Tissue Expression and Variation of the DGAT2 Gene and Its Effect on Carcass and Meat Quality Traits in Yak

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Carcass and Meat Quality Measurement
2.3. PCR-SSCP Analysis
2.4. Variant Sequencing and Analysis
2.5. Haplotype Determination
2.6. Statistical Analyses
2.7. RNA Extraction and RT-qPCR Analysis
3. Results
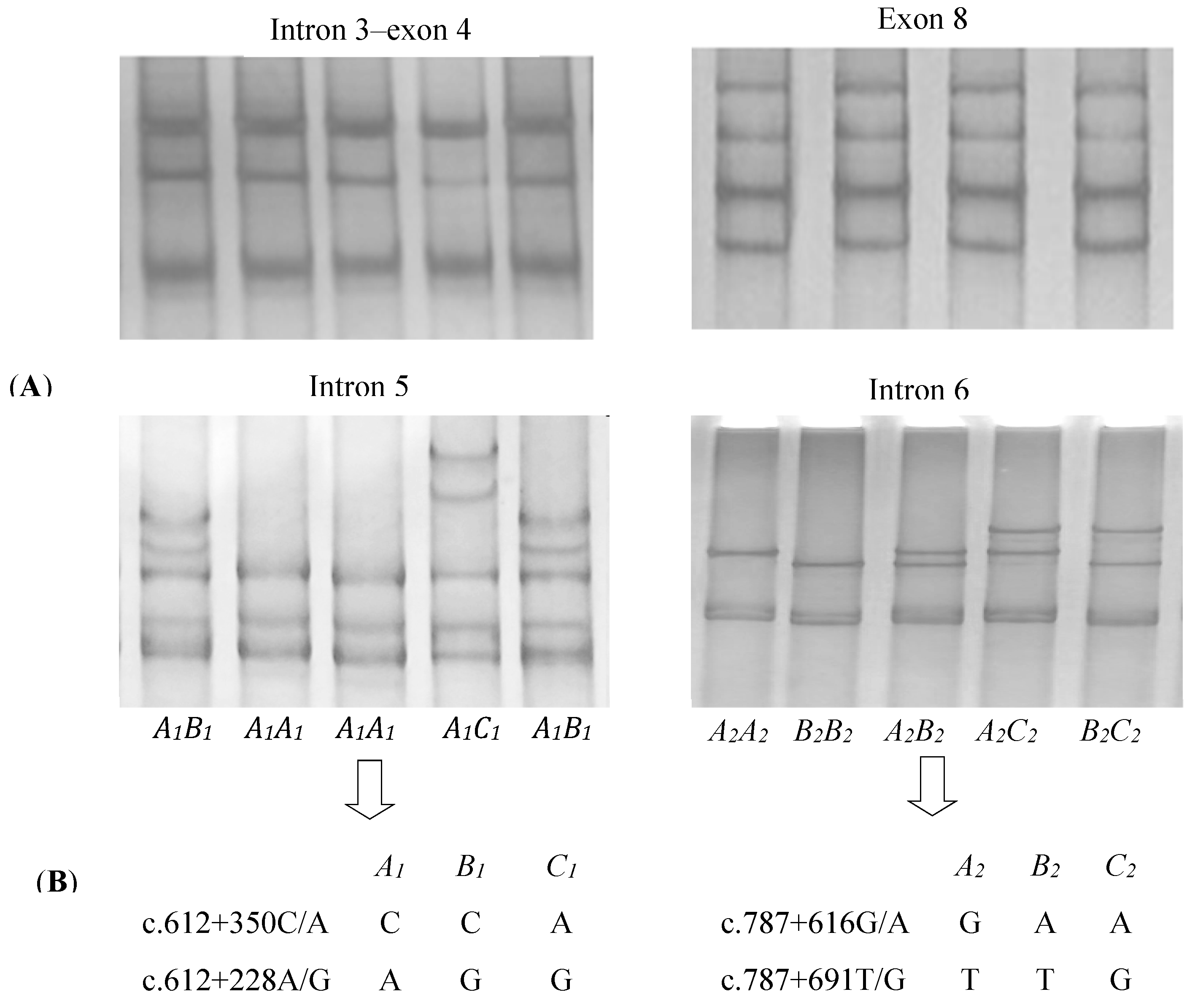
3.1. Identification of Variation in Yak DGAT2
3.2. Polymorphisms in Yak DGAT2
3.3. Associations of DGAT2 Variation with Carcass and Meat Quality Traits in Yak
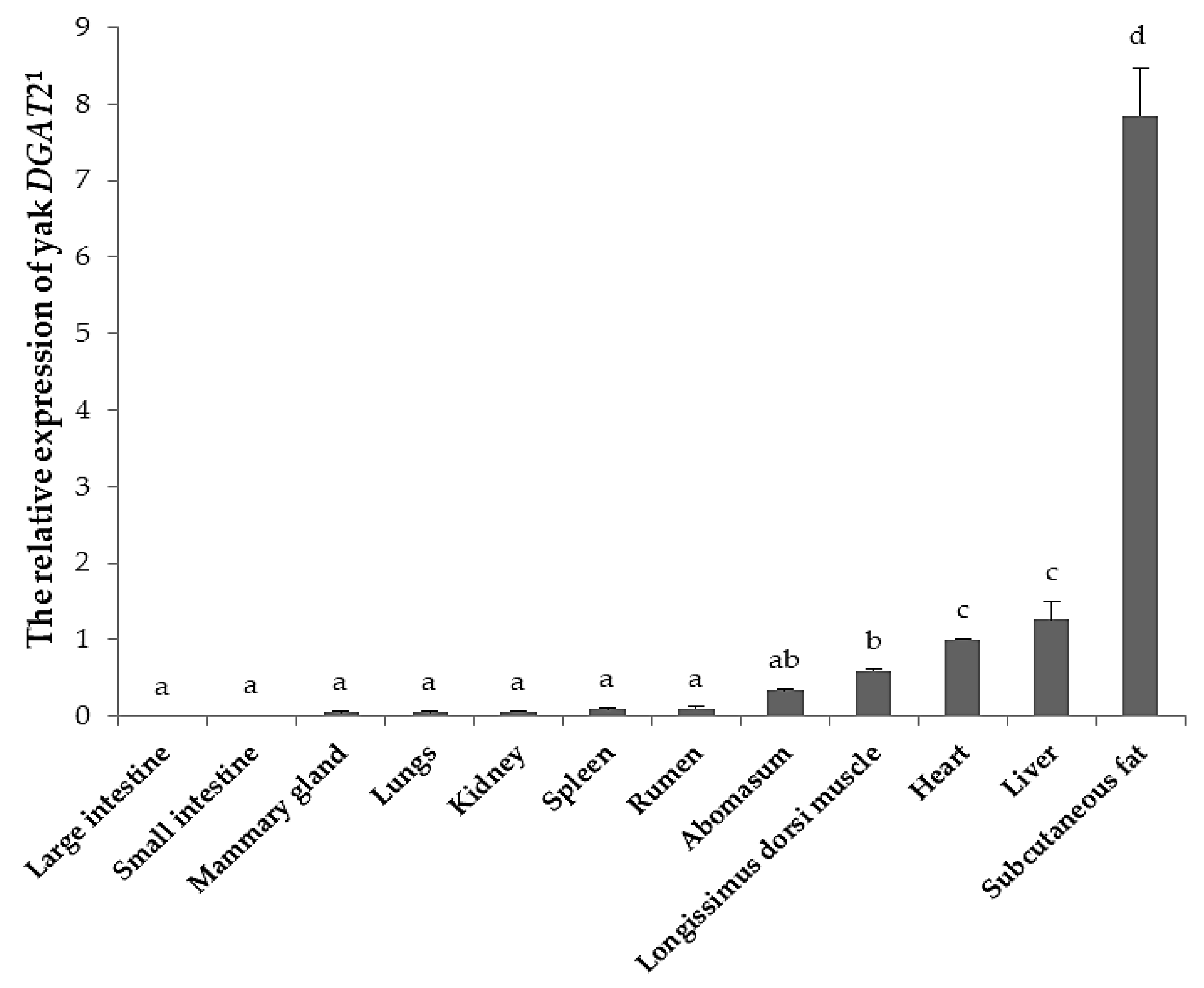
3.4. Tissue Expression of Yak DGAT2
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weiss, S.B.; Kennedy, E.P.; Kiyasu, J.Y. The enzymatic synthesis of triglycerides. J. Biol. Chem. 1960, 235, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Mayorek, N.; Grinstein, I.; Bar-Tana, J. Triacylglycerol synthesis in cultured rat hepatocytes. The rate-limiting role of diacylglycerol acyltransferase. Eur. J. Biochem. 1989, 182, 395–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cases, S.; Smit, S.J.; Zheng, Y.W.; Myers, H.M.; Lear, S.R.; Sande, E.; Novak, S.; Collins, C.; Welchi, B.C.; Lusisi, A.J.; et al. Identification of a gene encoding an acyl CoA: Diacylglycerol cyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. 1998, 95, 13018–13023. [Google Scholar] [CrossRef] [PubMed]
- Cases, S.; Stone, S.J.; Zhou, P.; Yen, E.; Tow, B.; Lardizabal, K.D.; Voelker, T.; Farese, R.V. Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase, and related family members. J. Biol. Chem. 2001, 276, 38870–38876. [Google Scholar] [CrossRef] [PubMed]
- Lardizabal, K.D.; Mai, J.T.; Wagner, N.W.; Wyrick, A.; Voelker, T.; Hawkins, D.J. DGAT2 is a new diacylglycerol acyltransferase gene family: Purification, cloning, and expression in insect cells of two polypeptides from Mortierella ramanniana with diacylglycerol acyltransferase activity. J. Biol. Chem. 2001, 276, 38862–38869. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.J.; Cases, S.; Jensen, D.R.; Chen, H.C.; Sande, E.; Tow, B.; Sanan, D.A.; Raber, J.; Eckel, R.H.; Farese, R.V., Jr. Obesity resistance and multiple mechanisms of triglyceride synthesis in mice lacking DGAT. Nat. Genet. 