Effects of Killing Methods on Lipid Oxidation, Colour and Microbial Load of Black Soldier Fly (Hermetia illucens) Larvae

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Killing Methods
2.2.1. Mechanical Disruption
2.2.2. Heating
2.2.3. Freezing
2.2.4. Asphyxiation
2.3. Analysis
2.3.1. Chemical Composition and pH
2.3.2. Primary Lipid Oxidation
2.3.3. Secondary Lipid Oxidation
2.3.4. Larval Colouration and Colour Change Occurring While Drying
2.3.5. Microbiological Analysis
2.4. Statistical Analysis
3. Results
3.1. Chemical Composition and pH
3.2. Lipid Oxidation
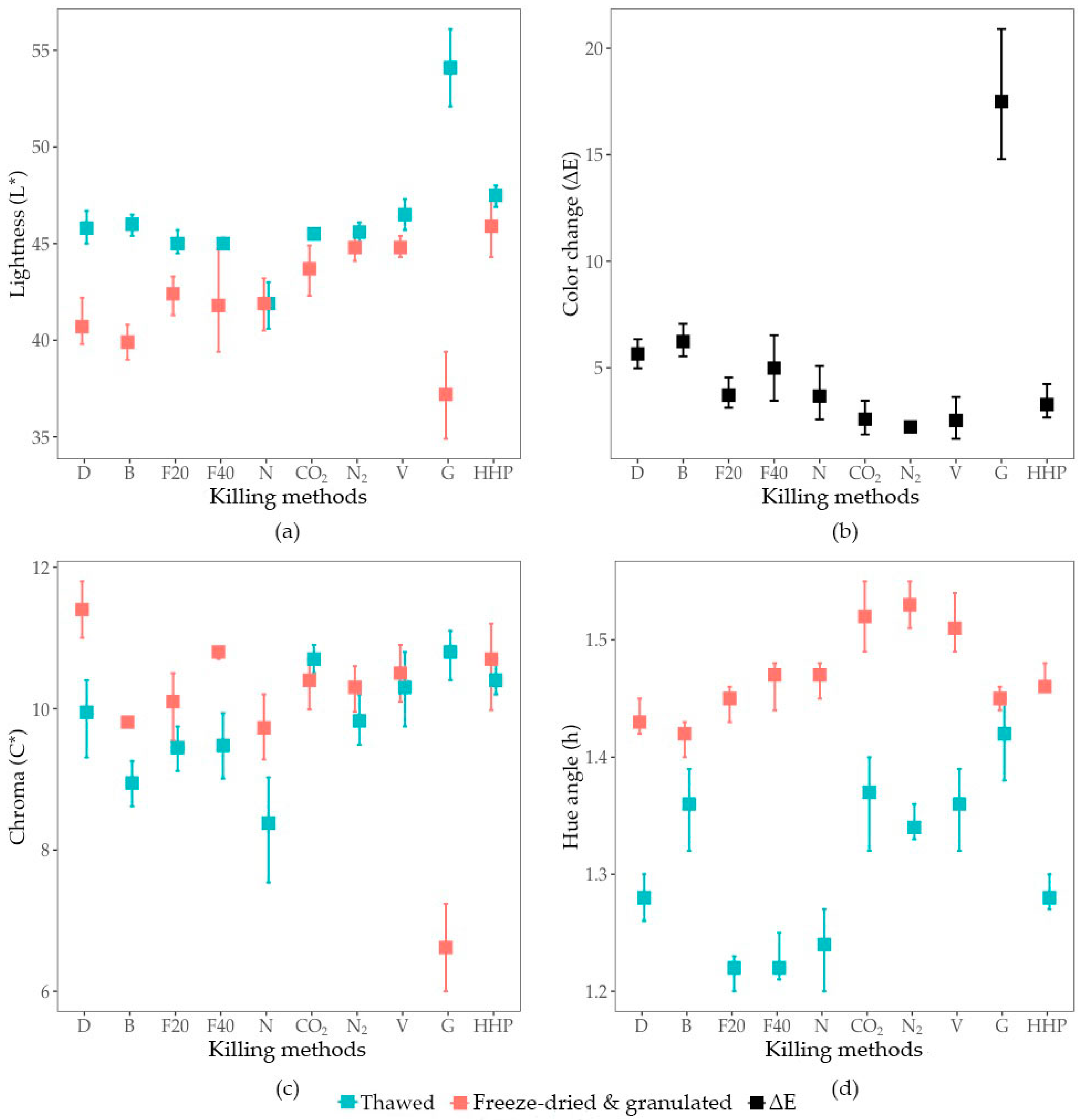
3.3. Larval Colour
3.3.1. Impact of Freeze-Drying on the Colour Change
3.3.2. Colour of the Freeze-Dried and Granulated Product
3.4. Microbial Analysis
4. Discussion
4.1. Chemical Composition
4.2. Colour
4.3. Microbiology
4.4. Heating
4.4.1. Desiccation
4.4.2. Blanching
4.5. Freezing
4.6. Asphyxiation
4.7. Mechanical Disruption
4.7.1. Grinding
4.7.2. High Hydrostatic Pressures
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Killing Methods | L* | a* | b* | Chroma (C*) | Hue Angle (h) | ΔE |
|---|---|---|---|---|---|---|
| Thawed | ||||||
| Desiccation | 45.8 de ± 1.0 | 2.9 gh ± 0.3 | 9.5 d ± 0.7 | 9.9 cd ± 0.7 | 1.28 ab ± 0.03 | 5.6 b ± 0.9 |
| Blanching | 46.0 de ± 0.7 | 1.9 def ± 0.4 | 8.7 bc ± 0.3 | 8.9 bc ± 0.4 | 1.36 bce ± 0.04 | 6.2 b ± 0.9 |
| −20 °C | 45.0 cde ± 0.7 | 3.3 h ± 0.2 | 8.9 bc ± 0.3 | 9.5 bcd ± 0.4 | 1.22 a ± 0.02 | 3.7 ab ± 0.9 |
| −40 °C | 45.0 cde ± 0.4 | 3.2 h ± 0.2 | 8.9 bc ± 0.6 | 9.5 bcd ± 0.6 | 1.22 a ± 0.02 | 5.0 ab ± 1.9 |
| Liquid N | 41.9 bc ± 1.5 | 2.7 fgh ± 0.4 | 7.9 ab ± 0.9 | 8.4 b ± 0.9 | 1.24 a ± 0.05 | 3.7 ab ± 1.4 |
| CO2 | 45.5 cde ± 0.2 | 2.2 efg ± 0.5 | 10.5 de ± 0.2 | 10.7 de ± 0.2 | 1.37 cdefg ± 0.05 | 2.6 a ± 0.9 |
| N2 | 45.6 cde ± 0.6 | 2.2 efg ± 0.2 | 9.6 cd ± 0.4 | 9.8 cd ± 0.4 | 1.34 bcd ± 0.02 | 2.2 a ± 0.2 |
| Vacuum | 46.5 e ± 0.9 | 2.2 efg ± 0.4 | 10.1 cde ± 0.7 | 10.3 cde ± 0.7 | 1.36 bcdef ± 0.04 | 2.5 a ± 1.1 |
| Grinding | 54.1 f ± 2.5 | 1.6 cde ± 0.4 | 10.6 de ± 0.4 | 10.8 de ± 0.4 | 1.42 cdefgh ± 0.04 | 17.5 c ± 3.6 |
| High presure | 47.5 e ± 0.7 | 3.0 gh ± 0.2 | 10.0 cde ± 0.3 | 10.4 de ± 0.3 | 1.28 ab ± 0.02 | 3.3 ab ± 1.0 |
| Freeze-dried and granulated | ||||||
| Desiccation | 40.7 ab ± 1.5 | 1.6 bcde ± 0.2 | 11.3 e ± 0.4 | 11.4 e ± 0.4 | 1.43 efgh ± 0.02 | |
| Blanching | 39.9 ab ± 1.1 | 1.5 bcde ± 0.2 | 9.7 cd ± 0.1 | 9.8 cd ± 0.1 | 1.42 dfgh ± 0.02 | |
| −20 °C | 42.4 bcd ± 1.2 | 1.3 abcd ± 0.3 | 10.1 cde ± 0.6 | 10.1 cde ± 0.6 | 1.45 ghi ± 0.02 | |
| −40 °C | 40.7 bc ± 1.6 | 1.1 abcd ± 0.3 | 10.5 de ± 0.3 | 10.6 de ± 0.3 | 1.47 hij ± 0.03 | |
| Liquid N | 41.9 bc ± 1.7 | 1.0 abc ± 0.2 | 9.7 cd ± 0.5 | 9.7 bcd ± 0.6 | 1.47 hij ± 0.02 | |
| CO2 | 44.8 bcde ± 1.1 | 0.6 a ± 0.4 | 10.5 de ± 0.5 | 10.5 de ± 0.5 | 1.52 ij ± 0.04 | |
| N2 | 44.8 cde ± 0.8 | 0.4 a ± 0.3 | 10.3 de ± 0.3 | 10.3 cde ± 0.3 | 1.53 j ± 0.02 | |
