Extracts from Microalga Chlorella sorokiniana Exert an Anti-Proliferative Effect and Modulate Cytokines in Sheep Peripheral Blood Mononuclear Cells

 , , , and
, , , and 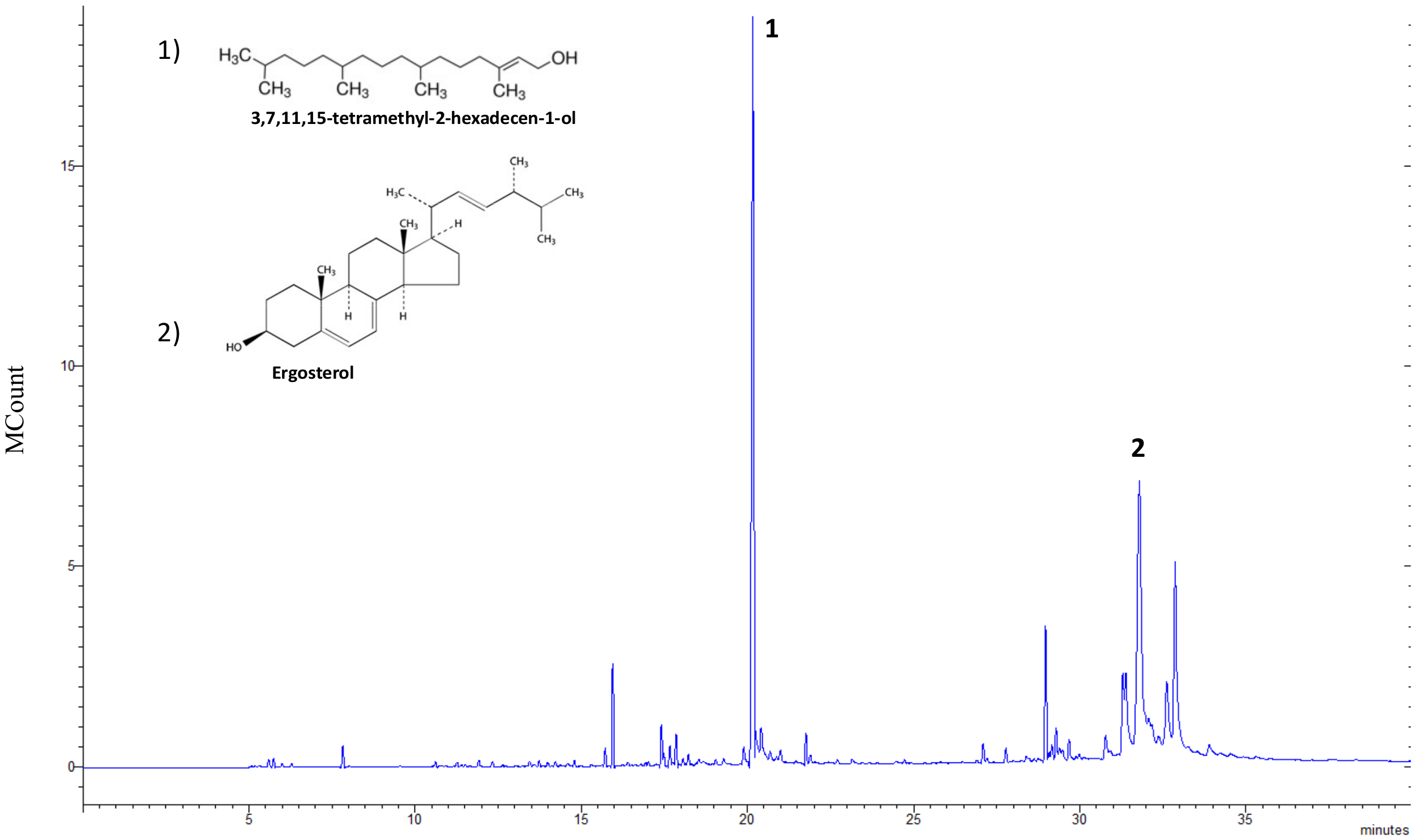{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgae Cultivation
2.2. Microalgal Extract Preparation and Chemical Characterization
2.3. Animals and Experimental Treatments
2.4. PBMCs for Lymphocyte Stimulation Assay and Cytokine Determination
2.5. Determination of Cytokines in Culture Supernatant by ELISA
2.6. Statistical Analysis
2.7. Ethical approval
3. Results
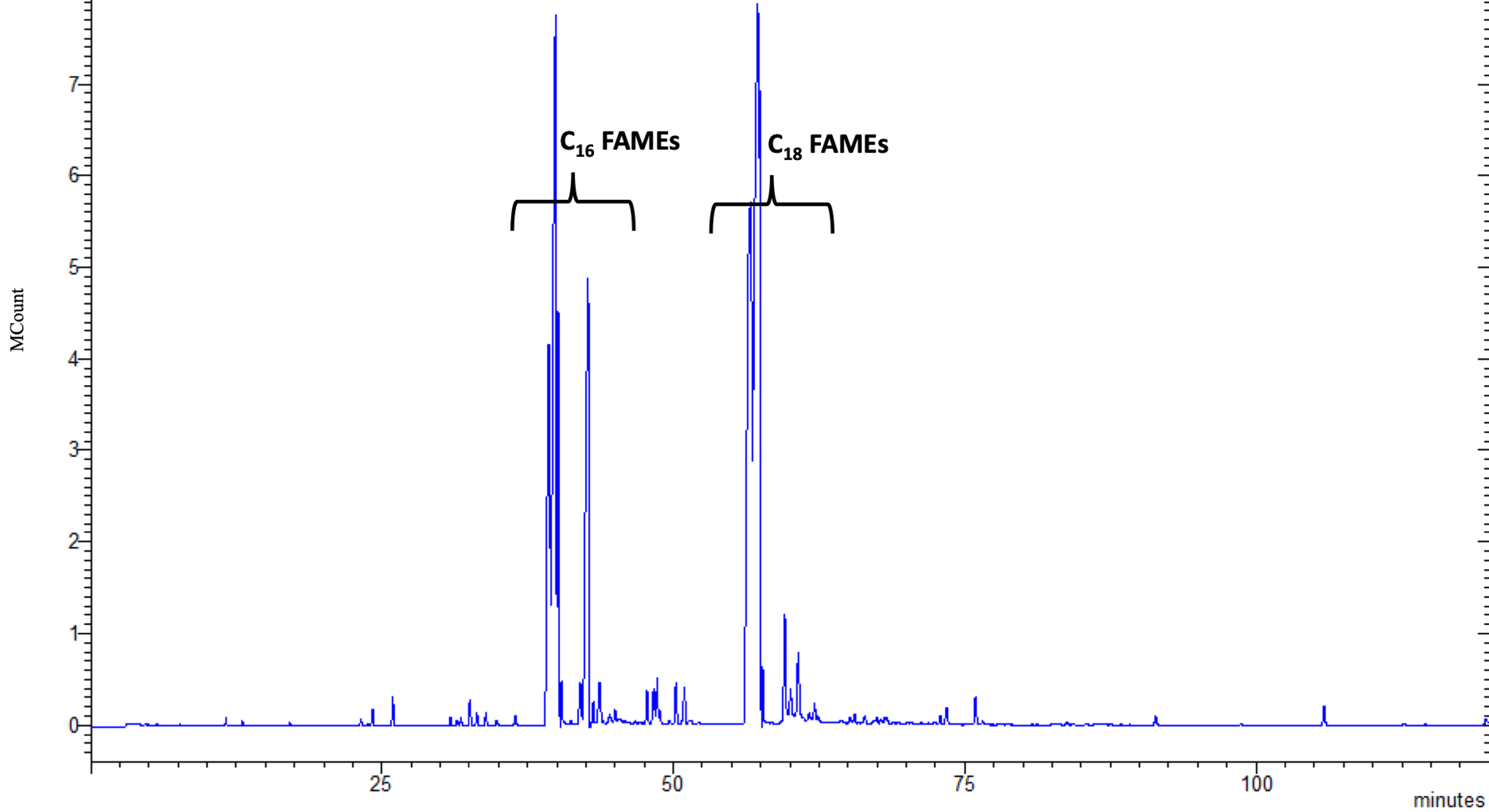
3.1. Chemical Characterization of Microalgal Extracts
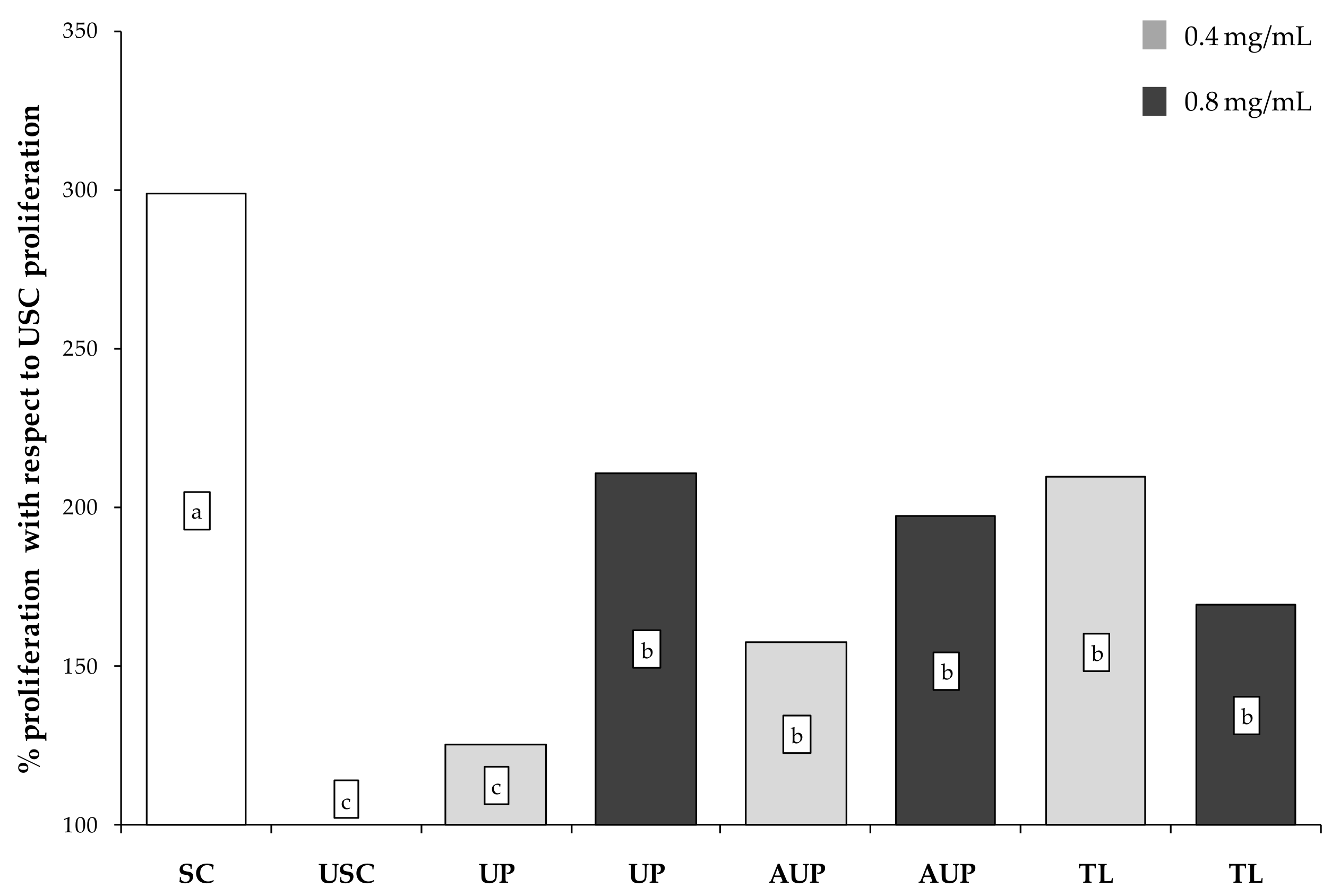
3.2. Proliferative Response to Clorella sorokiniana Extracts
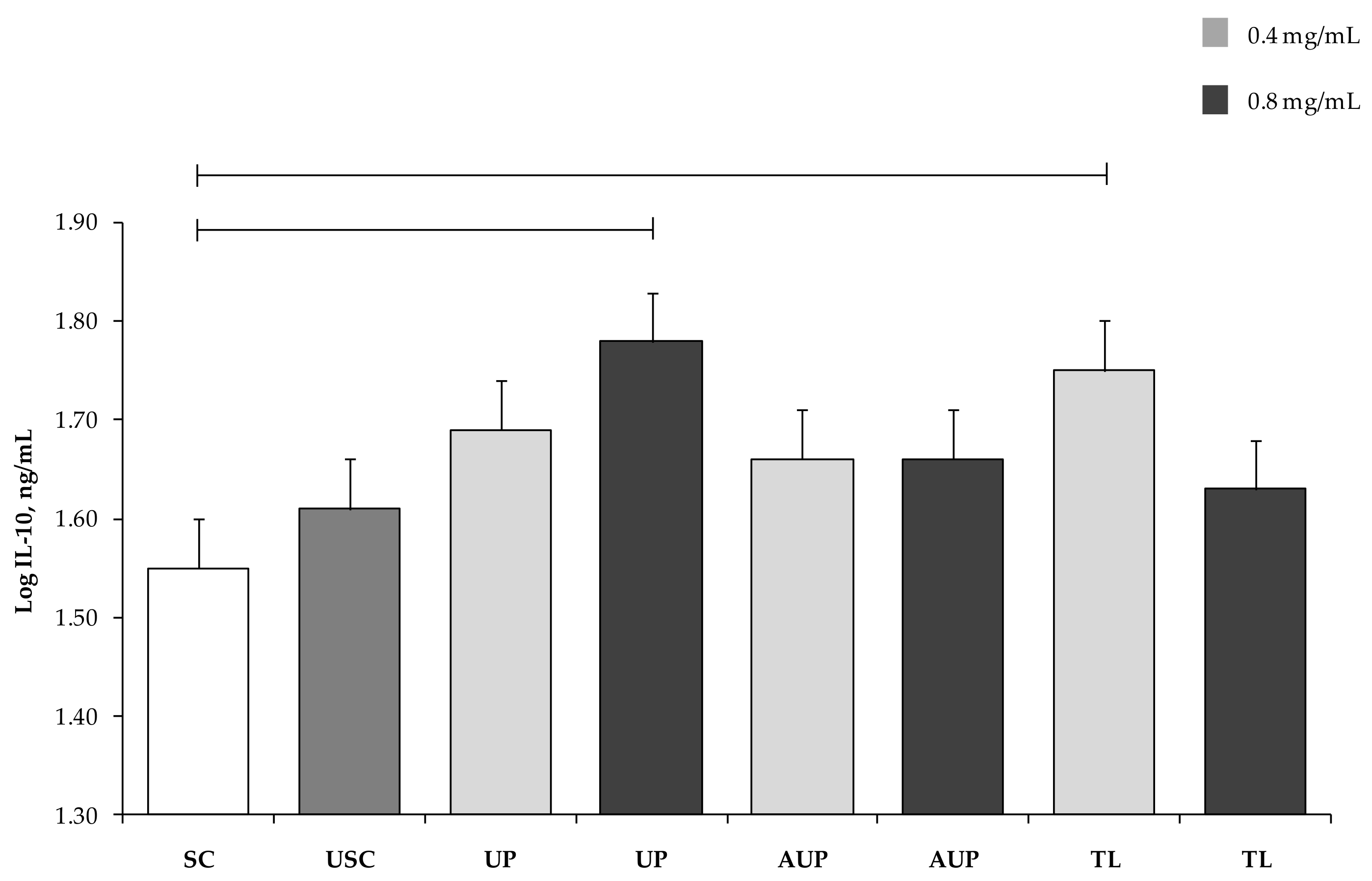
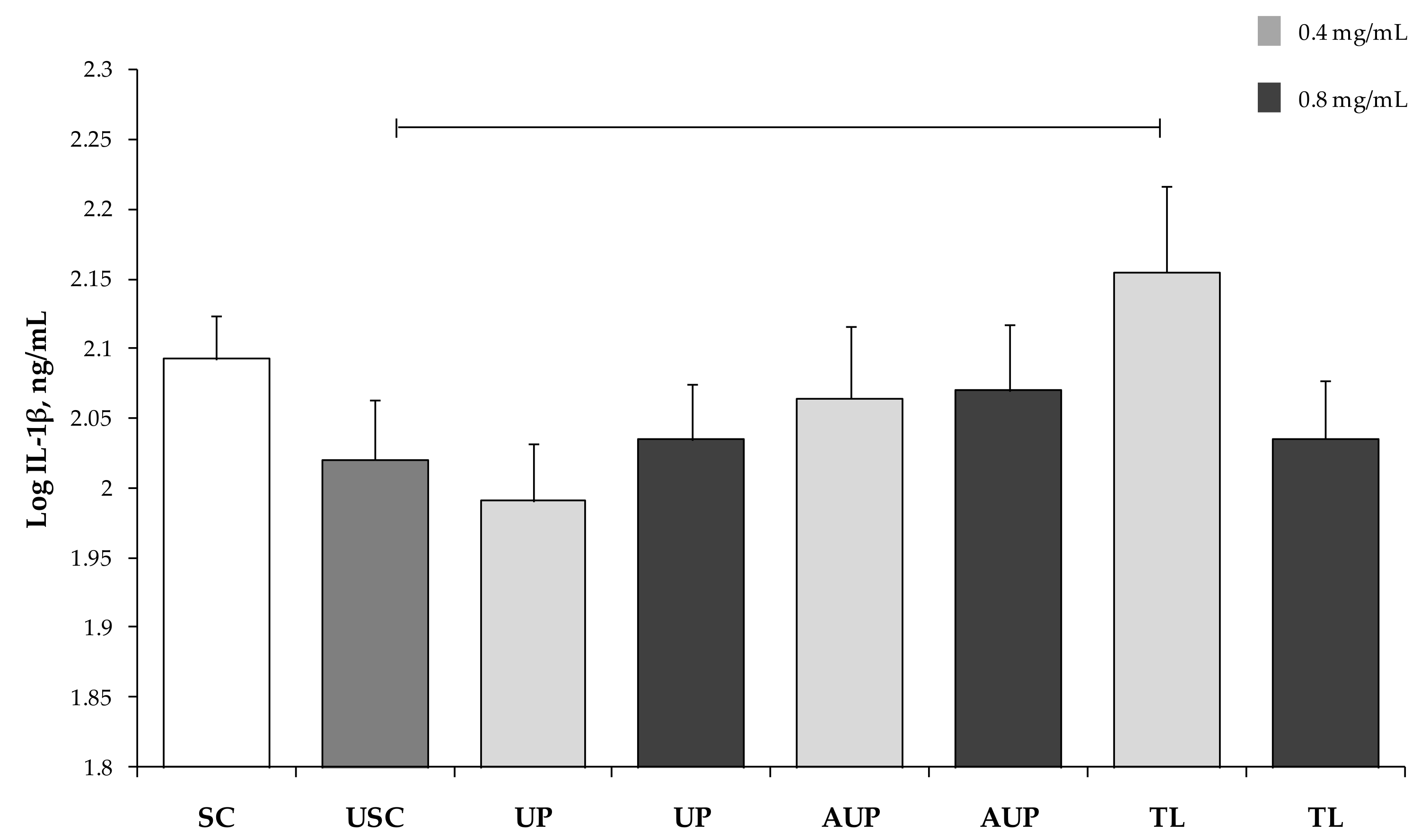
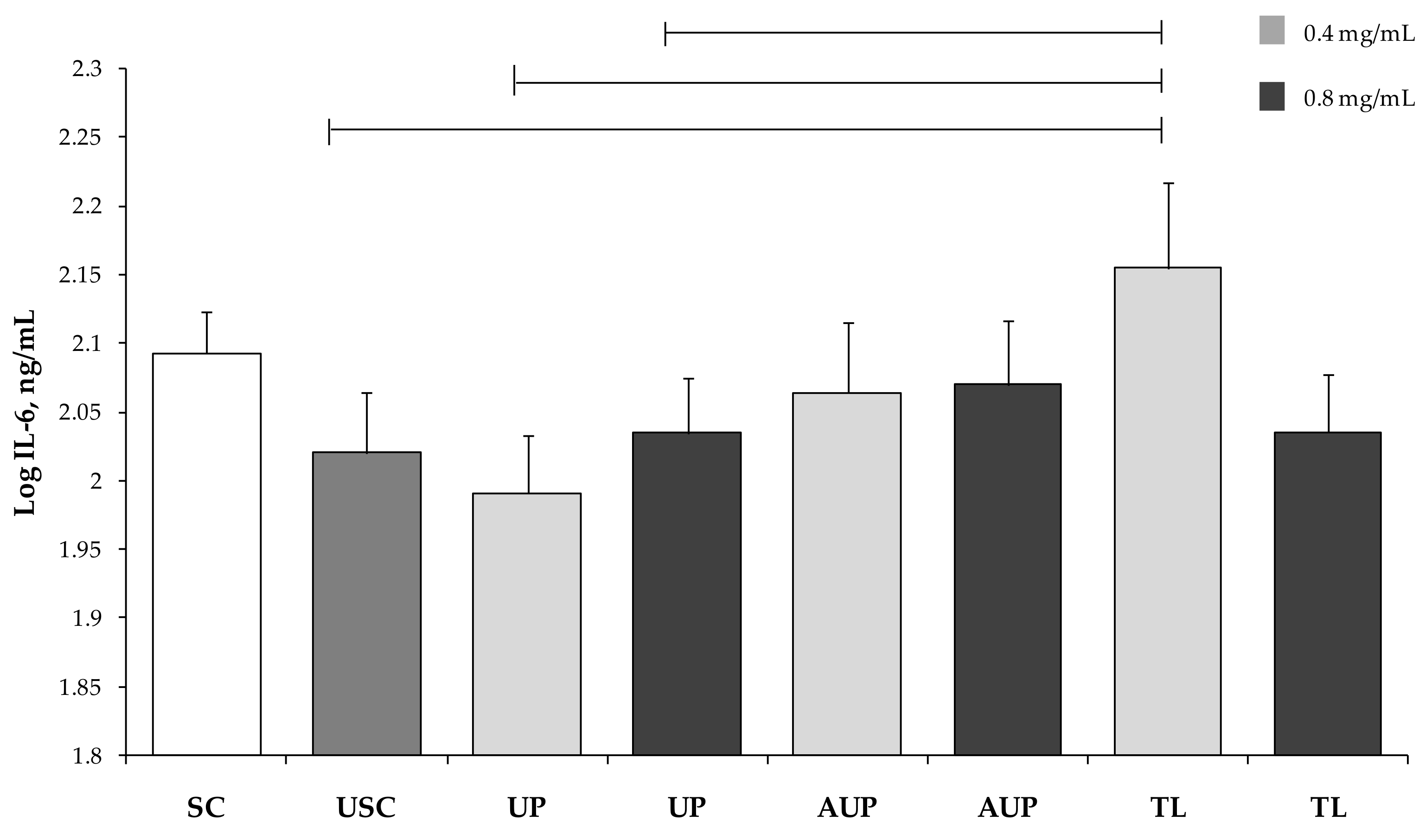
3.3. Cytokine Production by PBMCs
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Shields, R.J.; Lupatsch, I. Algae for aquaculture and animal feeds. J. Anim. Sci. 2012, 4, 23–37. [Google Scholar]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2017. [Google Scholar] [CrossRef]
- Torello, C.O.; Martins, F.; de Castro, T.C.L.; Saad, S.T.O.; Saad, M.J.A.; Queiroz, M.L.S. Chlorella modulation of cytokine production in obese mice and minireview of the effects of the alga in obesity. EC Nutr. 2016, 5.1, 1037–1045. [Google Scholar]