2000, 25, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Mcfie, P.J.; Banman, S.L.; Curtis Brandtm, C.; Stone, S.J. Diacylglycerol acyltransferase-2 (DGAT2) and monoacylglycerol acyltransferase-2 (MGAT2) interact to promote triacylglycerol synthesis. J. Biol. Chem. 2014, 289, 28237–28248. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.L.; Stone, S.J.; Koliwad, S.; Harris, C.; Farese, R.V., Jr. Thematic review series: Glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J. Lipid Res. 2008, 49, 2283–2301. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.J.; Myers, H.; Brown, B.E.; Watkins, S.M.; Feingold, K.R.; Elias, P.M.; Farese, R.V., Jr. Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J. Biol. Chem. 2004, 279, 11767–11776. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.; Van, E.M.; Bininda-Emonds, O.R.; Habermann, F.A.; Fries, R. Genomic organization of the DGAT2/MOGAT gene family in cattle (Bos taurus) and other mammals. Cytogenet. Genome Res. 2003, 102, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, X.; Zhang, Q.; Wang, X.; Deng, G.; Fang, X.; Gao, X.; Ren, H.; Xu, S. Association between single nucleotide polymorphisms in the DGAT2 gene and beef carcass and quality traits in commercial feedlot steers. Asian Austral. J. Anim. 2009, 22, 943–954. [Google Scholar] [CrossRef]
- Alshuhaib, M.B.S.; Alfihan, R.A.; Alqutbi, A.A.; Althuwaini, T.M. Potential consequences of DGAT2 and BTN genes polymorphism in Iraqi Holstein cattle. Sci. Agric. Bohemica 2017, 48, 127–141. [Google Scholar]
- Jeong, J.; Kwon, E.G.; Im, S.K.; Seo, K.S.; Baik, M. Expression of fat deposition and fat removal genes is associated with intramuscular fat content in longissimus dorsi muscle of Korean cattle steers. J. Anim. Sci. 2012, 90, 2044–2053. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zeng, Y.; Wang, H.; Chen, W.; Du, J.; Chen, Q.; Hu, Y.; Yang, L. The effects of DGAT1 and DGAT2 mRNA expression on fat deposition in fatty and lean breeds of pig. Livest. Sci. 2011, 140, 292–296. [Google Scholar] [CrossRef]
