Ovariectomy Impairs Socio-Cognitive Functions in Dogs

 , ,
, , 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Procedure
2.3. Data Analysis
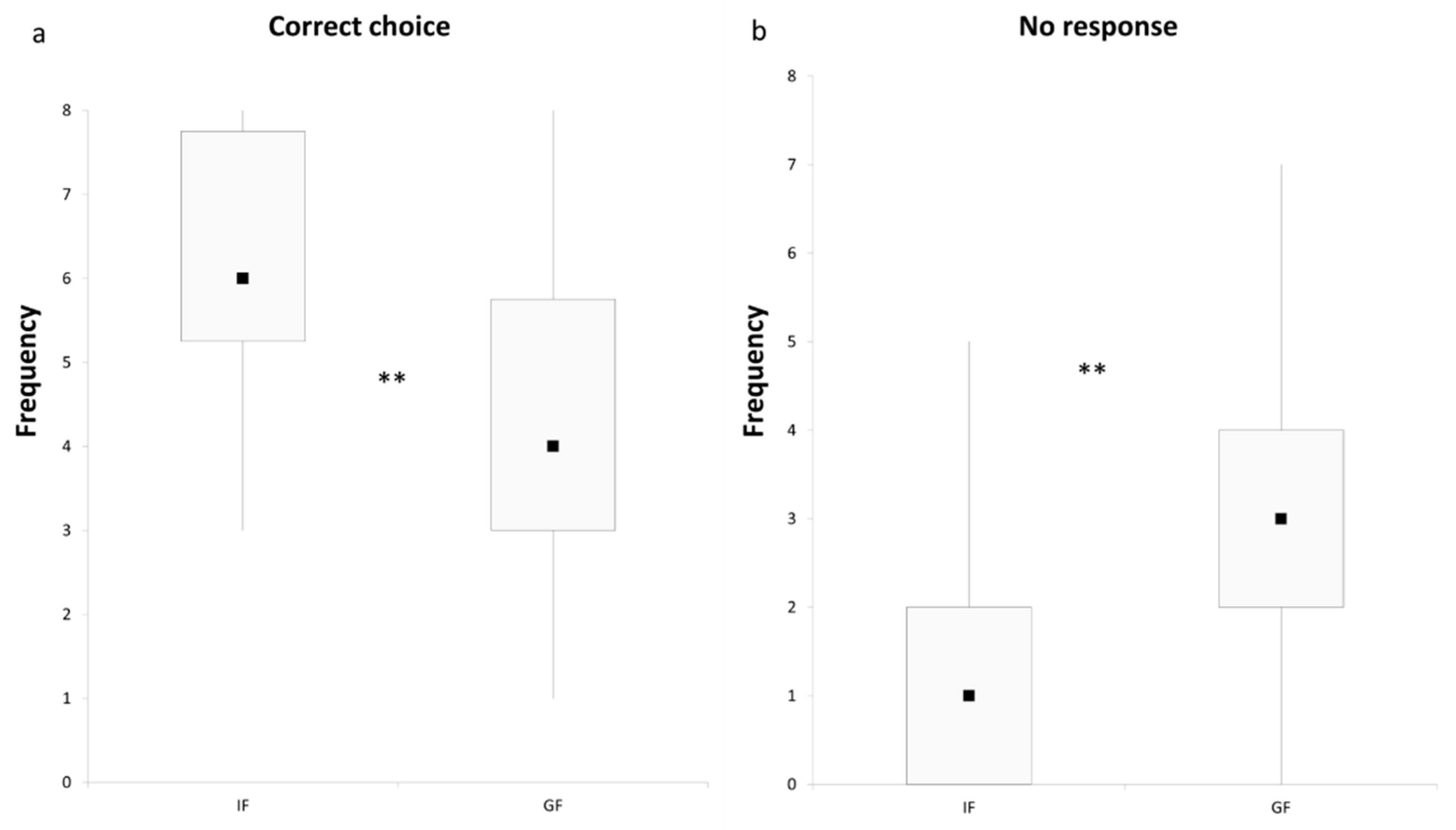
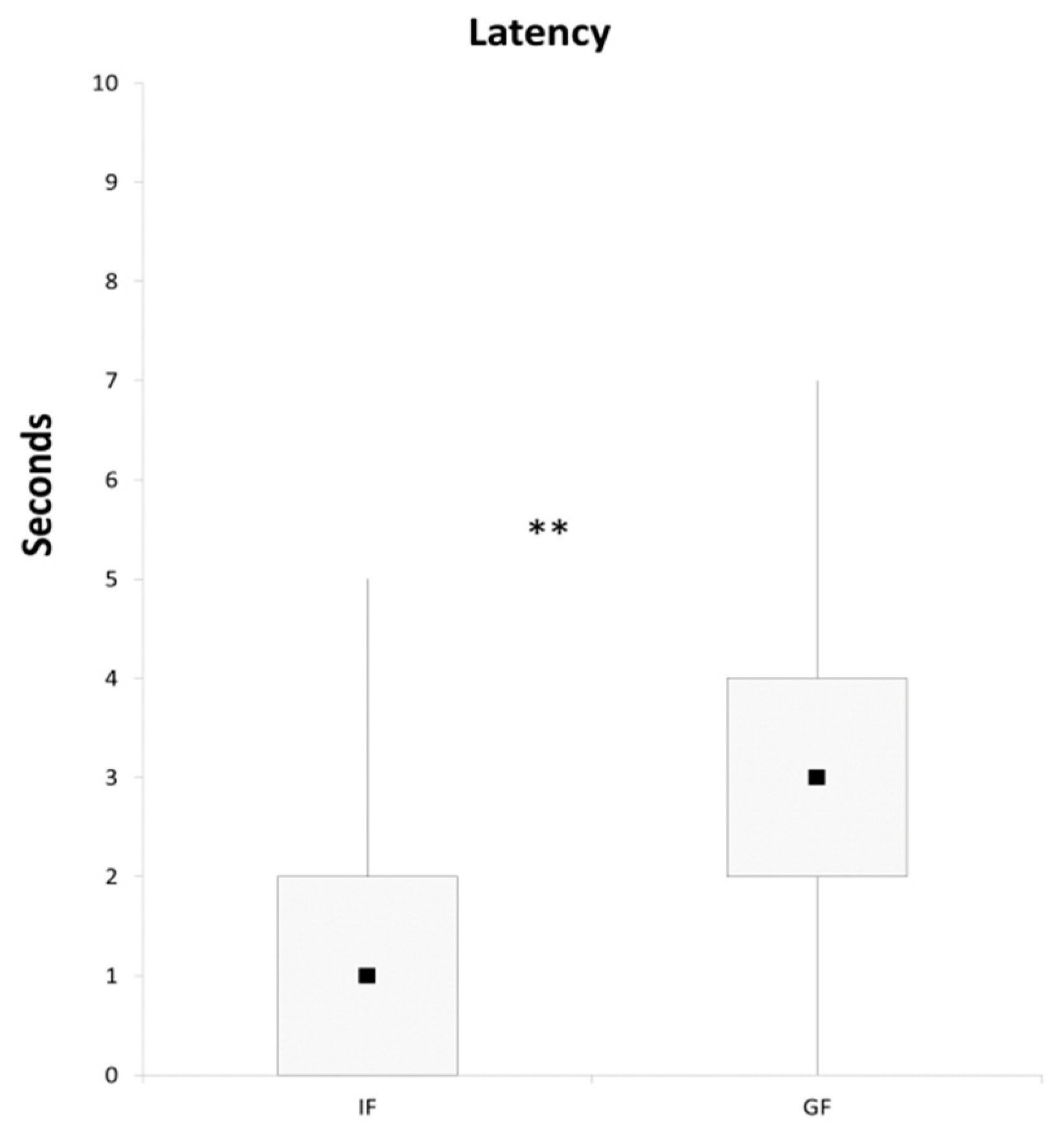
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Frick, K.M.; Kim, J.; Koss, W.A. Estradiol and hippocampal memory in female and male rodents. Curr. Opin. Behav. Sci. 2018, 23, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Scandurra, A.; Marinelli, L.; Lõoke, M.; D’Aniello, B.; Mongillo, P. The effect of age, sex and gonadectomy on dogs’ use of spatial navigation strategies. Appl. Anim. Behav. Sci. 2018, 205, 89–97. [Google Scholar] [CrossRef]
- Mongillo, P.; Scandurra, A.; D’Aniello, B.; Marinelli, L. Effect of sex and gonadectomy on dogs’ spatial performance. Appl. Anim. Behav. Sci. 2017, 191, 84–89. [Google Scholar] [CrossRef]
- Mongillo, P.; Pitteri, E.; Candaten, M.; Marinelli, L. Can attention be taught? Interspecific attention by dogs (Canis familiaris) performing obedience tasks. Appl. Anim. Behav. Sci. 2016, 182, 30–37. [Google Scholar] [CrossRef]
- D’Aniello, B.; Scandurra, A.; Alterisio, A.; Valsecchi, P.; Prato-Previde, E. The importance of gestural communication: A study of human-dog communication using incongruent information. Anim. Cogn. 2016, 19, 1231–1235. [Google Scholar] [CrossRef]
- Scandurra, A.; Alterisio, A.; Marinelli, L.; Mongillo, P.; Semin, G.R.; D’Aniello, B. Effectiveness of verbal and gestural signals and familiarity with signal-senders on the performance of working dogs. Appl. Anim. Behav. Sci. 2017, 191, 78–83. [Google Scholar] [CrossRef]