| Vacuum | 44.8 cde ± 0.7 | 0.6 a ± 0.3 | 10.5 de ± 0.5 | 10.5 de ± 0.5 | 1.51 ij ± 0.03 | |
| Grinding | 37.2 a ± 2.6 | 0.8 ab ± 0.1 | 6.6 a ± 0.8 | 6.6 a ± 0.8 | 1.45 hij ± 0.01 | |
| High pressure | 45.9 de ± 2.0 | 1.1 abcd ± 0.2 | 10.6 de ± 0.7 | 10.7 de ± 0.7 | 1.46 hij ± 0.01 |
References
- Oonincx, D.G.; van Broekhoven, S.; van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The Potential Role of Insects as Feed: A Multi-Perspective Review. Animals 2019, 9, 119. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquac. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.; Ur Rehman, K.; Li, W.; Cai, M.; Li, Q.; Mazza, L.; Zhang, J.; et al. Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 2017, 12, e0182601. [Google Scholar] [CrossRef] [PubMed]
- Wynants, E.; Frooninckx, L.; Crauwels, S.; Verreth, C.; De Smet, J.; Sandrock, C.; Wohlfahrt, J.; Van Schelt, J.; Depraetere, S.; Lievens, B.; et al. Assessing the microbiota of black soldier fly larvae (Hermetia illucens) reared on organic waste streams on four different locations at laboratory and large scale. Microb. Ecol. 2018, in press. [Google Scholar] [CrossRef]
- Kashiri, M.; Marin, C.; Garzon, R.; Rosell, C.M.; Rodrigo, D.; Martinez, A. Use of high hydrostatic pressure to inactivate natural contaminating microorganisms and inoculated E. coli O157:H7 on Hermetia illucens larvae. PLoS ONE 2018, 13, e0194477. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.; Tomberlin, J.K.; Diener, S.; Zurbrügg, C.; Mathys, A. Decomposition of biowaste macronutrients, microbes, and chemicals in black soldier fly larval treatment: A review. Waste Manag. 2018, 82, 302–318. [Google Scholar] [CrossRef]
- Contreras, S.P. Inactivación de Microorganismos Contaminantes Naturales y Salmonella typhimurium en Larvas de Moscas Soldado Negra (Hermetia illucens) Mediante Tratamientos de Alta Presión Hidrostática HHP. Master’s Thesis, Universitat Politècnica de València, València, Spain, 4 July 2017. [Google Scholar]
- Janssen, R.H.; Vincken, J.-P.; Arts, N.J.G.; Fogliano, V.; Lakemond, C.M.M. Effect of endogenous phenoloxidase on protein solubility and digestibility after processing of Tenebrio molitor, Alphitobius diaperinus and Hermetia illucens. Food Res. Int. 2019. [Google Scholar] [CrossRef]
- Van Huis, A.; Tomberlin, J.K. Insects as Food and Feed: From Production to Consumption; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; p. 447. [Google Scholar]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.A. Effect of CO2 on neurons of the house cricket, Acheta domestica. J. Neurobiol. 1982, 14, 237–250. [Google Scholar] [CrossRef]
- Fregin, T.; Bickmeyer, U. Electrophysiological investigation of different methods of anesthesia in lobster and crayfish. PLoS ONE 2016, 11, e0162894. [Google Scholar] [CrossRef]