- Yin, Q.; Yang, H.; Han, X.; Fan, B.; Liu, B. Isolation, mapping, SNP detection and association with backfat traits of the porcine CTNNBL1 and DGAT2 genes. Mol. Biol. Rep. 2012, 39, 4485–4490. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Sun, B.; Zhang, X.; Yang, C.; Kang, L.; Zhao, Z.; Jiang, Y. Identification of a 13 bp indel polymorphism in the 3′-UTR of DGAT2 gene associated with backfat thickness and lean percentage in pigs. Gene 2016, 576, 729–733. [Google Scholar]
- An, X.; Song, S.; Hou, J.; Zhua, C.; Penga, J.; Liua, X.; Liua, H.; Xiao, W.; Zhao, H.; Baia, L.; et al. Polymorphism identification in goat DGAT2, gene and association analysis with milk yield and fat percentage. Small Rumin. Res. 2011, 100, 107–112. [Google Scholar] [CrossRef]
- Mao, H.; Dong, X.; Cao, H.; Xu, N.; Yin, Z. Association of DGAT2 gene polymorphisms with carcass and meat quality traits in domestic pigeons (Columba livia). Brit. Poultry Sci. 2018, 59, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Peng, Q.; Tan, C.; Zou, H. Effects of different levels of protein supplementary diet on gene expressions related to intramuscular deposition in early-weaned yaks. Anim. Sci. J. 2014, 85, 411–419. [Google Scholar] [CrossRef]
- Tian, J.; Han, L.; Yu, Q.; Shi, X.; Wang, W. Changes in tenderness and cathepsins activity during post mortem ageing of yak meat. Can. J. Anim. Sci. 2013, 93, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Wiener, G.; Han, J.; Long, R. The Yak, 2nd ed.; FAO Regional Office for Asia and the Pacific Press: Bangkok, Thailand, 2003; p. 5. [Google Scholar]
- Zhou, H.; Hickford, J.G.H.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef]
- Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M. Evaluation of slice shear force as an objective method of assessing beef longissimus tenderness. J. Anim. Sci. 1999, 77, 2693–2699. [Google Scholar] [CrossRef] [PubMed]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 457–477. [Google Scholar] [CrossRef]
