Do the Calls of a Bird, the Noisy Miner (Manorina melanocephala), Need Adjustment for Efficient Communication in Urban Anthropogenic Noise?


{kind=link}
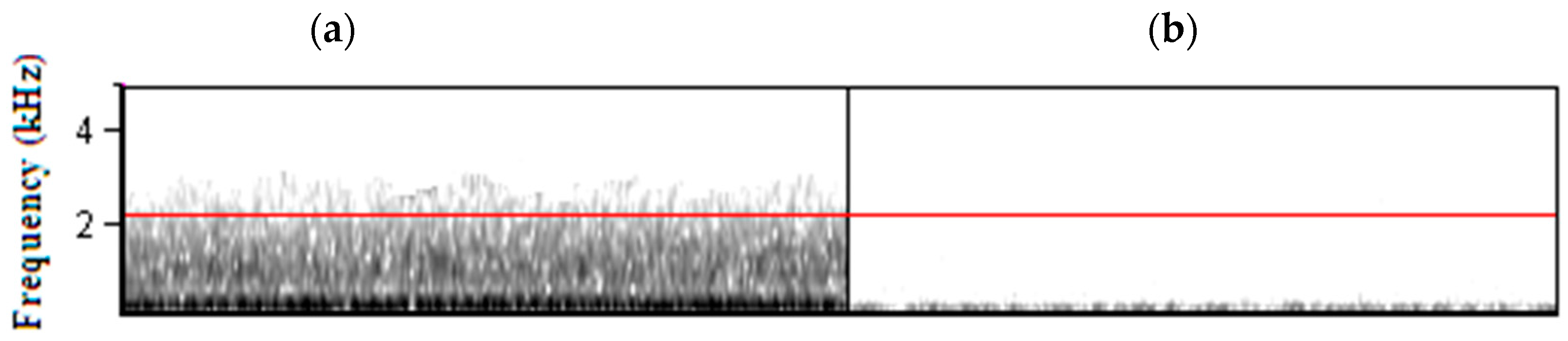{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Call Sampling
2.2. Ambient Noise Sampling
2.3. Analysis of Call Recordings
2.4. Analysis of Ambient Noise Recordings
2.5. Statistical Analyses
2.6. Ethical Standards
3. Results
3.1. Call Features
3.2. Ambient Noise
4. Discussion
4.1. Comparison of Call Features in Urban and Rural Habitats
4.1.1. Minimum Frequency
4.1.2. Dominant Frequency
4.1.3. Call Duration
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, Y.; Cardoso, G.C. Are bird species that vocalize at higher frequencies preadapted to inhabit noisy urban areas? Behav. Ecol. 2009, 20, 1268–1273. [Google Scholar] [CrossRef] [Green Version]
- Blair, R. The effects of urban sprawl on birds at multiple levels of biological organization. Ecol. Soc. 2004, 9, 2. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of species homogenisation. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Parsons, H.; Major, R.E.; French, K. Species interactions and habitat associations of birds inhabiting urban areas of Sydney, Australia. Austral Ecol. 2006, 31, 217–227. [Google Scholar] [CrossRef]
- Marsicovetere, V.; Lill, A. Doing what comes naturally: Are Australian Wood Ducks Chenonetta jubata pre-adapted for feeding in cities? Corella 2018, 42, 55–61. [Google Scholar]
- Lowry, H.; Lill, A.; Wong, B.B.M. Behavioural responses of wildlife to urban environments. Biol. Rev. 2013, 88, 537–549. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; Peet, M. Ecology: Birds sing at a higher pitch in urban noise. Nature 2003, 424, 267. [Google Scholar] [CrossRef]
- Lengagne, T. Traffic noise affects communication behaviour in a breeding anuran, Hyla arborea. Biol. Conserv. 2008, 141, 2023–2031. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; Ripmeester, E.A.P. Birdsong and anthropogenic noise: Implications and applications for conservation. Mol. Ecol. 2007, 17, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Slabbekoorn, H. Singing in the wild: The ecology of birdsong. In Nature’s Music: The Science of Birdsong; Marler, P., Slabbekoorn, H., Eds.; Elsevier Academic: Amsterdam, The Netherlands, 2004; pp. 178–205. [Google Scholar]
- Fuller, R.A.; Warren, P.H.; Gaston, K.J. Daytime noise predicts nocturnal singing in urban robins. Biol. Lett. 2007, 3, 368–370. [Google Scholar] [CrossRef] [Green Version]
- Brumm, H. The impact of environmental noise on song amplitude in a territorial bird. J. Anim. Ecol. 2004, 73, 434–440. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.E.; Yezerinac, S.M. Song Sparrow (Melospiza melodia) song varies with urban noise. Auk 2006, 123, 650–659. [Google Scholar] [CrossRef]
- Slabbekoorn, H.; den Boer-Visser, A. Cities change the songs of birds. Curr. Biol. 2006, 16, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Brumm, H. Blackbirds sing higher-pitched songs in cities: Adaptation to habitat acoustics or side-effect of urbanization? Anim. Behav. 2009, 28, 637–641. [Google Scholar] [CrossRef]
- Parris, K.M.; Schneider, A. Impacts of traffic volume on birds of roadside habitats. Ecol. Soc. 2009, 14, 29. [Google Scholar] [CrossRef]
- Potvin, D.A.; Parris, K.M.; Mulder, R.A. Geographically pervasive effects of urban noise on frequency and syllable rate of songs and calls in silvereyes (Zosterops lateralis). Proc. R. Soc. B 2011, 278, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Higgins, P.J.; Peter, J.M.; Steele, W.K. (Eds.) Noisy miner. In Handbook of Australian, New Zealand and Antarctic Birds, Volume 5. Tyrant-flycatchers to Chats; Oxford University Press: Melbourne, Australia, 2001; pp. 626–650. [Google Scholar]