- Mutungi, C.; Irungu, F.G.; Nduko, J.; Mutua, F.; Affognon, H.; Nakimbugwe, D.; Ekesi, S.; Fiaboe, K.K.M. Postharvest processes of edible insects in Africa: A review of processing methods, and the implications for nutrition, safety and new products development. Crit. Rev. Food Sci. Nutr. 2017, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Fröhling, A.; Reineke, K.; Knorr, D.; Boguslawski, S.; Ehlbeck, J.; Schlüter, O. Comparison of volumetric and surface decontamination techniques for innovative processing of mealworm larvae (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2014, 26, 232–241. [Google Scholar] [CrossRef]
- Vandeweyer, D.; Lenaerts, S.; Callens, A.; Van Campenhout, L. Effect of blanching followed by refrigerated storage or industrial microwave drying on the microbial load of yellow mealworm larvae (Tenebrio molitor). Food Control 2017, 71, 311–314. [Google Scholar] [CrossRef]
- Janssen, R.H.; Canelli, G.; Sanders, M.G.; Bakx, E.J.; Lakemond, C.M.M.; Fogliano, V.; Vincken, J.P. Iron-polyphenol complexes cause blackening upon grinding Hermetia illucens (black soldier fly) larvae. Sci. Rep. 2019, 9, 2967. [Google Scholar] [CrossRef] [PubMed]
- Leni, G.; Caligiani, A.; Sforza, S. Killing method affects the browning and the quality of the protein fraction of black soldier fly (Hermetia illucens) prepupae: A metabolomics and proteomic insight. Food Res. Int. 2019, 115, 116–125. [Google Scholar] [CrossRef]
- David-Birman, T.; Raften, G.; Lesmes, U. Effects of thermal treatments on the colloidal properties, antioxidant capacity and in-vitro proteolytic degradation of cricket flour. Food Hydrocolloids 2018, 79, 48–54. [Google Scholar] [CrossRef]
- Gonçalves, A.A.; de Oliveira, A.R.M. Melanosis in crustaceans: A review. LWT 2016, 65, 791–799. [Google Scholar] [CrossRef]
- Huang, W.; Ji, H.; Liu, S.; Zhang, C.; Chen, Y.; Guo, M.; Hao, J. Inactivation effects and kinetics of polyphenol oxidase from Litopenaeus vannamei by ultra-high pressure and heat. Innov. Food Sci. Emerg. Technol. 2014, 26, 108–115. [Google Scholar] [CrossRef]
- Zamorano, J.-P.; Martínez-Álvarez, O.; Montero, P.; Gómez-Guillén, M.d.C. Characterization and tissue distribution of polyphenol oxidase of deepwater pink shrimp (Parapenaeus longirostris). Food Chem. 2009, 112, 104–111. [Google Scholar] [CrossRef]
- Miyashita, K.; Kamal-Eldin, A.; Min, D.B. Lipid Oxidation Pathways; AOCS: Urbana, IL, USA, 2010; Volume 2, p. 324. [Google Scholar]
- Lenaerts, S.; Van Der Borght, M.; Callens, A.; Van Campenhout, L. Suitability of microwave drying for mealworms (Tenebrio molitor) as alternative to freeze drying: Impact on nutritional quality and colour. Food Chem. 2018, 254, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Medina-Meza, I.G.; Barnaba, C.; Barbosa-Cánovas, G.V. Effects of high pressure processing on lipid oxidation: A review. Innov. Food Sci. Emerg. Technol. 2014, 22, 1–10. [Google Scholar] [CrossRef]
- Lalander, C.H.; Fidjeland, J.; Diener, S.; Eriksson, S.; Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 2014, 35, 261–271. [Google Scholar] [CrossRef]
