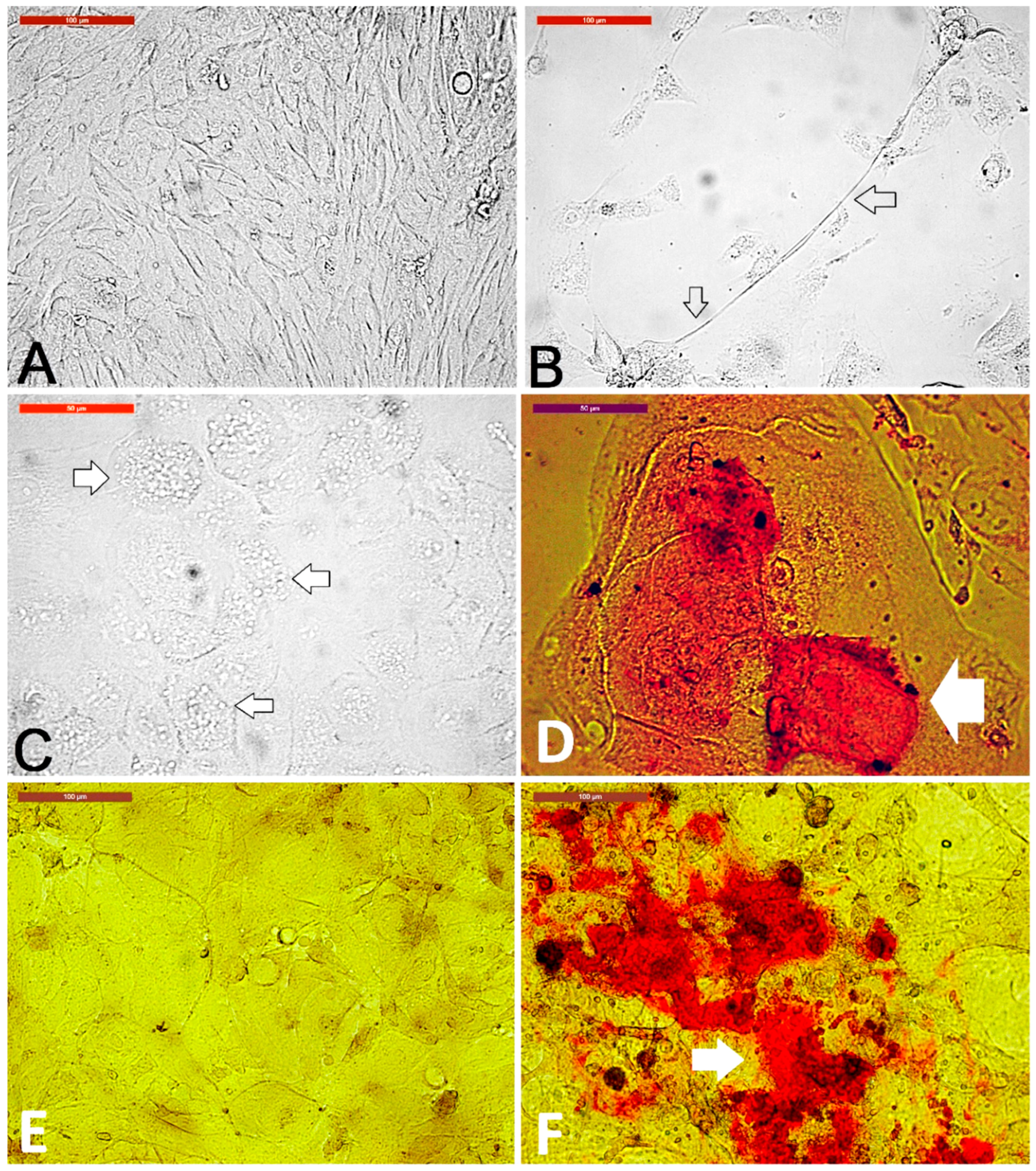
3.1. Camel Skin Explants and Outgrowths at P0 (Passage Zero)
3.2. Long-Term Culture of Camel Skin Explants
3.3. Sensitivity of Different Skin-Derived Cells to Trypsinization
3.4. Sensitivity of Different Trypsin-Sensitive Skin-Derived Cells to Heat Shock
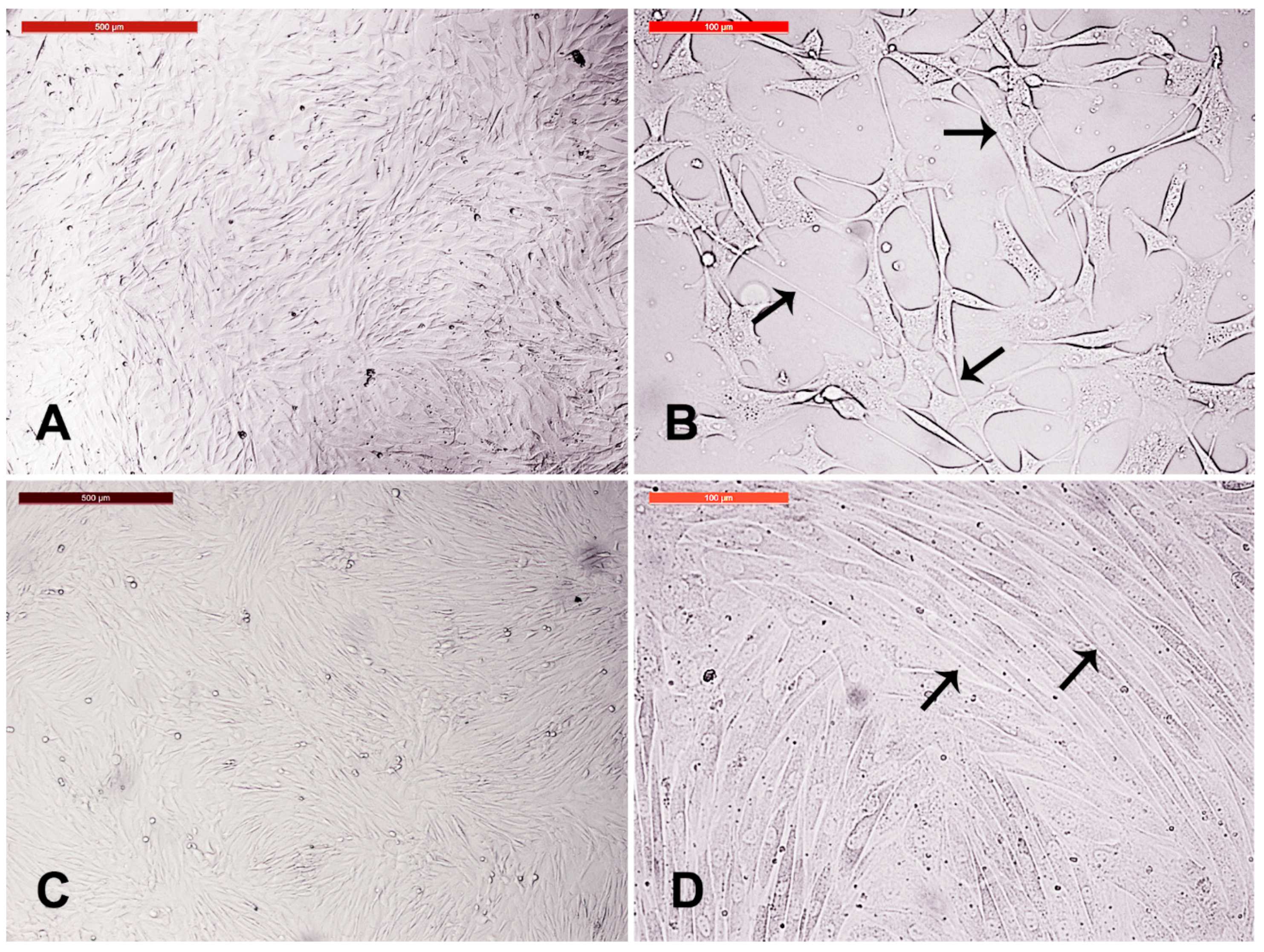
3.5. Fibroblast Subculture and Cryopreservation
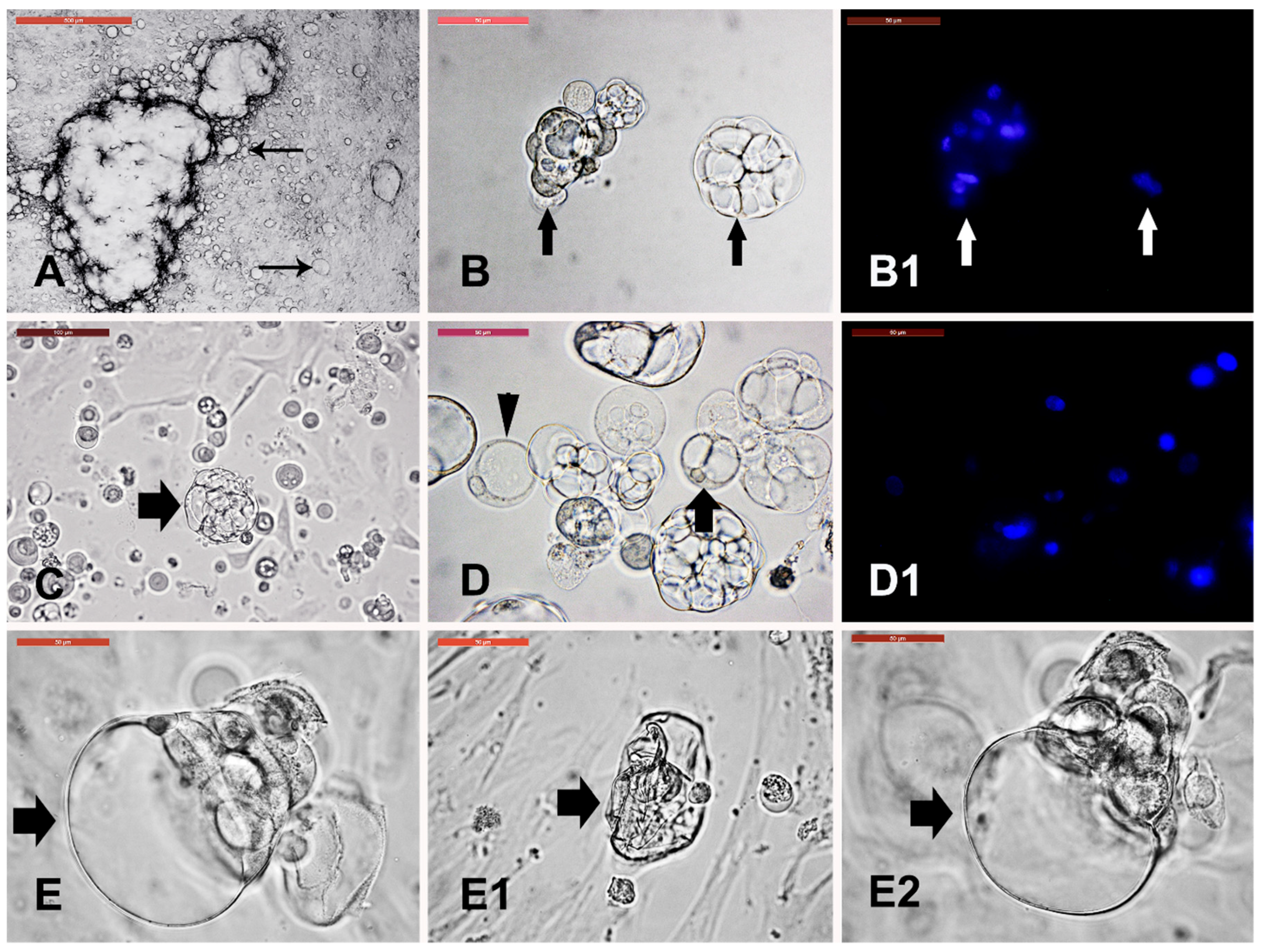
3.6. Cystic Cells Subculture, Cryopreservation, and Characterization
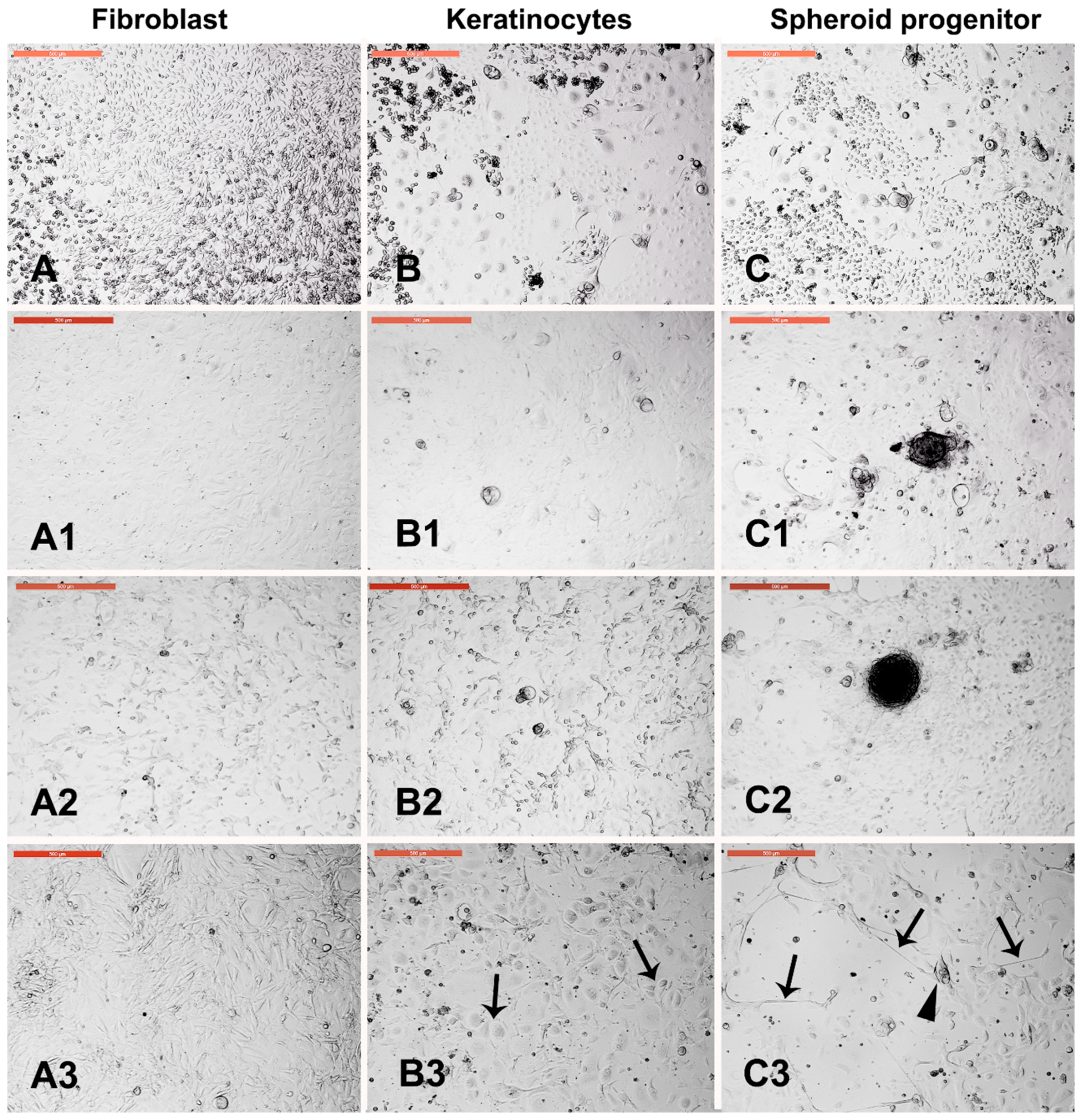
3.7. Dermal Spheroid Characterization
3.8. Dermal Spheroid Differentiation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schmidt-Nielsen, K. The physiology of the camel. Sci. Am. 1959, 201, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Gahlot, T.; Chouhan, D. Fractures in dromedary (Camelus dromedarius)—A retrospective study. J. Camel Pract. Res. 1994, 1, 9–14. [Google Scholar]
- Sheyn, D.; Ben-David, S.; Shapiro, G.; De Mel, S.; Bez, M.; Ornelas, L.; Sahabian, A.; Sareen, D.; Da, X.; Pelled, G. Human induced pluripotent stem cells differentiate into functional mesenchymal stem cells and repair bone defects. Stem Cells Transl. Med. 2016, 5, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Bhakat, C.; Sahani, M. Scope of value addition to camel hide. Nat. Prod. Rad. 2005, 4, 387–390. [Google Scholar]
- Fowler, M. Integumentary System. In Medicine and Surgery of Camelids; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 289–310. [Google Scholar]
- Tarin, D.; Croft, C.B. Ultrastructural features of wound healing in mouse skin. J. Anat. 1969, 105, 189–190. [Google Scholar] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Freitas, B.C.; Trujillo, C.A.; Carromeu, C.; Yusupova, M.; Herai, R.H.; Muotri, A.R. Stem cells and modeling of autism spectrum disorders. Exp. Neurol. 2014, 260, 33–43. [Google Scholar] [CrossRef]