- Talero, E.; Mauriño, S.G.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Soletto, D.; Binaghi, L.; Lodi, A.; Carvalho, J.C.M.; Converti, A. Batch and fedbatch cultivations of Spirulina platensis using ammonium sulphate and urea as nitrogen sources. Aquaculture 2005, 243, 217–224. [Google Scholar] [CrossRef]
- Rangel-Yagui, C.O.; Godoy-Danesi, E.D.; Carvalho, J.C.M.; Sato, S. Chlorophyll production from Spirulina platensis: Cultivation with urea addition by fed-batch process. Bioresour. Technol. 2004, 92, 114–133. [Google Scholar] [CrossRef]
- Dong, T.; Wang, J.; Miao, C.; Zheng, Y.; Chen, S. Two-step in situ biodiesel production from microalgae with high free fatty acid content. Bioresour. Technol. 2013, 136, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Doughman, S.D.; Krupanidhi, S.; Sanjeevi, C.B. ω-3 Fatty acids for nutrition and medicine: Considering microalgae oil as a vegetarian source of EPA and DHA. Curr. Diabetes Rev. 2007, 3, 198–203. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef]
- Ciliberti, M.G.; Francavilla, M.; Intini, S.; Albenzio, M.; Marino, R.; Santillo, A.; Caroprese, M. Phytosterols from Dunaliella tertiolecta reduce cell proliferation in sheep fed flaxseed during postpartum. Mar. Drugs 2017, 15, 216. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Ciliberti, M.G.; Francavilla, M.; Sevi, A. A mixture of phytosterols from Dunaliella tertiolecta affects proliferation of peripheral blood mononuclear cells and cytokine production in sheep. Vet. Immunol. Immunopathol. 