Effect of Soybean Isoflavones on Growth Performance, Immune Function, and Viral Protein 5 mRNA Expression in Broiler Chickens Challenged with Infectious Bursal Disease Virus

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds, Virus, and Diets
2.2. Experimental Design
2.3. Growth Performance
2.4. Blood Sampling and Laboratory Analyses
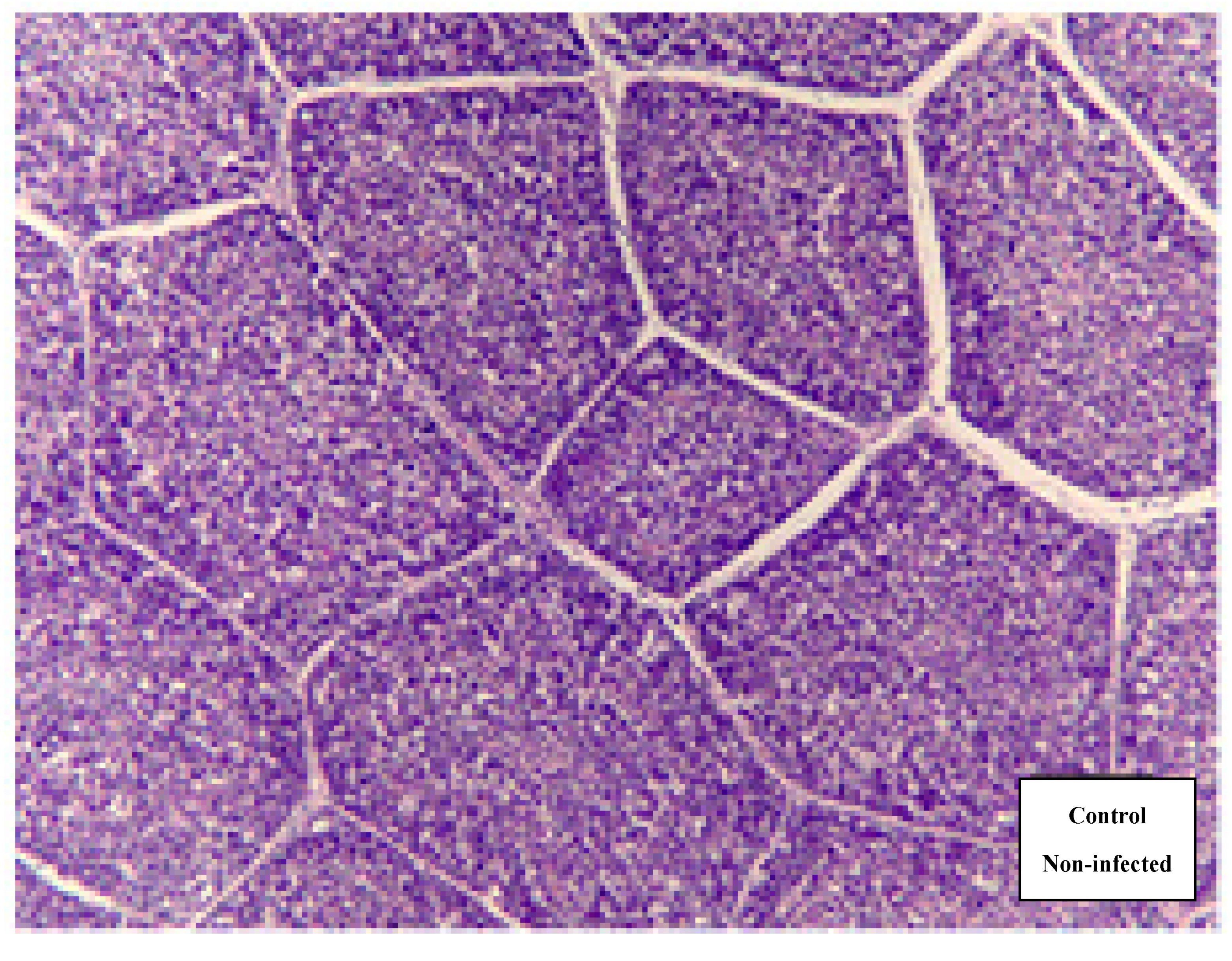
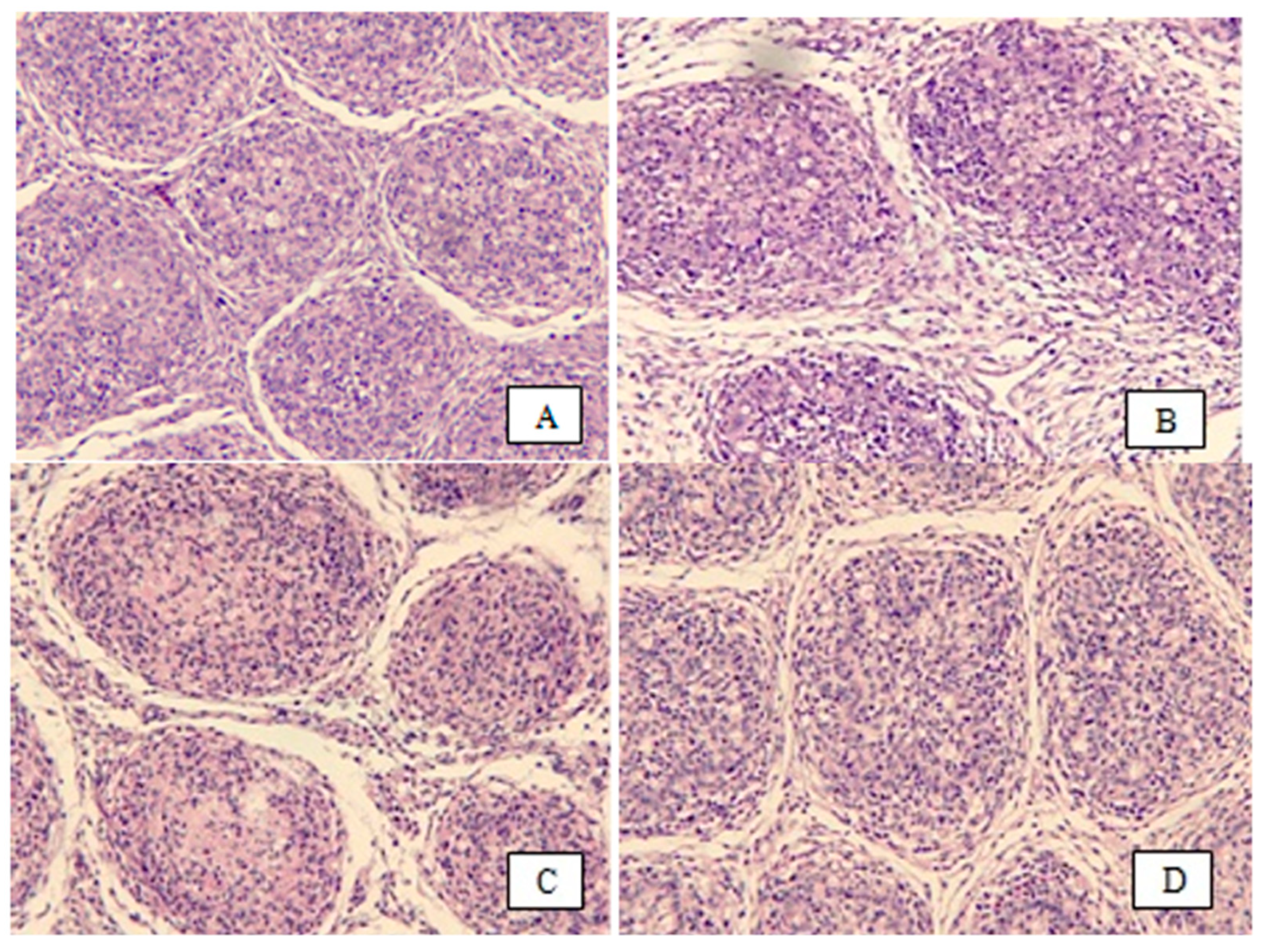
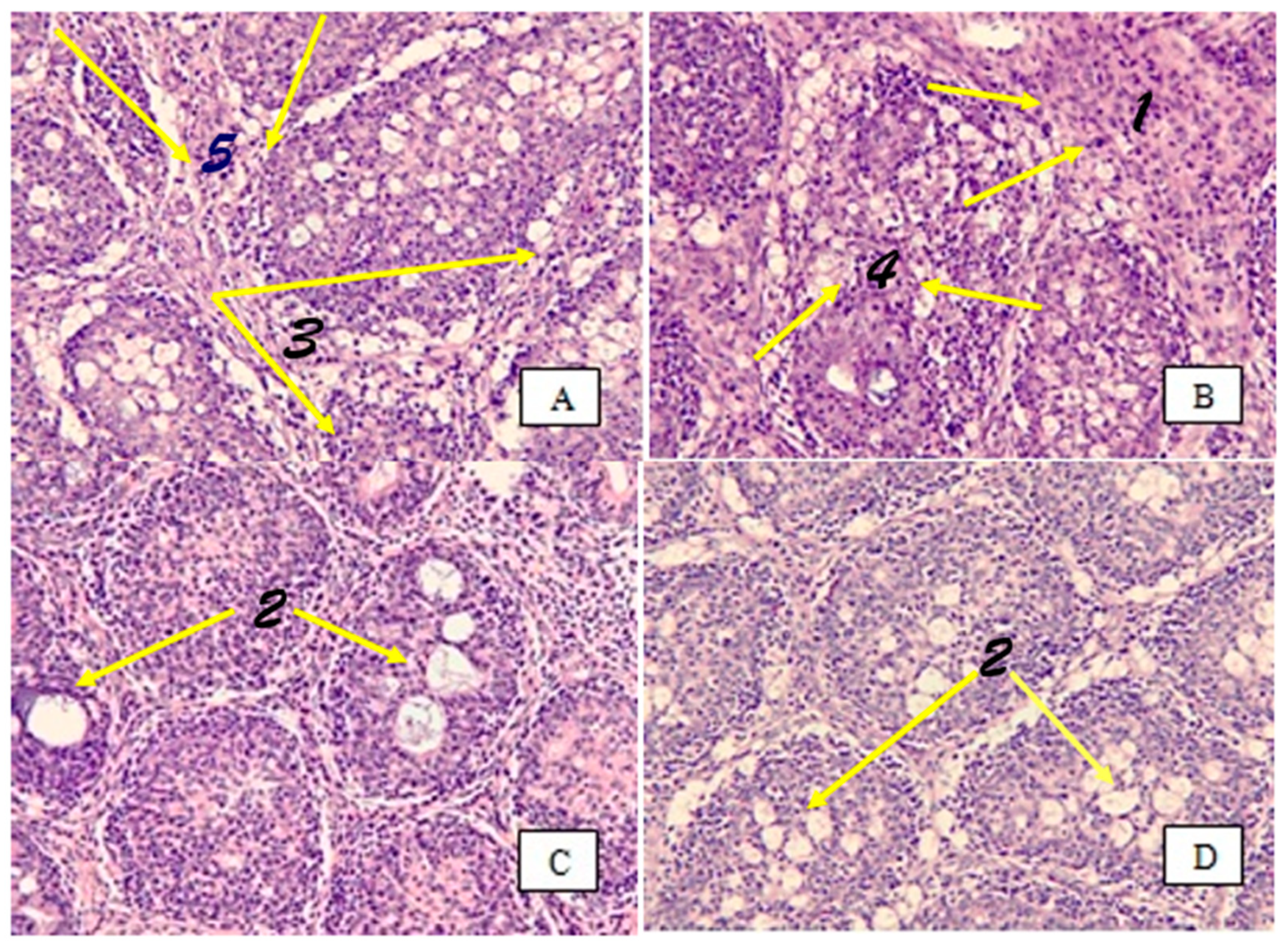
2.5. Histology of Bursa of Fabricius
2.6. Viral Protein 5 (VP5) mRNA Expression
2.7. Determination of Antioxidant Capacity in the Serum
2.8. Determination of Bursal Immunologic Indices
2.9. Statistical Analyses
3. Results and Discussions
3.1. Bursa of Fabricius
3.2. Viral Protein 5 (VP5) mRNA Expression
3.3. Immune Function
3.4. Antioxidant Capacity and Oxidative Stress
3.5. Growth Performance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mazariegos, L.A.; Lukert, P.D.; Brown, J. Pathogenicity and immunosuppressive properties of infectious bursal disease “intermediate” strains. Avian Dis. 1990, 34, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—The past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef]