- Ring, K.L.; Tong, L.M.; Balestra, M.E.; Javier, R.; Andrews-Zwilling, Y.; Li, G.; Walker, D.; Zhang, W.R.; Kreitzer, A.C.; Huang, Y. Direct reprogramming of mouse and human fibroblasts into multipotent neural stem cells with a single factor. Cell Stem Cell 2012, 11, 100–109. [Google Scholar] [CrossRef]
- Chang, T.; Zheng, W.; Tsark, W.; Bates, S.; Huang, H.; Lin, R.J.; Yee, J.K. Brief Report: Phenotypic Rescue of Induced Pluripotent Stem Cell-Derived Motoneurons of a Spinal Muscular Atrophy Patient. Stem Cells 2011, 29, 2090–2093. [Google Scholar] [CrossRef]
- Mestre-Citrinovitz, A.C.; Sestelo, A.J.; Ceballos, M.B.; Barañao, J.L.; Saragüeta, P. Isolation of primary fibroblast culture from wildlife: The Panthera onca case to preserve a South American endangered species. Curr. Prot. Mol. Biol. 2016, 116, 28.7.1–28.7.14. [Google Scholar]
- Guan, W.; Liu, C.; Li, C.; Liu, D.; Zhang, W.; Ma, Y. Establishment and cryopreservation of a fibroblast cell line derived from Bengal tiger (Panthera tigris tigris). Cryoletters 2010, 31, 130–138. [Google Scholar] [PubMed]
- Groeneveld, E.; Tinh, N.H.; Kues, W.; Vien, N.T. A protocol for the cryoconservation of breeds by low-cost emergency cell banks—A pilot study. Animal 2008, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Klopries, M.; Wernery, U.; Karden, O.-R. Characterization of the camel skin cell line Dubca. Br. Vet. J. 1995, 151, 555–565. [Google Scholar] [CrossRef]
- Gupta, S.; Gupta, N.; Ahlawat, S.; Kumar, A.; Taneja, R.; Sharma, R.; Sunder, S.; Tantia, M. In Vitro Culture of Skin Fibroblast Cells for Potential Cloning by Nuclear Transfer. In Applications of Gene-Based Technologies for Improving Animal Production and Health in Developing Countries; Springer: Berlin/Heidelberg, Germany, 2005; pp. 631–640. [Google Scholar]