- Scandurra, A.; Alterisio, A.; Aria, M.; Vernese, R.; D’Aniello, B. Should I fetch one or the other? A study on dogs on the object choice in the bimodal contrasting paradigm. Anim. Cogn. 2018, 21, 119–126. [Google Scholar] [CrossRef]
- Hare, B.; Brown, M.; Williamson, C.; Tomasello, M. The domestication of social cognition in dogs. Science 2002, 298, 1634–1636. [Google Scholar] [CrossRef]
- Wynne, C.D.L.; Udell, M.A.R.; Lord, K.A. Ontogeny’s impact on human-dog communication. Anim. Behav. 2008, 76, e1–e4. [Google Scholar] [CrossRef]
- D’Aniello, B.; Alterisio, A.; Scandurra, A.; Petremolo, E.; Iommelli, M.R.; Aria, M. What’s the point? Golden and Labrador retrievers living in kennels do not understand human pointing gestures. Anim. Cogn. 2017, 20, 777–787. [Google Scholar] [CrossRef]
- D’Aniello, B.; Scandurra, A. Ontogenetic effects on gazing behaviour: A case study of kennel dogs (Labrador Retrievers) in the impossible task paradigm. Anim. Cogn. 2016, 19, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Svartberg, K. Breed-typical behaviour in dogs—Historical remnants or recent constructs? Appl. Anim. Behav. Sci. 2006, 96, 293–313. [Google Scholar] [CrossRef] [Green Version]
- Starling, M.J.; Branson, N.; Thomson, P.C.; McGreevy, P.D. Age, sex and reproductive status affect boldness in dogs. Vet. J. 2013, 197, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Arden, R.; Adams, M.J. A general intelligence factor in dogs. Intelligence 2016, 55, 79–85. [Google Scholar] [CrossRef]