- Balamurugan, V.; Kataria, J.M. Economically important non-oncogenic immunosuppressive viral diseases of chicken—Current status. Vet. Res. Commun. 2006, 30, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Wang, Y.; Yu, J.; Hu, S. Enhanced immune responses of chickens to oral vaccination against infectious bursal disease by ginseng stem-leaf saponins. Poult. Sci. 2014, 93, 2473–2481. [Google Scholar] [CrossRef] [Green Version]
- Giambrone, J.J.; Clay, R.P. Isolation of variant infectious bursal disease viruses from commercial broiler chickens. Poult. Sci. 1986, 65, 47. [Google Scholar]
- Fussell, L.W. Poultry industry strategies for control of immunosuppressive diseases. Poult. Sci. 1998, 77, 1193–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butcher, G.D.; Miles, R.D. Interrelationship of Nutrition and Immunity. University of Florida Cooperative Extension Service; Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2002. [Google Scholar]
- Abdukalykova, S.T.; Zhao, X.; Ruiz-Feria, C.A. Arginine and vitamin E modulate the subpopulations of T lymphocytes in broiler chickens. Poult. Sci. 2008, 87, 50–55. [Google Scholar] [CrossRef]
- Azzam, M.M.M.; Zou, X.T.; Dong, X.Y.; Xie, P. Effect of supplemental L-threonine on mucin 2 gene expression and intestine mucosal immune and digestive enzymes activities of laying hens in environments with high temperature and humidity. Poult. Sci. 2011, 90, 2251–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.Y.; Azzam, M.M.M.; Zou, X.T. Effects of dietary threonine supplementation on intestinal barrier function and gut microbiota of laying hens. Poult. Sci. 2017, 96, 3654–3663. [Google Scholar] [CrossRef]
- Messina, M. Soy and health update: Evaluation of the clinical and epidemiologic literature. Nutrients 2016, 8, 754. [Google Scholar] [CrossRef]
- Rietjens, I.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef]
- Setchell, K.D. Phytoestrogens: The biochemistry, physiology, and implications for human health of soy isoflavones. Am. J. Clin. Nutr. 1998, 68, 1333S–1346S. [Google Scholar] [CrossRef]
- Wei, H.; Bowen, R.; Cai, Q.; Barnes, S.; Wang, Y. Antioxidant and antipromotional effects of the soybean isoflavonegenistein. Proc. Soc. Exp. Biol. Med. 1995, 208, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Larrea, M.B.; Mohan, A.R.; Paganga, G.; Miller, N.J.; Bolwell, G.P.; Rice-Evans, C.A. Antioxidant activity of phytoestrogenicisoflavones. Free Radic. Res. 1997, 26, 63–70. [Google Scholar] [CrossRef]
- Liu, J.; Chang, S.K.; Wiesenborn, D. Antioxidant properties of soybean isoflavone extract and tofu in vitro and in vivo. J. Agric. Food Chem. 2005, 53, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Jiang, S.Q.; Lin, Y.C.; Xi, P.B.; Yu, D.Q.; Wu, T.X. Effects of soybean isoflavone on growth performance, meat quality, and antioxidation in male broilers. Poult. Sci. 2007, 86, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Dixit, A.K.; Bhatnagar, D.; Kumar, V.; Chawla, D.; Fakhruddin, K.; Bhatnagar, D. Antioxidant potential and radioprotective effect of soy isoflavone against gamma irradiation induced oxidative stress. J. Funct. Foods 2012, 4, 197–206. [Google Scholar] [CrossRef]
- Huang, L.; Ma, X.Y.; Jiang, Z.Y.; Hu, Y.J.; Zheng, C.T.; Yang, X.F.; Wang, L.; Gao, K.G. Effects of soybean isoflavone on intestinal antioxidant capacity and cytokines in young piglets fed oxidized fish oil. J. Zhejiang Univ. Sci. B 2016, 17, 965–974. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Poultry, 9th rev. ed.; National Academies Press: Washington, DC, USA, 1994.