- Sharma, R.; Sharma, H.; Ahlawat, S.; Aggarwal, R.; Vij, P.; Tantia, M. First attempt on somatic cell cryopreservation of critically endangered Camelus bactrianus of India. Gene Rep. 2018, 10, 109–115. [Google Scholar] [CrossRef]
- Wani, N.A.; Hong, S.B. Source, treatment and type of nuclear donor cells influences in vitro and in vivo development of embryos cloned by somatic cell nuclear transfer in camel (Camelus dromedarius). Theriogenology 2018, 106, 186–191. [Google Scholar] [CrossRef]
- Zhou, H.; Guo, Z. Heterogeneous nuclear-transferred-embryos reconstructed with camel (Camelus bactrianus) skin fibroblasts and enucleated ovine oocytes and their development HM. Anim. Reprod. Sci. 2006, 95, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med. 2013, 19, 35–42. [Google Scholar] [CrossRef]
- Sipp, D.; Robey, P.G.; Turner, L. Clear up this stem-cell mess. Nature 2018, 561, 455–457. [Google Scholar] [CrossRef]
- Nadri, S.; Soleimani, M. Isolation murine mesenchymal stem cells by positive selection. In Vitro Cell. Dev. Biol. 2007, 43, 276–282. [Google Scholar] [CrossRef]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; Schwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.R.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedjafari, E.; Soleimani, M.; Ghaemi, N.; Shabani, I. Nanohydroxyapatite-coated electrospun poly(l-lactide) nanofibers enhance osteogenic differentiation of stem cells and induce ectopic bone formation. Biomacromolecules 2010, 11, 3118–3125. [Google Scholar] [CrossRef] [PubMed]
- Kisiel, A.H.; McDuffee, L.A.; Masaoud, E.; Bailey, T.R.; Esparza Gonzalez, B.P.; Nino-Fong, R. Isolation, characterization, and in vitro proliferation of canine mesenchymal stem cells derived from bone marrow, adipose tissue, muscle, and periosteum. Am. J. Vet. Res. 2012, 73, 1305–1317. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Swelum, A.A.