- Canadian Food Inspection Agency (CFIA). Bacterial Pathogens in Edible Insects; CFIA: Ottawa, ON, Canada, 2018; p. 9.
- Caligiani, A.; Marseglia, A.; Sorci, A.; Bonzanini, F.; Lolli, V.; Maistrello, L.; Sforza, S. Influence of the killing method of the black soldier fly on its lipid composition. Food Res. Int. 2019, 116, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Adámková, A.; Adámek, M.; Mlček, J.; Borkovcová, M.; Bednářová, M.; Kouřimská, L.; Skácel, J.; Vítová, E. Welfare of the mealworm (Tenebrio molitor) breeding with regard to nutrition value and food safety. Potravinarstvo Slovak J. Food Sci. 2017, 11, 460–465. [Google Scholar] [CrossRef]
- Farina, M.F. How method of killing crickets impact the sensory qualities and physiochemical properties when prepared in a broth. Int. J. Gastron. Food Sci. 2017, 8, 19–23. [Google Scholar] [CrossRef]
- Hogsette, J.A. New diets for production of house-flies and stable flies (Diptera, Muscidae) in the laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef] [PubMed]
- Paz, A.S.P.; Carrejo, N.S.; Rodriguez, C.H.G. Effects of larval density and feeding rates on the bioconversion of vegetable waste using black soldier fly larvae Hermetia illucens (L.), (Diptera: Stratiomyidae). Waste Biomass Valoriz. 2015, 6, 1059–1065. [Google Scholar]
- Armstrong, J.W.; Tang, J.; Wang, S. Thermal death kinetics of mediterranean, malaysian, melon, and oriental fruit fly (Diptera: Tephritidae) eggs and third instars. J. Econ. Entomol. 2009, 102, 522–532. [Google Scholar] [CrossRef]
- Hashem, M.Y.; Ahmed, S.S.; El-Mohandes, M.A.; Hussain, A.R.E.; Ghazy, S.M. Comparative effectiveness of different modified atmospheres enriched with carbon dioxide and nitrogen on larval instars of almond moth Ephestia cautella (Walker) (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2014, 59, 314–319. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis, 20th ed.; AOCS: Rockville, MD, USA, 2016; p. 3172. [Google Scholar]
- AOCS (American Oil Chemists’ Society). Official Methods and Recommended Practices of the American Oil Chemists’ Society, 5th ed.; AOCS Press: Urbana, IL, USA, 1998; p. 1200. [Google Scholar]
- Koniecko, E.S. Handbook of Meat Analysis, 2nd ed.; Avery Pub Group: New York City, NY, USA, 1984; p. 289. [Google Scholar]
- Hermes-Lima, M.; Willmore, W.G.; Storey, K.B. Quantification of lipid peroxidation in tissue extracts based on Fe(III)xylenol orange complex formation. Free Radic. Biol. Med. 1995, 19, 271–280. [Google Scholar] [CrossRef]
- Grau, A.; Codony, R.; Rafecas, M.; Barroeta, A.C.; Guardiola, F. Lipide hydroperoxide determination in dark chicken meat through a ferrous oxidation—Xylenol orange method. J. Agric. Food Chem. 2000, 48, 4136–4143. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Chem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Joanisse, D.R.; Storey, K.B. Oxidative stress and antioxidants in stress and recovery of cold-hardy insects. Insect. Biochem. Mol. Biol. 1998, 28, 23–30. [Google Scholar] [CrossRef]