- Beatty, W.W. Gonadal hormones and sex differences in non-reproductive behaviors in rodents: Organizational and activational influences. Horm. Behav. 1979, 12, 112–163. [Google Scholar] [CrossRef]
- Nene, A.S.; Pazare, P.A. A study of auditory reaction time in different phases of the normal menstrual cycle. Indian J. Physiol. Pharmacol. 2010, 54, 386–390. [Google Scholar]
- Niruba, R.; Maruthy, K.N. Assessment of auditory and visual reaction time in type 2 diabetics-A case control study. Al Ameen J. Med. Sci. 2011, 4, 274–279. [Google Scholar]
- Nikam, L.H.; Gadkari, J.V. Effect of age, gender and body mass index on visual and auditory reaction times in Indian population. Indian J. Physiol. Pharmacol. 2012, 56, 94–99. [Google Scholar]
- Frank, L.A.; Rohrbach, B.W.; Bailey, E.M.; West, J.R.; Oliver, J.W. Steroid hormone concentration profiles in healthy intact and neutered dogs before and after cosyntropin administration. Domest. Anim. Endocrinol. 2003, 24, 43–57. [Google Scholar] [CrossRef]
- Martin, R.B.; Butcher, R.L.; Sherwood, L.L.; Buckendahl, P.; Boyd, R.D.; Farris, D.; Sharkey, N.; Dannucci, G. Effects of ovariectomy in beagle dogs. Bone 1987, 8, 23–31. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenesis: Unanswered questions. Trends Endocrinol. Metabol. 2017, 28, 771–793. [Google Scholar] [CrossRef] [PubMed]
- De Lisa, E.; Paolucci, M.; Di Cosmo, A. Conservative nature of oestradiol signalling pathways in the brain lobes of Octopus vulgaris involved in reproduction, learning and motor coordination. J. Neuroendocrinol. 