-A.; Elsafadi, M.; Mahmood, A.; Alfayez, M.; Alowaimer, A.N. Cumulus cells of camel (Camelus dromedarius) antral follicles are multipotent stem cells. Theriogenology 2018, 118, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Sangcheshmeh, A.; Shafiee, A.; Seyedjafari, E.; Dinarvand, P.; Toghdory, A.; Bagherizadeh, I.; Schellander, K.; Cinar, M.U.; Soleimani, M. Isolation, characterization, and mesodermic differentiation of stem cells from adipose tissue of camel (Camelus dromedarius). In Vitro Cell. Dev. Biol. 2013, 49, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Swelum, A.A.-A.; Elsafadi, M.; Moumen, A.F.; Alzahrani, F.A.; Mahmood, A.; Alfayez, M.; Alowaimer, A.N. Isolation and characterization of the trophectoderm from the Arabian camel (Camelus dromedarius). Placenta 2017, 57, 113–122. [Google Scholar] [CrossRef]
- Wong, V.W.; Levi, B.; Rajadas, J.; Longaker, M.T.; Gurtner, G.C. Stem cell niches for skin regeneration. Int. J. Biomater. 2012, 2012, 926059. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, E.K. Melanocyte stem cells: A melanocyte reservoir in hair follicles for hair and skin pigmentation. Pigment. Cell Melanoma Res. 2011, 24, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fukunaga-Kalabis, M.; Yu, H.; Xu, X.; Kong, J.; Lee, J.T.; Herlyn, M. Human dermal stem cells differentiate into functional epidermal melanocytes. J. Cell Sci. 2010, 123, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Dyce, P.W.; Zhu, H.; Craig, J.; Li, J. Stem cells with multilineage potential derived from porcine skin. Biochem. Biophys. Res. Commun. 2004, 316, 651–658. [Google Scholar] [CrossRef]
- Sun, T.; Yu, C.; Gao, Y.; Zhao, C.; Hua, J.; Cai, L.; Guan, W.; Ma, Y. Establishment and biological characterization of a dermal mesenchymal stem cells line from bovine. Biosci. Rep. 2014, 34, e00101. [Google Scholar] [CrossRef]