- Sharma, J.M.; Dohms, J.E.; Metz, A.L. Comparative pathogenesis of serotype 1 and variant serotype 1 isolates of infectious bursal disease virus and their effect on humoral and cellular immune competence of specific-pathogen-free chickens. Avian Dis. 1989, 33, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.J.; Gagic, M.; Sharma, J.M. Recovery of antibody-producing ability and lymphocyte repopulation of bursal follicles in chickens exposed to infectious bursal disease virus. Avian Dis. 1999, 43, 401–413. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Li, L.; Kubasova, T.; Rychlik, I.; Hoerr, F.J.; Rautenschlein, S. Infectious bursal disease virus infection leads to changes in the gut associated-lymphoid tissue and the microbiota composition. PLoS ONE 2018, 13, e0192066. [Google Scholar] [CrossRef] [PubMed]
- Eterradossi, N.; Saif, Y.M. Infectious bursal disease. Dis. Poult. 2008, 12, 185–208. [Google Scholar]
- Eterradossi, N.; Saif, Y.M. Infectious bursal disease. In Diseases of Poultry, 13rd ed.; Swayne, D.E., Ed.; Wiley: Ames, IA, USA, 2013. [Google Scholar]
- Sharma, J.M.; Kim, I.J.; Rautenschlein, S.; Yeh, H.Y. Infectious bursal disease virus of chickens: Pathogenesis and immunosuppression. Dev. Comp. Immunol. 2000, 24, 223–235. [Google Scholar] [CrossRef]
- Abdel-Alim, G.A.; Saif, Y.M. Immunogenicity and antigenicity of very virulent strains of infectious bursal disease viruses. Avian Dis. 2001, 29, 92–101. [Google Scholar] [CrossRef]
- Abdel-Alim, G.A.; Awaad, M.H.H.; Saif, Y.M. Characterization of Egyptian field strains of infectious bursal disease virus. Avian Dis. 2003, 47, 1452–1457. [Google Scholar] [CrossRef]
- Lombardo, E.; Maraver, A.; Espinosa, I.; Fernández-Arias, A.; Rodriguez, J.F. VP5, the nonstructural polypeptide of infectious bursal disease virus, accumulates within the host plasma membrane and induces cell lysis. J. Virol. 2000, 277, 345–357. [Google Scholar] [CrossRef]
- Liu, M.; Vakharia, V.N. Nonstructural protein of infectious bursal disease virus inhibits apoptosis at the early stage of virus infection. J. Virol. 2006, 80, 3369–3377. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zheng, S. Infectious bursal disease virus-host interactions: Multifunctional viral proteins that perform multiple and differing jobs. Int. J. Mol. Sci. 2017, 18, 161. [Google Scholar] [CrossRef]
- Wei, L.; Hou, L.; Zhu, S.; Wang, J.; Zhou, J.; Liu, J. Infectious bursal disease virus activates the phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway by interaction of VP5 protein with the p85α subunit of PI3K. J. Virol. 2011, 417, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, Y.; Xue, Y.; Li, X.; Cao, H.; Zheng, S.J. Critical role for voltage-dependent anion channel 2 in infectious bursal disease virus-induced apoptosis in host cells via interaction with VP5. J. Virol. 2012, 86, 1328–1338. [Google Scholar] [CrossRef]