- International Commission on Illumination (CIE). Colorimetry, 3rd ed.; CIE Central Bureau: Vienna, Austria, 2004; p. 111. [Google Scholar]
- Purschke, B.; Brüggen, H.; Scheibelberger, R.; Jäger, H. Effect of pre-treatment and drying method on physico-chemical properties and dry fractionation behaviour of mealworm larvae (Tenebrio molitor L.). Eur. Food Res. Technol. 2018, 244, 269–280. [Google Scholar] [CrossRef]
- Hosseinpour, S.; Rafiee, S.; Mohtasebi, S.S.; Aghbashlo, M. Application of computer vision technique for on-line monitoring of shrimp color changes during drying. J. Food Eng. 2013, 115, 99–114. [Google Scholar] [CrossRef]
- Klunder, H.C.; Wolkers-Rooijackers, J.; Korpela, J.M.; Nout, M.J.R. Microbiological aspects of processing and storage of edible insects. Food Control 2012, 26, 628–631. [Google Scholar] [CrossRef]
- Health Canada: The Compendium of Analytical Methods—Vol.2. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/research-programs-analytical-methods/analytical-methods/compendium-methods/methods-microbiological-analysis-foods-compendium-analytical-methods.html (accessed on 30 January 2018).
- Kone, A.P.; Desjardins, Y.; Gosselin, A.; Cinq-Mars, D.; Guay, F.; Saucier, L. Plant extracts and essential oil product as feed additives to control rabbit meat microbial quality. Meat Sci. 2019, 150, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Lebeuf, Y.; Vandenberg, G.W. Microbial Load Analysis of Frozen Black Soldier Fly (Hermetia illucens) Larvae from the Rearing Facility in the Laboratoire des Sciences Aquatiques of Université Laval; Département des Sciences Animales, Université Laval: Québec, QC, Canada, 2019. [Google Scholar]
- Driemeyer, H. Evaluation of Black Soldier Fly (Hermetia illucens) Larvae as an Alternative Protein Source in Pig Creep Diets in Relation to Production, Blood and Manure Microbiology Parameters. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, December 2016. [Google Scholar]
- Seenger, J.; Nuernberg, G.; Hartung, M.; Szúcs, E.; Ender, K.; Nuernberg, K. ANKOM—A new instrument for the determination of fat in muscle and meat cuts—A comparison. Arch. Tierz. 2008, 51, 449–457. [Google Scholar] [CrossRef]
- Lewicki, P.P. Water as the determinant of food engineering properties. A review. J. Food Eng. 2004, 61, 483–495. [Google Scholar] [CrossRef]
- Erdogdu, F.; Balaban, M.O.; Otwell, W.S.; Garrido, L. Cook-related yield loss for pacific white (Penaeus vannamei) shrimp previously treated with phosphates: Effects of shrimp size and internal temperature distribution. J. Food Eng. 2004, 64, 297–300. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Minister of Justice. Food and Drug Regulations—Part B: Foods; Government of Canada, Minister of Justice: Ottawa, ON, Canada, 2018; Volume B.27.