- Braun, J.; Hack, A.; Weis-Klemm, M.; Conrad, S.; Treml, S.; Kohler, K.; Walliser, U.; Skutella, T.; Aicher, W.K. Evaluation of the osteogenic and chondrogenic differentiation capacities of equine adipose tissue-derived mesenchymal stem cells. Am. J. Vet. Res. 2010, 71, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Al-Sobayil, F. Circadian rhythm of bone formation biomarkers in serum of dromedary camels. Res. Vet. Sci. 2010, 89, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Vangipuram, M.; Ting, D.; Kim, S.; Diaz, R.; Schüle, B. Skin punch biopsy explant culture for derivation of primary human fibroblasts. J. Vis. Exp. 2013, 7, e3779. [Google Scholar] [CrossRef] [PubMed]
- Reiisi, S.; Esmaeili, F.; Shirazi, A. Isolation, culture and identification of epidermal stem cells from newborn mouse skin. In Vitro Cell. Dev. Biol. 2010, 46, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, B.; Kobayasi, T. Separation of human epidermal cells from fibroblasts in primary skin culture. Arch. Dermatol. Res. 1975, 252, 39–46. [Google Scholar] [CrossRef]
- Louis, K.S.; Siegel, A.C. Cell viability analysis using trypan blue: Manual and automated methods. In Mammalian Cell Viability; Springer: Berlin/Heidelberg, Germany, 2011; pp. 7–12. [Google Scholar]
- Fernandes, K.J.; McKenzie, I.A.; Mill, P.; Smith, K.M.; Akhavan, M.; Barnabé-Heider, F.; Biernaskie, J.; Junek, A.; Kobayashi, N.R.; Toma, J.G.; et al. A dermal niche for multipotent adult skin-derived precursor cells. Nat. Cell Biol. 2004, 6, 1082–1093. [Google Scholar] [CrossRef] [PubMed]
- Iribar, H.; Castro-Feo, B.; Azcoitia-Ramsden, I.; de Paz-Alonso, N.; Izeta, A.; Gutiérrez-Rivera, A. Optimization of the porcine adult skin-derived precursor cell isolation protocol. Stem Cell Discov. 