2012, 150, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Morgese, M.G.; Mhillaj, E.; Francavilla, M.; Bove, M.; Morgano, L.; Tucci, P.; Trabace, L.; Schiavone, S. Chlorella sorokiniana extract improves short-term memory in rats. Molecules 2016, 21, 1311. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, M.; Colaianna, M.; Zotti, M.; Morgese, M.G.; Trotta, P.; Tucci, P.; Schiavone, S.; Cuomo, V.; Trabace, L. Extraction, characterization and in vivo neuromodulatory activity of phytosterols from microalga Dunaliella tertiolecta. Curr. Med. Chem. 2012, 19, 3058–3067. [Google Scholar] [CrossRef]
- Meshram, G.A.; Patil, V.D. Simple and efficient method for acetylation of alcohols, phenols, amines, and thiols using anhydrous NiCl2 under solvent-free conditions. Synth. Commun. 2009, 39, 2516–2528. [Google Scholar] [CrossRef]
- Wattegedera, S.; Sills, K.; Howard, C.J.; Hope, J.C.; McInnes, C.J.; Entrican, G. Variability in cytokine production and cell proliferation by mitogen-activate ovine peripheral blood mononuclear cells: Modulation by interleukin (IL)-10 and IL-12. Vet. Immunol. Immunopathol. 2004, 102, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1965, 52, 591–601. [Google Scholar] [CrossRef]
- SAS Institute. SAS Enterprise Guide: Statistics, version 6.1; SAS Inst. Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Chou, N.T.; Cheng, C.F.; Wu, H.C.; Lai, C.P.; Lin, L.T.; Pan, I.H.; Ko, C.H. Chlorella sorokiniana-induced activation and maturation of human monocyte-derived dendritic cells through NF-κB and PI3K/MAPK pathways. J. Evid. Based Complement. Altern. Med. 2012, 2012, 735396. [Google Scholar] [CrossRef] [PubMed]