- Halliwall, B.; Chirico, S. Lipid peroxidation: Its mechanism, measurement, and significance. Am. J. Clin. Nutr. 1993, 57, 715–725. [Google Scholar] [CrossRef]
- Barden, L.; Decker, E.A. Lipid oxidation in low-moisture food: A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 2467–2482. [Google Scholar] [CrossRef] [PubMed]
- Robles-Martinez, C.; Cervantes, E.; Ke, P.J. Recommended method for testing the objective rancidity development in fish based on TBARS formation. Can. Tech. Rep. Fish. Aquat. Sci. 1982, 1089, 1–27. [Google Scholar]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbo, R.; Torstensen, B.E.; Lock, E.J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef] [PubMed]
- Sosa, D.A.T.; Fogliano, V. Potential of insect-derived ingredients for food applications. In Insect Physiology and Ecology; IntechOpen: London, UK, 2017; p. 19. [Google Scholar]
- Montero, P.; Avalos, A.; Pérez-Mateos, M. Characterization of polyphenoloxidase of prawns (Penaeus japonicus). Alternatives to inhibition: Additives and high-pressure treatment. Food Chem. 2001, 75, 317–324. [Google Scholar] [CrossRef]
- Melis, R.; Braca, A.; Mulas, G.; Sanna, R.; Spada, S.; Serra, G.; Fadda, M.L.; Roggio, T.; Uzzau, S.; Anedda, R. Effect of freezing and drying processes on the molecular traits of edible yellow mealworm. Innov. Food Sci. Emerg. Technol. 2018, 48, 138–149. [Google Scholar] [CrossRef]
- Francis, F.J.; Clydesdate, F.M. Food Colorimetry: Theory and Applications; The AVI Publishing Co., Inc.: Wesport, CT, USA, 1975; p. 477. [Google Scholar]
- Larouche, J.; Vandenberg, G.W. Microbial Load Analysis of the Gainesville Diet Use as Feed for Black Soldier Fly (Hermetia illucens) Larvae in the Laboratoire des Sciences Aquatiques of Université Laval; Département des Sciences Animales, Université Laval: Québec, QC, Canada, 2019. [Google Scholar]
- Health Canada. Microbial Guidelines for Ready-to-Eat Foods—A Guide for the Conveyances Industry and Environmental Health Officers (EHO); Health Canada: Ottawa, ON, Canada, 2013; p. 9.
- Russell, N.J.; Gould, G.W. Food Preservatives, 2nd ed.; Springer: Berlin, Germany, 2003; p. 380. [Google Scholar]
- Ross, T.; Dalgaard, P.; Tienungoon, S. Predictive modelling of the growth and survival of Listeria in fishery products. Int. J. Food Microbiol. 2000, 62, 231–245. [Google Scholar] [CrossRef]
- Moats, W.A. Kinetics of thermal death of bacteria. J. Bacteriol. 1971, 105, 165–171. [Google Scholar]
- Connell, J.J. Control of Fish Quality; Fishing News: West Byfleet, UK, 1975; p. 179. [Google Scholar]
- Azzollini, D.; Derossi, A.; Severini, C. Understanding the drying kinetic and hygroscopic behaviour of larvae of yellow mealworm (Tenebrio molitor) and the effects on their quality. J. Insects Food Feed 2016, 2, 233–243. [Google Scholar] [CrossRef]
- Reis, F.R. New Perspectives on Food Blanching; Springer Nature: Basingstoke, UK, 2017; p. 162. [Google Scholar]
- Bhat, T.H.; Chouksey, M.K.; Balange, A.K.; Nayak, B.B. Effect of heat treatment at different stages of rigor on the quality of pacific white shrimp (Litopenaeus vannamei). J. Aquat. Food Prod. Technol. 2017, 26, 770–780. [Google Scholar] [CrossRef]
- Tranter, H.S.; Board, R.G. The influence of incubation temperature and pH on the antimicrobial properties of hen egg albumen. J. Appl. Microbiol. 1984, 56, 53–61. [Google Scholar] [CrossRef]