- Liu, M.; Peng, J.; Xu, D.; Zheng, R.; Li, F.; Li, J.; Zuo, B.; Lei, M.; Xiong, Y.; Deng, C.; et al. Association of MYF5 and MYOD1 gene polymorphisms and meat quality traits in large whitex meishan F2 pig populations. Biochem. Genet. 2008, 46, 720–732. [Google Scholar] [CrossRef]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, H.; Smyth, A.; Luo, Y.; Hickford, J.G.H. Polymorphism of the bovine ADRB3 gene. Mol. Biol. Rep. 2010, 37, 3389–3392. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethicket, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Devlin, D.J.; Gault, N.F.S.; Moss, B.W.; Tolland, E.; Tollerton, J.; Farmer, L.J.; Gordon, A.W. Factors affecting eating quality of beef. Adv. Animal Biosci. 2017, 8, s2–s5. [Google Scholar] [CrossRef]
- Nishimura, T.; Hattori, A.; Takahashi, K. Structural changes in intramuscular connective tissue during the fatting of Japanese black cattle: Effect of marbling on beef tenderization. J. Anim. Sci. 1999, 77, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, K.; Chiba, H.; Michibata, H.; Seishima, M.; Kawasaki, S.; Okubo, K.; Mitsui, H.; Torii, H.; Mai, Y.I. A novel diacylglycerol acyltransferase (dgat2) is decreased in human psoriatic skin and increased in diabetic mice. Biochem. Bioph. Res. Commun. 2003, 310, 296–302. [Google Scholar] [CrossRef]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Nott, A.; Meislin, S.H.; Moore, M.J. A quantitative analysis of intron effects on mammalian gene expression. RNA 2003, 9, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhang, J.; Xu, H.; Zhang, C.; Du, Y.; Shi, X.; Chen, D.; Sun, J.; Jin, Q.; Lan, X.; et al. Polymorphisms of diacylglycerol acyltransferase 2 gene and their relationship with growth traits in goats. Mol. Bio. Rep. 2012, 39, 1801–1807. [Google Scholar] [CrossRef]
- Snyder, M.W.; Adey, A.; Kitzman, J.O.; Shendure, J. Haplotype-resolved genome sequencing: Experimental methods and applications. Nat. Rev. Genet. 2015, 16, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Akey, J.; Jin, L.; Xiong, M. Haplotypes vs. single marker linkage disequilibrium tests: What do we gain? Eur. J. Hum. Genet. 2001, 9, 291–300. [Google Scholar] [CrossRef]
- Scheike, T.H.; Martinussen, T.; Silver, J.D. Estimating haplotype effects for survival data. Biometrics 2010, 66, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Zollner, S.; von Haeseler, A. Coalescent approach to study linkage disequilibrium between single-nucleotide polymorphisms. Am. J. Hum. Genet. 2000, 66, 615–628. [Google Scholar] [CrossRef]