- Merchant, R.E.; Andre, C.A. A review of recent clinical trials of the nutritional supplement Chlorella pyrenoidosa in the treatment of fibromyalgia, hypertension, and ulcerative colitis. Altern. Ther. Health Med. 2001, 7, 79–91. [Google Scholar]
- Yang, F.; Shi, Y.; Sheng, J.; Hu, Q. In vivo immunomodulatory activity of polysaccharides derived from Chlorella pyrenoidosa. Eur. Food Res. Technol. 2006, 224, 225–228. [Google Scholar] [CrossRef]
- An, H.J.; Rim, H.K.; Jeong, H.J.; Hong, S.H.; Um, J.Y.; Kim, H.M. Hot water extracts of Chlorella vulgaris improve immune function in protein-deficient weanling mice and immune cells. Immunopharmacol. Immunotoxicol. 2010, 32, 585–592. [Google Scholar] [CrossRef]
- Lin, Y.L.; Liang, Y.C.; Lee, S.S.; Chiang, B.L. Polysaccharide purified from Ganoderma lucidum induced activation and maturation of human monocyte-derived dendritic cells by the NF-κB and p38 mitogen-activated protein kinase pathways. J. Leukoc. Biol. 2005, 78, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Bai, S.C.; Koo, J.W.; Wang, X.; Kim, S.K. Effects of dietary Chlorella Ellipsoidea supplementation on growth, blood characteristics, and whole- body composition in juvenile Japanese flounder Paralichthys olivaceus. J. World Aquac. Soc. 2002, 2002. 33, 425–431. [Google Scholar] [CrossRef]