- Jay, J.M. Modern Food Microbiology, 4th ed.; Chapman & Hall: New York, NY, USA, 1992; p. 701. [Google Scholar]
- Yu, L.; Jiang, Q.; Yu, D.; Xu, Y.; Gao, P.; Xia, W. Quality of giant freshwater prawn (Macrobrachium rosenbergii) during the storage at −18°C as affected by different methods of freezing. Int. J. Food Prop. 2018, 21, 2100–2109. [Google Scholar] [CrossRef]
- Nilson, T.L.; Sinclair, B.J.; Roberts, S.P. The effects of carbon dioxide anesthesia and anoxia on rapid cold-hardening and chill coma recovery in Drosophila melanogaster. J. Insect Physiol. 2006, 52, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Wong-Corral, F.J.; Castañé, C.; Riudavets, J. Lethal effects of CO2-modified atmospheres for the control of three Bruchidae species. J. Stored Prod. Res. 2013, 55, 62–67. [Google Scholar] [CrossRef]
- Veen, M.; Lang, C. Production of lipid compounds in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2004, 63, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Marrakchi, H.; Choi, K.H.; Rock, C.O. A new mechanism for anaerobic unsaturated fatty acid formation in Streptococcus pneumoniae. J. Biol. Chem. 2002, 277, 44809–44816. [Google Scholar] [CrossRef]
- Woodring, J.P.; Clifford, C.W.; Roe, R.M.; Beckman, B.R. Effects of CO2 and anoxia on feeding, growth, metabolism, water balance, and blood composition in larval female house crickets, Acheta domesticus. J. Insect Physiol. 1978, 24, 499–509. [Google Scholar] [CrossRef]
- Sun, B.; Zhao, Y.; Yu, J.; Ling, J.; Shang, H.; Liu, Z. The combined efficacy of superchilling and high CO2 modified atmosphere packaging on shelf life and quality of swimming crab (Portunus trituberculatus). J. Aquat. Food Prod. Technol. 2017, 26, 655–664. [Google Scholar] [CrossRef]
- Farber, J.M. Microbiological aspects of modified-atmosphere packaging technology—A review. J. Food Prot. 1991, 54, 58–70. [Google Scholar] [CrossRef]
- Shimoda, M.; Cocunubo-Castellanos, J.; Kago, H.; Miyake, M.; Osajima, Y.; Hayakawa, I. The influence of dissolved CO2 concentration on the death kinetics of Saccharomyces cerevisiae. J. Appl. Microbiol. 2001, 91, 306–311. [Google Scholar] [CrossRef]
- Balasubramaniam, V.M.; Martinez-Monteagudo, S.I.; Gupta, R. Principles and application of high pressure-based technologies in the food industry. Annu. Rev. Food Sci. Technol. 2015, 6, 435–462. [Google Scholar] [CrossRef] [PubMed]
- Margosch, D.; Ehrmann, M.A.; Buckow, R.; Heinz, V.; Vogel, R.F.; Ganzle, M.G. High-pressure-mediated survival of Clostridium botulinum and Bacillus amyloliquefaciens endospores at high temperature. Appl. Environ. Microbiol. 2006, 72, 3476–3481. [Google Scholar] [CrossRef] [PubMed]
- Cheftel, J.C. Review: High-pressure, microbial inactivation and food preservation. Food Sci. Technol. Int. 1995, 1, 75–90. [Google Scholar] [CrossRef]
- Barba, F.J.; Terefe, N.S.; Buckow, R.; Knorr, D.; Orlien, V. New opportunities and perspectives of high pressure treatment to improve health and safety attributes of foods. A review. Food Res. Int. 2015, 77, 725–742. [Google Scholar] [CrossRef]


| Killing Methods | Total Moisture Content (%; Wet Basis) | Ash (%; Dry Basis) | Ether Extract (%; Dry Basis) | FOX (mg CHP equivalents/kg; Wet Basis) | TBARS (mg MDA/kg; Wet Basis) | pH |
|---|---|---|---|---|---|---|
| Heating | ||||||
| D | 78.1 a ± 1.0 | 7.9 ab ± 0.6 | 13.4 ab ± 1.6 | 5.8 a ± 2.6 | 2.5 c ± 0.2 | 7.8 c ± 0.5 |