- Baik, M.; Nguyen, T.H.; Jin, Y.J.; Min, Y.P.; Kang, H.J. Effects of castration on expression of lipid metabolism genes in the liver of Korean cattle. Asian Australas. J. Anim. Sci. 2015, 28, 127–134. [Google Scholar] [CrossRef]
- Van Den Top, A.M.; Wensing, T.; Geelen, M.J.H.; Wentink, G.H.; Van’t Klooster, A.T.; Beynen, A.C. Time trends of plasma lipids and enzymes synthesizing hepatic triacylglycerol during postpartum development of fatty liver in dairy cows. J. Dairy Sci. 1995, 78, 2208–2220. [Google Scholar] [CrossRef]
- BioGPS. Available online: http://biogps.org/sheepatlas/#goto=genereport&id=101110535 (accessed on 28 January 2019).



{kind=link}
{kind=link}
| Region | Primer Sequence (5′→3′) | Amplicon Size (bp) | Annealing Temperature | SSCP Condition |
|---|---|---|---|---|
| Intron 3−exon 4 | TAAGCCTGGGCATGGTTC | 344 | 61 °C | 220 V, 14%, 15 °C |
| CCTCCCAAGATAACACCTGC | ||||
| Intron 5 | TAGGAAACCTTCTCTGACCC | 288 | 64 °C | 260 V, 14%, 12 °C |
| CAGCCACTTAGAAGAACAGC | ||||
| Intron 6 | CCTATGCCAAAGCCTGTCAC | 459 | 64 °C | 240 V, 14%, 10 °C |
| CCCAGACACCAGCCAAACT | ||||
| Exon 8 | CACATCTGGGCCTTTATG | 229 | 63 °C | 220 V, 14%, 15 °C |
| CTTGGCAAGAGGGTTTAGTC |
| Genotype Frequency (%) | Variant Frequency (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Intron 5 | Intron 6 | Intron 5 | Intron 6 | ||||||||||
| A1A1 | A1B1 | A1C1 | A2A2 | B2B2 | A2B2 | A2C2 | B2C2 | A1 | B1 | C1 | A2 | B2 | C2 |
| 39.91 | 42.24 | 15.85 | 47.41 | 24.35 | 7.35 | 11.82 | 9.08 | 69.96 | 22.12 | 7.92 | 65.49 | 24.06 | 10.45 |
| Region | Trait (unit) 2 | Variant | Other Variants in Model | n | Mean ± SE | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Present | Absent | Present | Absent | |||||
| Intron 5 | HCW (kg) | B1 | 417 | 277 | 105.16 ± 2.89 | 107.53 ± 3.01 | 0.4996 | |
| C1 | 110 | 584 | 101.52 ± 4.55 | 106.89 ± 2.42 | 0.2223 | |||
| REA (cm2) | B1 | 417 | 277 | 32.06 ± 0.54 | 32.00 ± 0.59 | 0.9201 | ||
| C1 | 110 | 584 | 31.20 ± 0.86 | 32.16 ± 0.49 | 0.2431 | |||