2013, 3, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Toma, J.G.; McKenzie, I.A.; Bagli, D.; Miller, F.D. Isolation and characterization of multipotent skin-derived precursors from human skin. Stem Cells 2005, 23, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.M.; Kajiura, S.; Cao, W.; Liu, F.; Amoh, Y. Cryopreservation of Hair-Follicle Associated Pluripotent (HAP) Stem Cells Maintains Differentiation and Hair-Growth Potential. In Biobanking and Cryopreservation of Stem Cells; Karimi-Busheri, F., Weinfeld, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 191–198. [Google Scholar]
- Tsuji, K.; Ojima, M.; Otabe, K.; Horie, M.; Koga, H.; Sekiya, I.; Muneta, T. Effects of Different Cell-Detaching Methods on the Viability and Cell Surface Antigen Expression of Synovial Mesenchymal Stem Cells. Cell Transplant. 2017, 26, 1089–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernerd, F.; Schweizer, J.; Demarchez, M. Dermal cysts of the rhino mouse develop into unopened sebaceous glands. Arch. Dermatol. Res. 1996, 288, 586–595. [Google Scholar] [CrossRef]
- Nørgaard, R.; Kassem, M.; Rattan, S.I.S. Heat Shock–Induced Enhancement of Osteoblastic Differentiation of hTERT-Immortalized Mesenchymal Stem Cells. Ann. N. Y. Acad. Sci. 2006, 1067, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, Z.-D.; Ji, X.; Morales, J.; Zhang, J.; Kaur, N.; Wang, S. Enhanced Osteogenesis of Human Mesenchymal Stem Cells by Periodic Heat Shock in Self-Assembling Peptide Hydrogel. Tissue Eng. Part A 2013, 19, 716–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Upadhyay, R.; Malakar, D.; Kumar, S.; Singh, S. Effect of thermal stress on HSP70 expression in dermal fibroblast of zebu (Tharparkar) and crossbred (Karan-Fries) cattle. J. Thermal Biol. 2014, 43, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Saadeldin, I.M.; Swelum, A.A.-A.; Elsafadi, M.; Mahmood, A.; Alfayez, M.; Alowaimer, A.N. Differences between the tolerance of camel oocytes and cumulus cells to acute and chronic hyperthermia. J. Therm. Biol. 2018, 74, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, G., Jr.; Yoo, J.J.; De Coppi, P.; Siddiqui, M.M.; Schuch, G.; Pohl, H.G.; Fuhr, J.; Perin, L.; Soker, S.; Atala, A. Propagation, expansion, and multilineage differentiation of human somatic stem cells from dermal progenitors. Stem Cells Dev. 2005, 14, 337–348. [Google Scholar] [CrossRef]