| B | 78.6 a ± 0.7 | 8.5 b ± 0.3 | 14.5 bc ± 0.5 | 6.4 a ± 2.3 | 1.0 a ± 0.1 | 8.7 d ± 0.1 |
| Freezing | ||||||
| F20 | 81.6 b ± 0.3 | 7.1 a ± 0.4 | 12.8 ab ± 0.5 | 7.2 a ± 0.9 | 1.7 b ± 0.2 | 7.4 bc ± 0.3 |
| F40 | 81.3 b ± 0.4 | 7.0 a ± 0.5 | 12.4 a ± 0.7 | 7.4 a ± 1.0 | 2.0 bc ± 0.3 | 7.4 bc ± 0.3 |
| N | 80.8 b ± 1.5 | 7.1 a ± 0.3 | 12.6 a ± 1.2 | 7.9 a ± 2.2 | 1.8 b ± 0.3 | 7.3 bc ± 0.3 |
| Asphyxiation | ||||||
| CO2 | 83.5 c ± 0.3 | 7.1 a ± 0.4 | 15.9 cd ± 1.6 | 19.4 b ± 4.8 | 2.0 b ± 0.2 | 6.1 a ± 0.3 |
| N2 | 83.2 c ± 1.3 | 7.3 a ± 0.7 | 16.6 d ± 1.6 | 18.6 b ± 7.1 | 1.6 b ± 0.1 | 6.3 a ± 0.1 |
| V | 83.6 c ± 0.5 | 7.3 a ± 0.4 | 15.9 cd ± 1.9 | 18.7 b ± 7.2 | 1.9 b ± 0.3 | 6.2 a ± 0.4 |
| Mechanical disruption | ||||||
| G | 80.3 b ± 0.2 | 7.9 ab ± 0.3 | 11.9 a ± 0.3 | 6.7 a ± 1.2 | 2.0 b ± 0.3 | 7.5 bc ± 0.2 |
| HHP | 80.8 b ± 1.0 | 7.0 a ± 0.6 | 12.0 a ± 0.7 | 7.3 a ± 3.4 | 1.6 b ± 0.3 | 7.2 b ± 0.1 |
| Killing Methods | Total Viable Aerobic Count | Lactic Acid Bacteria | Pseudomonas spp. | Yeast and Moulds | Enterobacteria | Coliforms | Listeria spp. | Clostridia and Other Anaerobic |
|---|---|---|---|---|---|---|---|---|
| Heating | ||||||||
| D | 7.7 b ± 0.7 | 7.8 bc ± 0.7 | 4.8 b ± 0.4 | 5.1 bc ± 0.5 | 4.1 b ± 0.8 | 3.5 b ± 1.3 | 5.2 a ± 0.2 | 7.9 c ± 0.5 |
| B | 5.6 a ± 0.3 | 3.1 a ± 0.8 | <2.1 a ± 0.1 | <2.1 a ± 0.1 | <1.1 a ± 0.1 | <1.1 a ± 0.1 | 5.2 a ± 0.1 | 4.8 a ± 0.3 |
| Freezing | ||||||||
| F20 | 8.3 bc ± 0.5 | 8.4 cd ± 0.6 | 5.6 b ± 0.3 | 6.3 c ± 0.8 | 4.5 b ± 0.9 | 4.5 b ± 0.9 | 5.5 ab ± 0.2 | 8.6 cd ± 0.2 |
| F40 | 8.5 c ± 0.1 | 8.6 cde ± 0.1 | 6.7 c ± 0.7 | 6.4 c ± 0.7 | 5.3 b ± 0.9 | 5.1 bc ± 0.8 | 5.1 a ± 0.1 | 8.4 cd ± 0.1 |
| N | 8.3 bc ± 0.2 | 8.7 cde ± 0.1 | 7.0 cd ± 0.4 | 6.1 c ± 0.7 | 4.6 b ± 0.7 | 4.5 b ± 0.7 | 5.2 a ± 0.1 | 8.4 cd ± 0.3 |
| Asphyxiation | ||||||||
| CO2 | 9.8 d ± 0.2 | 9.9 f ± 0.2 | 7.8 d ± 0.4 | 5.8 bc ± 0.6 | 7. 5 c ± 0.6 | 7.6 d ± 0.6 | 7.0 c ± 0.6 | 9.9 f ± 0.1 |
| N2 | 9.6 d ± 0.2 | 9.7 ef ± 0.1 | 7.3 cd ± 0.3 | 5.3 bc ± 0.9 | 7. 3 c ± 0.6 | 7.2 d ± 0.4 | 6.7 c ± 0.7 | 9.6 ef ± 0.1 |
| V | 9.6 d ± 0.2 | 9.7 ef ± 0.3 | 7.3 cd ± 0.2 | 4.2 b ± 1.2 | 6.9 c ± 0.7 | 6.7 cd ± 0.5 | 6.3 bc ± 0.8 | 9.6 ef ± 0.2 |
| Mechanical disruption | ||||||||
| G | 8.5 c ± 0.5 | 9.1 def ± 0.3 | 6.5 c ± 0.3 | 6.1 c ± 0.4 | 4.7 b ± 0.7 | 4.6 b ± 0.6 | 5.2 a ± 0.3 | 8.9 de ± 0.3 |
| HHP | 6.3 a ± 0.5 | 6.7 b ± 0.9 | <2.1 a ± 0.1 | <2.1 a ± 0.1 | <1.1 a ± 0.1 | 1.3 a ± 0.4 | 5.5 ab ± 0.5 | 6.1 b ± 0.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larouche, J.; Deschamps, M.-H.; Saucier, L.; Lebeuf, Y.; Doyen, A.; Vandenberg, G.W. Effects of Killing Methods on Lipid Oxidation, Colour and Microbial Load of Black Soldier Fly (Hermetia illucens) Larvae. Animals 2019, 9, 182. https://doi.org/10.3390/ani9040182
Larouche J, Deschamps M-H, Saucier L, Lebeuf Y, Doyen A, Vandenberg GW. Effects of Killing Methods on Lipid Oxidation, Colour and Microbial Load of Black Soldier Fly (Hermetia illucens) Larvae. Animals. 2019; 9(4):182. https://doi.org/10.3390/ani9040182
Chicago/Turabian StyleLarouche, Jennifer, Marie-Hélène Deschamps, Linda Saucier, Yolaine Lebeuf, Alain Doyen, and Grant W. Vandenberg. 2019. "Effects of Killing Methods on Lipid Oxidation, Colour and Microbial Load of Black Soldier Fly (Hermetia illucens) Larvae" Animals 9, no. 4: 182. https://doi.org/10.3390/ani9040182