- Suárez, E.R.; Kralovec, J.A.; Grindley, T.B. Isolation of phosphorylated polysaccharides from algae: The immunostimulatory principle of Chlorella pyrenoidosa. Carbohydr. Res. 2010, 345, 1190–1204. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Matsueda, T.; Lida, T.; Hasegawa, T. Chlorella accelerates dioxin excretion in rats. J. Nutr. 1999, 129, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Yamada, A.; Nada, K.; Shoyama, Y.; Kubo, C.; Nomoto, K. Oral administration of a unicellular green algae, Chlorella vulgaris, prevents stress-induced ulcer. Planta Med. 1997, 63, 465–466. [Google Scholar] [CrossRef] [PubMed]
- Jong-Yuh, C.; Mei-Fen, S. Potential hypoglycemic effects of Chlorella in streptozotocin-induced diabetic mice. Life Sci. 2005, 77, 980–990. [Google Scholar] [CrossRef]
- Zhao, J.C.; Zhang, Y.; Xu, D.M.; Huang, G.Q.; Xu, Y.L.; Wang, Z.Y.; Fang, S.D.; Chen, Y.; Gu, Y.L. The anti-atherogenic effects of components isolated from pollen Typhae. Thromb. Res. 1990, 57, 957–966. [Google Scholar] [CrossRef]
- Yasukawa, K.; Akihisa, T.; Kanno, H.; Kaminaga, T.; Izumida, M.; Sakoh, T.; Tamura, T.; Takido, M. Inhibitory effects of sterols isolated from Chlorella vulgaris on 12-otetradecanoylphorbol-13-acetate-induced inflammation and tumor promotion in mouse skin. Biol. Pharm. Bull. 1996, 19, 573–576. [Google Scholar] [CrossRef]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyana, M.; Shinmoto, H. Ergosterol peroxide from an edible mushroom suppresses inflammatory responses in RAW264.7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharmacol. 