| WBSF (kg) | B1 | 417 | 277 | 5.12 ± 0.09 | 5.44 ± 0.10 | 0.0020 | ||
| C1 | 110 | 584 | 5.50 ± 0.14 | 5.22 ± 0.08 | 0.0434 | |||
| B1 | C1 | 417 | 277 | 5.25 ± 0.10 | 5.70 ± 0.12 | 0.0001 | ||
| C1 | B1 | 110 | 584 | 5.72 ± 0.15 | 5.23 ± 0.08 | 0.0008 | ||
| DLR (%) | B1 | 417 | 277 | 21.75 ± 0.38 | 22.23 ± 0.41 | 0.2643 | ||
| C1 | 110 | 584 | 20.86 ± 0.59 | 22.12 ± 0.34 | 0.0271 | |||
| CLR (%) | B1 | 417 | 277 | 33.89 ± 0.39 | 34.15 ± 0.42 | 0.5593 | ||
| C1 | 110 | 584 | 33.96 ± 0.62 | 34.01 ± 0.35 | 0.9294 | |||
| Intron 6 | HCW (kg) | A2 | 604 | 90 | 106.74 ± 2.42 | 102.10 ± 4.95 | 0.3383 | |
| B2 | 259 | 435 | 103.92 ± 3.18 | 107.66 ± 2.68 | 0.2711 | |||
| C2 | 145 | 549 | 106.39 ± 3.83 | 106.24 ± 2.50 | 0.9695 | |||
| REA (cm2) | A2 | 604 | 90 | 32.06 ± 0.48 | 31.81 ± 0.95 | 0.7836 | ||
| B2 | 259 | 435 | 31.76 ± 0.62 | 32.19 ± 0.53 | 0.4949 | |||
| C2 | 145 | 549 | 32.90 ± 0.79 | 31.87 ± 0.49 | 0.1691 | |||
| WBSF (kg) | A2 | 604 | 90 | 5.27 ± 0.08 | 5.20 ± 0.16 | 0.6429 | ||
| B2 | 259 | 435 | 5.37 ± 0.10 | 5.20 ± 0.09 | 0.1132 | |||
| C2 | 145 | 549 | 5.05 ± 0.13 | 5.30 ± 0.08 | 0.0441 | |||
| B2 | C2 | 259 | 435 | 5.29 ± 0.11 | 5.10 ± 0.10 | 0.0853 | ||
| C2 | B2 | 145 | 549 | 5.06 ± 0.13 | 5.33 ± 0.08 | 0.0339 | ||
| DLR (%) | A2 | 604 | 90 | 21.96 ± 0.33 | 21.94 ± 0.66 | 0.9752 | ||
| B2 | 259 | 435 | 21.54 ± 0.43 | 22.19 ± 0.36 | 0.1327 | |||
| C2 | 145 | 549 | 22.61 ± 0.54 | 21.83 ± 0.34 | 0.1288 | |||
| CLR (%) | A2 | 604 | 90 | 34.04 ± 0.34 | 33.60 ± 0.68 | 0.4966 | ||
| B2 | 259 | 435 | 33.30 ± 0.44 | 34.40 ± 0.37 | 0.0142 | |||
| C2 | 145 | 549 | 34.50 ± 0.56 | 33.90 ± 0.35 | 0.2606 | |||
| Trait (unit) 2 | Haplotype | Other Haplotypes in Model | n | Mean ± SE | p-Value | ||
|---|---|---|---|---|---|---|---|
| Present | Absent | Present | Absent | ||||
| HCW (kg) | A1-A2 | 428 | 41 | 107.90 ± 2.82 | 94.74 ± 8.63 | 0.1234 | |
| A1-B2 | 99 | 370 | 99.74 ± 5.20 | 109.33 ± 3.01 | 0.0832 | ||
| A1-C2 | 31 | 438 | 108.42 ± 11.95 | 107.35 ± 2.83 | 0.9281 | ||
| B1-A2 | 133 | 336 | 108.81 ± 4.72 | 106.90 ± 3.08 | 0.7051 | ||
| C1-A2 | 41 | 428 | 100.04 ± 11.86 | 107.63 ± 2.85 | 0.5254 | ||
| REA (cm2) | A1-A2 | 428 | 41 | 31.89 ± 0.58 | 33.11 ± 1.40 | 0.3644 | |