- Chen, F.G.; Zhang, W.J.; Bi, D.; Liu, W.; Wei, X.; Chen, F.F.; Zhu, L.; Cui, L.; Cao, Y. Clonal analysis of nestin(−) vimentin(+) multipotent fibroblasts isolated from human dermis. J Cell Sci. 2007, 120, 2875–2883. [Google Scholar] [CrossRef] [PubMed]
- Zong, Z.; Li, N.; Ran, X.; Su, Y.; Shen, Y.; Shi, C.-M.; Cheng, T.-M. Isolation and characterization of two kinds of stem cells from the same human skin back sample with therapeutic potential in spinal cord injury. PLoS ONE 2012, 7, e50222. [Google Scholar] [CrossRef] [PubMed]
- Volkova, N.A.; Mazur, S.P.; Kholodnyy, V.S.; Petrenko, A.Y. Skin Stem Cells as an Object for Cryopreservation. 1. Skin Stem Reserve. Probl. Cryobiol. Cryomed. 2014, 24, 3–15. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Mistriotis, P.; Andreadis, S.T. Clonal multipotency and effect of long-term in vitro expansion on differentiation potential of human hair follicle derived mesenchymal stem cells. Stem Cell Res. 2012, 8, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Uchugonova, A.; Kimura, H.; Zhang, C.; Zhao, M.; Zhang, L.; Koenig, K.; Duong, J.; Aki, R.; Saito, N. The bulge area is the major hair follicle source of nestin-expressing pluripotent stem cells which can repair the spinal cord compared to the dermal papilla. Cell Cycle 2011, 10, 830–839. [Google Scholar] [CrossRef] [Green Version]
- Linher, K.; Dyce, P.; Li, J. Primordial germ cell-like cells differentiated in vitro from skin-derived stem cells. PLoS ONE 2009, 4, e8263. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-T.; Bennett, M.W.; Zhang, X.; Spate, L.; Whitworth, K.M.; Murphy, C.N.; Rieke, A.; Zhang, Y.; Prather, R.S. Contribution to neural and mesodermal lineages by porcine skin-derived progenitors (SKPs) in vivo. Cell Cycle 2010, 9, 2040–2041. [Google Scholar] [CrossRef]