2011, 24, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Caruso, D.; Pesaresi, M.; Maschi, O.; Giatti, S.; Garcia-Segura, L.M.; Melcandi, R.C. Effect of short-and long-term gonadectomy on neuroactive steroid levels in the central and peripheral nervous system of male and female rats. J. Neuroendocrinol. 2010, 22, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Hojo, Y.; Higo, S.; Komatsuzaki, Y.; Murakami, G.; Yoshino, H.; Uebayashi, M.; Kawato, S. Female hippocampal estrogens have a significant correlation with cyclic fluctuation of hippocampal spines. Front. Neural Circuits 2013, 7, 149. [Google Scholar] [CrossRef] [PubMed]
- Luine, V. Neuroendocrinology of memory and cognition. In Handbook of Neurochemistry and Molecular Neurobiology; Behavioral Neurochemistry and Neuroendocrinology; Lajtha, A., Jeffrey, D.B., Eds.; Springer: Berlin, Germany, 2006; pp. 775–800. [Google Scholar]
- Fazio, E.; Medica, P.; Cravana, C.; Pupillo, A.; Ferlazzo, A. Effects of ovariohysterectomy in dogs and cats on adrenocortical, haematological and behavioural parameters. Acta Sci. Vet. 2015, 43, 1339. [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scandurra, A.; Alterisio, A.; Di Cosmo, A.; D’Ambrosio, A.; D’Aniello, B. Ovariectomy Impairs Socio-Cognitive Functions in Dogs. Animals 2019, 9, 58. https://doi.org/10.3390/ani9020058
Scandurra A, Alterisio A, Di Cosmo A, D’Ambrosio A, D’Aniello B. Ovariectomy Impairs Socio-Cognitive Functions in Dogs. Animals. 2019; 9(2):58. https://doi.org/10.3390/ani9020058
Chicago/Turabian StyleScandurra, Anna, Alessandra Alterisio, Anna Di Cosmo, Antonio D’Ambrosio, and Biagio D’Aniello. 2019. "Ovariectomy Impairs Socio-Cognitive Functions in Dogs" Animals 9, no. 2: 58. https://doi.org/10.3390/ani9020058