- Low, T. The New Nature; Penguin Books: Camberwell, Australia, 2002; p. 416. [Google Scholar]
- Davitt, G.; Maute, K.; Major, R.E.; McDonald, P.G.; Maron, M. Short-term response of a declining woodland bird assemblage to the removal of a despotic competitor. Ecol. Evol. 2018, 8, 4771–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurisevic, M.A.; Sanderson, K.J. The vocal repertoires of six honeyeater (Meliphagidae) species from Adelaide, South Australia. Emu 1994, 94, 141–148. [Google Scholar] [CrossRef]
- Cunningham, S.; Magrath, R.D. Functionally referential alarm calls in noisy miners communicate about predator behavior. Anim. Behav. 2017, 129, 171–179. [Google Scholar] [CrossRef]
- Farrow, L.F.; Doohan, S.J.; McDonald, P.G. Alarm calls of a cooperative bird are referential and elicit context-specific antipredator behavior. Behav. Ecol. 2017, 28, 724–731. [Google Scholar] [CrossRef]
- Holt, J.; Barati, A.; McDonald, P.G. The complex acoustic repertoire of a highly social species, the Noisy Miner, Manorina melanocephala. Emu-Austral Ornithol. 2017, 117, 19–30. [Google Scholar]
- Clarke, M.F.; Oldland, J.M. Penetration of remnant edges by Noisy miners (Manorina melanocephala) and implications for habitat restoration. Wildl. Res. 2007, 34, 253–261. [Google Scholar] [CrossRef]
- Charif, R.A.; Clarke, C.W.; Fristrup, K.M. Ravenpro 1.3 User’s Manual 2007; Cornell University Laboratory of Ornithology: Ithaca, NY, USA, 2007. [Google Scholar]
- Hochachka, W.; Winter, M.; Charif, R.A. Sources of variation in singing probability of Florida grasshopper sparrows, and implications for design and analysis of auditory surveys. Condor 2009, 111, 349–360. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Poston, R.; Collibus, K.D.; Morgan, K.D.; Bastain, B.; Martin, C.; Jones, K.; Tremininio, R. Microhabitat selection and singing behavior patterns of male house finches (Carpodacus mexicanus) in urban parks in a heavily urbanized landscape in the Western U.S. Urban Habitats 2005, 3, 49–69. [Google Scholar]
- Slabbekoorn, H.; Yeh, P.; Hunt, K. Sound transmission and song divergence: A comparison of urban and forest acoustics. Condor 2007, 109, 67–78. [Google Scholar] [CrossRef]
- Hu, Y.; Cardoso, G.C. Which birds adjust the frequency of vocalizations in urban noise? Anim. Behav. 2010, 79, 863–867. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Candolin, U. Behavioral responses to changing environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Potvin, D.A.; Mulder, R.A.; Parris, K.M. Silvereyes decrease acoustic frequency but increase efficacy of alarm calls in urban noise. Anim. Behav. 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Parris, K.M.; Velik-Lord, M.; North, J.M.A. Frogs call at a higher pitch in traffic noise. Ecol. Soc. 2009, 14, 25. [Google Scholar] [CrossRef]
- Nemeth, E.; Brumm, H. Birds and anthropogenic noise: Are urban songs adaptive? Am. Nat. 2010, 176, 465–475. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A.; Wong, B.B.M. How noisy does a Noisy miner have to be? Amplitude adjustments of alarm calls in an avian urban ‘adapter’. PLoS ONE 2012, 7, e29960. [Google Scholar] [CrossRef]
- Brumm, H.; Zollinger, S.A.; Niemela, P.T.; Sprau, P. Measurement artefacts lead to false positives in the study of birdsong in noise. Methods Ecol. Evol. 2017, 8, 1617–1625. [Google Scholar] [CrossRef]