- Lavoie, J.-F.; Biernaskie, J.A.; Chen, Y.; Bagli, D.; Alman, B.; Kaplan, D.R.; Miller, F.D. Skin-derived precursors differentiate into skeletogenic cell types and contribute to bone repair. Stem Cells Dev. 2009, 18, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-T.; Prather, R.S. The multi-potentiality of skin-derived stem cells in pigs. Theriogenology 2011, 75, 1372–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Nbaheen, M.; Ali, D.; Bouslimi, A.; Al-Jassir, F.; Megges, M.; Prigione, A.; Adjaye, J.; Kassem, M.; Aldahmash, A. Human stromal (mesenchymal) stem cells from bone marrow, adipose tissue and skin exhibit differences in molecular phenotype and differentiation potential. Stem Cell Rev. 2013, 9, 32–43. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saadeldin, I.M.; Swelum, A.A.-A.; Noreldin, A.E.; Tukur, H.A.; Abdelazim, A.M.; Abomughaid, M.M.; Alowaimer, A.N. Isolation and Culture of Skin-Derived Differentiated and Stem-Like Cells Obtained from the Arabian Camel (Camelus dromedarius). Animals 2019, 9, 378. https://doi.org/10.3390/ani9060378
Saadeldin IM, Swelum AA-A, Noreldin AE, Tukur HA, Abdelazim AM, Abomughaid MM, Alowaimer AN. Isolation and Culture of Skin-Derived Differentiated and Stem-Like Cells Obtained from the Arabian Camel (Camelus dromedarius). Animals. 2019; 9(6):378. https://doi.org/10.3390/ani9060378
Chicago/Turabian StyleSaadeldin, Islam M., Ayman Abdel-Aziz Swelum, Ahmed E. Noreldin, Hammed A. Tukur, Aaser M. Abdelazim, Mosleh M. Abomughaid, and Abdullah N. Alowaimer. 2019. "Isolation and Culture of Skin-Derived Differentiated and Stem-Like Cells Obtained from the Arabian Camel (Camelus dromedarius)" Animals 9, no. 6: 378. https://doi.org/10.3390/ani9060378