| A1-B2 | 99 | 370 | 32.38 ± 0.93 | 31.83 ± 0.61 | 0.5587 | ||
| A1-C2 | 31 | 438 | 34.19 ± 1.58 | 31.87 ± 0.57 | 0.1283 | ||
| B1-A2 | 133 | 336 | 32.05 ± 0.83 | 31.91 ± 0.62 | 0.8651 | ||
| C1-A2 | 41 | 428 | 32.97 ± 1.35 | 31.84 ± 0.59 | 0.4051 | ||
| WBSF (kg) | A1-A2 | 428 | 41 | 5.29 ± 0.10 | 4.76 ± 0.23 | 0.0191 | |
| A1-B2 | 99 | 370 | 5.24 ± 0.16 | 5.27 ± 0.10 | 0.8518 | ||
| A1-C2 | 31 | 438 | 5.28 ± 0.27 | 5.26 ± 0.10 | 0.9506 | ||
| B1-A2 | 133 | 336 | 4.93 ± 0.14 | 5.39 ± 0.10 | 0.0010 | ||
| C1-A2 | 41 | 428 | 5.48 ± 0.23 | 5.24 ± 0.10 | 0.2903 | ||
| A1-A2 | B1-A2 | 428 | 41 | 5.18 ± 0.10 | 4.48 ± 0.24 | 0.0021 | |
| B1-A2 | A1-A2 | 133 | 336 | 4.56 ± 0.18 | 5.10 ± 0.14 | 0.0001 | |
| DLR (%) | A1-A2 | 428 | 41 | 22.23 ± 0.41 | 22.20 ± 0.99 | 0.9756 | |
| A1-B2 | 99 | 370 | 22.37 ± 0.66 | 22.19 ± 0.43 | 0.7854 | ||
| A1-C2 | 31 | 438 | 23.51 ± 1.12 | 22.18 ± 0.41 | 0.2186 | ||
| B1-A2 | 133 | 336 | 22.71 ± 0.59 | 22.04 ± 0.44 | 0.2580 | ||
| C1-A2 | 41 | 428 | 20.83 ± 0.96 | 22.38 ± 0.42 | 0.1055 | ||
| CLR (%) | A1-A2 | 428 | 41 | 34.02 ± 0.40 | 34.03 ± 0.98 | 0.9925 | |
| A1-B2 | 99 | 370 | 33.37 ± 0.65 | 34.20 ± 0.42 | 0.2058 | ||
| A1-C2 | 31 | 438 | 34.66 ± 1.11 | 34.00 ± 0.40 | 0.5360 | ||
| B1-A2 | 133 | 336 | 34.17 ± 0.58 | 33.97 ± 0.43 | 0.7337 | ||
| C1-A2 | 41 | 428 | 33.51 ± 0.94 | 34.08 ± 0.41 | 0.5513 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Shi, B.; Xie, J.; Zhou, H.; Wang, J.; Liu, X.; Li, S.; Zhao, Z.; Luo, Y. Tissue Expression and Variation of the DGAT2 Gene and Its Effect on Carcass and Meat Quality Traits in Yak. Animals 2019, 9, 61. https://doi.org/10.3390/ani9020061
Hu J, Shi B, Xie J, Zhou H, Wang J, Liu X, Li S, Zhao Z, Luo Y. Tissue Expression and Variation of the DGAT2 Gene and Its Effect on Carcass and Meat Quality Traits in Yak. Animals. 2019; 9(2):61. https://doi.org/10.3390/ani9020061
Chicago/Turabian StyleHu, Jiang, Bingang Shi, Jianpeng Xie, Huitong Zhou, Jiqing Wang, Xiu Liu, Shaobin Li, Zhidong Zhao, and Yuzhu Luo. 2019. "Tissue Expression and Variation of the DGAT2 Gene and Its Effect on Carcass and Meat Quality Traits in Yak" Animals 9, no. 2: 61. https://doi.org/10.3390/ani9020061