- Bonham, M.; O’Connor, J.M.; Hannigan, B.M.; Strain, J.J. The immune system as a physiological indicator of marginal copper status? Br. J. Nutr. 2002, 87, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, F.Y.; Guo, Y.M.; Wang, Z.; Liu, D.; Zhang, B.K.; Yang, X. Conjugated linoleic acids alleviate infectious bursal disease virus-induced immunosuppression in broiler chickens. Poult. Sci. 2011, 90, 1926–1933. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nature 1996, 383, 787–793. [Google Scholar] [CrossRef]
- Ruby, T.; Whittaker, C.; Withers, D.R.; Chelbi-Alix, M.K.; Morin, V.; Oudin, A.; Young, J.R.; Zoorob, R. Transcriptional profiling reveals a possible role for the timing of the inflammatory response in determining susceptibility to a viral infection. J. Virol. 2006, 80, 9207–9216. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy isoflavones and prostate cancer: A review of molecular mechanisms. J. Steroid. Biochem. Mol. Biol. 2014, 140, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Messina, S.; Bitto, A.; Aguennouz, M.H.; Vita, G.L.; Polito, F.; Irrera, N.; Altavilla, D.; Marini, H.; Migliorato, A.; Squadrito, F.; et al. The soy isoflavone genistein blunts nuclear factor kappa-B, MAPKs and TNF-α activation and ameliorates muscle function and morphology in mdx mice. Neuromuscul. Disord. 2011, 21, 579–589. [Google Scholar] [CrossRef]
- Lv, Z.; Fan, H.; Zhang, B.; Xing, K.; Guo, Y. Dietary genistein supplementation for breeders and their offspring improves the growth performance and immune function of broilers. Sci. Rep. 2018, 8, 5161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wu, C. Dietary soy isoflavones alleviate dextran sulfate sodium-induced inflammation and oxidative stress in mice. Exp. Ther. Med. 2017, 14, 276–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, L.L.; Stahly, T.S.; Stabel, T.J. The effect of dietary soy daidzein on pig growth and viral replication during a viral challenge. J. Anim. Sci. 2001, 79, 3113–3119. [Google Scholar] [CrossRef]
- Greiner, L.L.; Stahly, T.S.; Stabel, T.J. The effect of dietary soy genistein on pig growth and viral replication during a viral challenge. J. Anim. Sci. 2001, 79, 1272–1279. [Google Scholar] [CrossRef]
- Aricibasi, M.; Jung, A.; Heller, E.D.; Rautenschlein, S. Differences in genetic background influence the induction of innate and acquired immune responses in chickens depending on the virulence of the infecting infectious bursal disease virus (IBDV) strain. Vet. Immunol. Immunopathol. 2010, 135, 79–92. [Google Scholar] [CrossRef]
- Tippenhauer, M.; Heller, D.E.; Weigend, S.; Rautenschlein, S. The host genotype influences infectious bursal disease virus pathogenesis in chickens by modulation of T cells responses and cytokine gene expression. Dev. Comp. Immunol. 2013, 40, 1–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Composition, g/kg | |
|---|---|---|