- Roca, I.T.; Desrochers, L.; Giacomazzo, M.; Bertolo, A.; Bolduc, P.; Deschesnes, R.; Martin, C.A.; Rainville, V.; Rheault, G.; Proulx, R. Shifting song frequencies in response to anthropogenic noise: A meta-analysis on birds and anurans. Behav. Ecol. 2016, 27, 1269–1274. [Google Scholar] [CrossRef]
- Rheindt, F.E. The impact of roads on birds: Does song frequency play a role in determining susceptibility to noise pollution? J. Ornithol. 2003, 144, 295–306. [Google Scholar] [CrossRef]
- Luther, D.A.; Phillips, J.; Derryberry, E.P. Not so sexy in the city: Urban birds adjust songs to noise but compromise vocal performance. Behav. Ecol. 2016, 27, 332–340. [Google Scholar] [CrossRef]
- Hunter, M.L.; Krebs, J.R. Geographical variation in the song of the Great tit (Parus major) in relation to ecological factors. J. Anim. Ecol. 1979, 48, 759–785. [Google Scholar] [CrossRef]
- Shy, E. The relation of geographical variation in song to habitat characteristics and body size in Northern American Tanagers (Thraupinae: Piranga). Behav. Ecol. Sociobiol. 1983, 12, 71–76. [Google Scholar] [CrossRef]
- Wiley, R.H. Associations of song properties with habitats for territorial oscine birds of Eastern North America. Am. Nat. 1991, 138, 973–993. [Google Scholar] [CrossRef]
- Nicholls, J.A.; Goldizen, A.W. Habitat type and density influence vocal signal design in satin bowerbirds. J. Anim. Ecol. 2006, 75, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayly, K.L.; Blumstein, D.T. Pied Currawongs and the decline of native birds. Emu 2001, 101, 199–204. [Google Scholar] [CrossRef]
- Rollinson, D.J. Synanthropy in the Australian Magpie: A Comparison of Urban and Rural Populations in Southeastern Queensland, Australia. Ph.D. Thesis, Griffith University, Brisbane, Australia, 2003. [Google Scholar]
- Shukuroglou, P.; McCarthy, M.A. Modelling the occurrence of rainbow lorikeets (Trichoglossus haematodus) in Melbourne. Austral Ecol. 2006, 31, 240–253. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A. Factors facilitating city-dwelling in red-rumped parrots. Wildl. Res. 2007, 34, 624–631. [Google Scholar] [CrossRef]
- Patricelli, G.L.; Blickley, J.L. Avian communication in Urban noise: Causes and consequences of vocal adjustment. Auk 2006, 123, 639–649. [Google Scholar] [CrossRef]
- Walsh, S.; Goulet, C.T.; Wong, B.B.M.; Chapple, D.G. Inherent behavioural traits enable a widespread lizard to cope with urban life. J. Zool. 2018, 306, 189–196. [Google Scholar] [CrossRef]
- Kirshel, A.N.G.; Blumstein, D.T.; Cohen, R.E.; Buermann, W.; Smith, T.B.; Slabbekoorn, H. Birdsong tuned to the environment: Green hylia song varies with elevation, tree cover, and noise. Behav. Ecol. 2009, 20, 1089–1095. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lowry, H.; Lill, A.; Wong, B.B.M. Do the Calls of a Bird, the Noisy Miner (Manorina melanocephala), Need Adjustment for Efficient Communication in Urban Anthropogenic Noise? Animals 2019, 9, 118. https://doi.org/10.3390/ani9030118
Lowry H, Lill A, Wong BBM. Do the Calls of a Bird, the Noisy Miner (Manorina melanocephala), Need Adjustment for Efficient Communication in Urban Anthropogenic Noise? Animals. 2019; 9(3):118. https://doi.org/10.3390/ani9030118
Chicago/Turabian StyleLowry, Hélène, Alan Lill, and Bob B. M. Wong. 2019. "Do the Calls of a Bird, the Noisy Miner (Manorina melanocephala), Need Adjustment for Efficient Communication in Urban Anthropogenic Noise?" Animals 9, no. 3: 118. https://doi.org/10.3390/ani9030118