2007, 150, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Lin, P.Y.; Tsai, C.T.; Chuang, W.L.; Chao, Y.H.; Pan, I.H.; Chen, Y.K.; Lin, C.C.; Wang, B.Y. Chlorella sorokiniana induces mitochondrial-mediated apoptosis in human non-small cell lung cancer cells and inhibits xenograft tumor growth in vivo. BMC Complement. Altern. Med. 2017, 17, 88. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, M.L.S.; Bincoletto, C.; Valadares, M.C.; Dantas, D.C.; Santos, L.M. Effects of Chlorella vulgaris extract on cytokines production in Listeria monocytogenes infected mice. Immunopharmacol. Immunotoxicol. 2002, 24, 483–496. [Google Scholar] [CrossRef] [PubMed]
- De Souza Queiroz, J.; Torello, C.O.; Palermo-Neto, J.; Valadares, M.C.; Queiroz, M.L.S. Hematopoietic response of rats exposed to the impact of an acute psychophysiological stressor on responsiveness to an in vivo challenge with Listeria monocytogenes: Modulation by Chlorella vulgaris prophylactic treatment. Brain Behav. Immun. 2008, 22, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, M.L.S.; da Rocha, M.C.; Torello, C.O.; de Souza Queiroz, J.; Bincoletto, C.; Morgano, M.A.; Romano, M.R.; Paredes-Gamero, E.J.; Barbosa, C.M.; Calgarotto, A.K. Chlorella vulgaris restores bone marrow cellularity and cytokine production in lead-exposed mice. Food Chem. Toxicol. 2011, 49, 2934–2941. [Google Scholar] [CrossRef] [PubMed]