| 1–21 Days of Age | 22–42 Days of Age | |
| Maize | 584.0 | 608.0 |
| Wheat bran | 43.0 | 38.0 |
| Fish meal | 22.0 | 10.0 |
| Soybean meal | 264.0 | 220.0 |
| Maize gluten meal | 20.0 | 30.0 |
| Soybean oil | 13.0 | 29.0 |
| Lysine | 0.0 | 1.0 |
| Methionine | 1.0 | 0.8 |
| Limestone | 12.7 | 12.0 |
| Dicalcium phosphate | 15.1 | 14.5 |
| Salt | 2.5 | 2.5 |
| Zeolite | 12.7 | 24.2 |
| Vitamin-mineral premix 1 | 10.0 | 10.0 |
| Total | 1000.0 | 1000.0 |
| Chemical composition, g/kg 2 | ||
| Crude protein | 199.3 | 187.4 |
| Lysine | 10.5 | 9.8 |
| Methionine | 4.6 | 4.0 |
| Calcium | 10.0 | 9.0 |
| Non-phytate P | 4.5 | 4.0 |
| Metabolizable Energy, MJ/kg | 12.13 | 12.55 |
| Treatments | Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) |
|---|---|---|---|---|---|
| SI, mg/kg | 0 | 0 | 10 | 20 | 40 |
| IBDV | − | + | + | + | + |
| Gene name | Primer Sequence | Amplicon Size [bp] |
|---|---|---|
| VP5 1 | F:5′-GAGCCTTCTGATGCCAACAAC-3′ R:5′-CAAATTGTAGGTCGAGGTCTCTGA-3′ | 94 bp |
| β-actin | F:5′-TGGCATTGCTGACAGGAT-3′ R:5′-CTGCTTGCTGATCCACAT-3′ | 160 bp |
| Indices | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| Bursa weight, g | 3days PI | 2.45 ± 0.15 A | 1.60 ± 0.05 B | 1.88 ± 0.18 | 1.88 ± 0.17 | 1.80 ± 0.10 |
| 7days PI | 2.25 ± 0.22 A | 0.58 ± 0.06 Bb | 0.87 ± 0.05 a | 0.63 ± 0.08 b | 0.59 ± 0.05 b | |
| Bursa index, % | 3days PI | 0.46 ± 0.03 A | 0.32±0.02 B | 0.34 ± 0.04 | 0.36 ± 0.04 | 0.35 ± 0.02 |
| 7days PI | 0.32 ± 0.02 A | 0.10±0.01 B | 0.14 ± 0.00 | 0.13 ± 0.02 | 0.10 ± 0.01 | |
| Indices | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| Bursa Score | 3days PI | 0.00±0.00 B | 4.00 ± 0.00 Aa | 3.63 ± 0.26 ab | 3.13 ± 0.30 b | 3.75 ± 0.16 ab |
| 7days PI | 0.00±0.00 B | 3.88 ± 0.13 Aa | 4.00 ± 0.00a | 3.13 ± 0.30 b | 3.86 ± 0.14 a | |
| Indices | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| viral protein 5 [VP5] | 3days PI | 0.00 ± 0.00 B | 0.0554 ± 0.0211 Aa | 0.0492 ± 0.0080 a | 0.0052 ± 0.0026 b | 0.0548 ± 0.0197 a |
| 7days PI | 0.00 ± 0.00 B | 0.0025 ± 0.0005 Aa | 0.0012 ± 0.0003 b | 0.0010 ± 0.0004 b | 0.0034 ± 0.0004 a | |
| Indices | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| IL-2, pg/mL | 3d PI | 106.52 ± 5.55 | 99.90 ± 3.34 b | 104.05 ± 3.91 ab | 111.90 ± 2.99 a | 113.32 ± 3.43 a |
| 7d PI | 216.07 ± 14.27 | 201.13 ± 9.86 b | 220.39 ± 15.57 b | 195.21 ± 9.47b | 270.04 ± 24.25 a | |
| IL-6, pg/mL | 3d PI | 50.22 ± 5.62 | 40.24 ± 3.14 b | 53.61 ± 4.78 a | 48.29 ± 2.78 ab | 47.41 ± 2.53 ab |
| 7d PI | 33.71 ± 4.47 B | 56.78 ± 6.02 A | 47.58 ± 5.09 | 51.43 ± 2.87 | 55.31 ± 3.24 | |
| IFNγ, ng/mL | 3d PI | 7.95 ± 1.05 B | 12.94 ± 0.85 Aa | 9.69 ± 0.52 b | 10.34 ± 1.34 ab | 9.24 ± 1.06 b |
| 7d PI | 5.58 ± 0.24 B | 7.64 ± 0.96 A | 8.53 ± 0.38 | 6.49 ± 0.89 | 6.38 ± 1.14 | |
| NF-κB, pg/mL | 3d PI | 54.26 ± 8.67 | 50.64 ± 7.29 | 58.39 ± 9.80 | 67.52 ± 7.29 | 50.50 ± 6.90 |