- Konishi, F.; Tanaka, K.; Himeno, K.; Taniguchi, K.; Nomoto, K. Antitumor effect induced by a hot water extract of Chlorella vulgaris (CE): Resistance to Meth-A tumor growth mediated by CE-induced polymorphonuclear leukocytes. Cancer Immunol. Immunother. 1985, 19, 73–78. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Changes occurring in immune responsiveness of single- and twin bearing Comisana ewes during the transition period. J. Dairy Sci. 2006, 89, 562–568. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Annicchiarico, G.; Albenzio, M.; Muscio, A.; Sevi, A. Hypothalamic-pituitary-adrenal axis activation and immune regulation in heat-stressed sheep after supplementation with polyunsaturated fatty acids. J. Dairy Sci. 2014, 97, 4247–4258. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Mattmiller, S.A.; Raphael, W.; Gandy, G.C.; Sordillo, L.M. Enhanced n-3 phospholipid content reduces inflammatory responses in bovine endothelial cells. J. Dairy Sci. 2012, 95, 7137–7150. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Ciliberti, M.G.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Dietary polyunsaturated fatty acids from flaxseed affect immune responses of dairy sheep around parturition. Vet. Immunol. Immunopathol. 2015, 168, 56–60. [Google Scholar] [CrossRef]
- Galvao, K.N.; Felippe, M.J.B.; Brittin, S.B.; Sper, R.; Fraga, M.; Galvao, J.S.; Caixeta, L.; Guard, C.L.; Ricci, A.; Gilbert, R.O. Evaluation of cytokine expression by blood monocytes of lactating Holstein cows with or without postpartum uterine disease. Theriogenology 2012, 77, 356–372. [Google Scholar] [CrossRef]