| 7d PI | 58.91 ± 1.11 B | 113.08 ± 9.50 Aa | 68.93 ± 6.42 b | 80.56 ± 13.31 b | 92.85 ± 8.93 ab | |
| Indices 2 | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| TAOC, U/mL | 3days PI | 9.86 ± 0.36 A | 4.20 ± 0.20 Bb | 7.30 ± 0.33 a | 6.63 ± 0.54 a | 6.84 ± 0.45 a |
| 7days PI | 7.70 ± 0.63 | 7.60 ± 0.61 | 7.24 ± 0.49 | 7.30 ± 0.43 | 6.66 ± 0.52 | |
| MDA, mol/mL | 3days PI | 5.19 ± 0.57 | 4.12 ± 0.55 | 5.21 ± 0.81 | 4.40 ± 0.98 | 3.18 ± 0.68 |
| 7days PI | 4.33 ± 0.21 | 4.45 ± 0.75 a | 3.37 ± 0.57 ab | 2.45 ± 0.30 b | 2.07 ± 0.26 b | |
| Indices | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | IBDV (0 SI) | IBDV (10 SI) | IBDV (20 SI) | IBDV (40 SI) | ||
| BW, g | 1 | 39.55 ± 0.19 | 39.75 ± 0.17 | 40.00 ± 0.07 | 39.93 ± 0.20 | 39.70 ± 0.14 |
| 21 | 455.00 ± 2.04 | 452.50 ± 14.22 ab | 473.75 ± 5.54 a | 460.00 ± 2.89 ab | 436.25 ± 9.87 b | |
| 42 | 1351.50 ± 16.05 | 1288.04 ± 27.34 ab | 1320.58 ± 13.98 a | 1332.06 ± 23.64 a | 1228.58 ± 40.72 b | |
| Average daily gain, g | 1–21 | 20.77 ± 0.10 | 20.64 ± 0.71 ab | 21.69 ± 0.28 a | 21.09 ± 0.11 ab | 19.83 ± 0.49 b |
| 22–42 | 40.75 ± 0.79 A | 37.98 ± 0.73 B | 38.49 ± 0.66 | 39.57 ± 1.04 | 36.02 ± 1.69 | |
| 1–42 | 31.23 ± 0.39 | 29.72 ± 0.65 ab | 30.49 ± 0.33 a | 30.76 ± 0.56 a | 28.31 ± 0.97 b | |
| ADFI, g | 1–21 | 34.05 ± 0.32 | 34.26 ± 0.37 b | 35.87 ± 0.43 a | 34.78 ± 0.22 b | 32.38 ± 0.24 c |
| 22–42 | 77.55 ± 1.57 A | 73.23 ± 0.42 Bb | 77.11 ± 1.21 a | 77.90 ± 1.43 a | 71.79 ± 1.11 b | |
| 1–42 | 56.84 ± 0.74 | 54.67 ± 0.80 ab | 57.47 ± 0.56 a | 57.37 ± 0.66 a | 52.18 ± 1.28 b | |
| FCR, g:g | 1–21 | 1.63 ± 0.00 | 1.67 ± 0.06 | 1.66 ± 0.01 | 1.66 ± 0.01 | 1.64 ± 0.03 |
| 22–42 | 1.92 ± 0.00 | 1.93 ± 0.04 | 2.01 ± 0.03 | 1.95 ± 0.02 | 2.00 ± 0.06 | |
| 1–42 | 1.83 ± 0.00 | 1.84 ± 0.01 | 1.89 ± 0.02 | 1.85 ± 0.02 | 1.85 ± 0.02 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azzam, M.M.; JIANG, S.-q.; CHEN, J.-l.; LIN, X.-j.; GOU, Z.-y.; FAN, Q.-l.; WANG, Y.-b.; LI, L.; JIANG, Z.-y. Effect of Soybean Isoflavones on Growth Performance, Immune Function, and Viral Protein 5 mRNA Expression in Broiler Chickens Challenged with Infectious Bursal Disease Virus. Animals 2019, 9, 247. https://doi.org/10.3390/ani9050247
Azzam MM, JIANG S-q, CHEN J-l, LIN X-j, GOU Z-y, FAN Q-l, WANG Y-b, LI L, JIANG Z-y. Effect of Soybean Isoflavones on Growth Performance, Immune Function, and Viral Protein 5 mRNA Expression in Broiler Chickens Challenged with Infectious Bursal Disease Virus. Animals. 2019; 9(5):247. https://doi.org/10.3390/ani9050247
Chicago/Turabian StyleAzzam, Mahmoud Mostafa, Shou-qun JIANG, Jia-li CHEN, Xia-jing LIN, Zhong-yong GOU, Qiu-li FAN, Yi-bing WANG, Long LI, and Zong-yong JIANG. 2019. "Effect of Soybean Isoflavones on Growth Performance, Immune Function, and Viral Protein 5 mRNA Expression in Broiler Chickens Challenged with Infectious Bursal Disease Virus" Animals 9, no. 5: 247. https://doi.org/10.3390/ani9050247