- Souza, C.; Teixeira, A.S.A.; Biondo, L.A.; Silveir, L.S.; Calder, P.C.; Neto, J.C.R. Palmitoleic acid reduces the inflammation in LPS-stimulated macrophages by inhibition of NFκB, independently of PPARs. Clin. Exp. Pharmacol. Physiol. 2017, 44, 566–575. [Google Scholar] [CrossRef]
- Spaner, D.E.; Lee, E.; Shi, Y.; Wen, F.; Li, Y.; Tung, S.; McCaw, L.; Wong, K.; Gary-Gouy, H.; Dalloul, A.; et al. PPAR-alpha is a therapeutic target for chronic lymphocytic leukemia. Leukemia 2013, 27, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Fraile, L.; Crisci, E.; Córdoba, L.; Navarro, M.A.; Osada, J.; Montoya, M. Modulatory properties of beta-sytosterol in pig immune responses. Int. Immunopharmacol. 2012, 13, 316–321. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciliberti, M.G.; Albenzio, M.; Francavilla, M.; Neglia, G.; Esposito, L.; Caroprese, M. Extracts from Microalga Chlorella sorokiniana Exert an Anti-Proliferative Effect and Modulate Cytokines in Sheep Peripheral Blood Mononuclear Cells. Animals 2019, 9, 45. https://doi.org/10.3390/ani9020045
Ciliberti MG, Albenzio M, Francavilla M, Neglia G, Esposito L, Caroprese M. Extracts from Microalga Chlorella sorokiniana Exert an Anti-Proliferative Effect and Modulate Cytokines in Sheep Peripheral Blood Mononuclear Cells. Animals. 2019; 9(2):45. https://doi.org/10.3390/ani9020045
Chicago/Turabian StyleCiliberti, Maria Giovanna, Marzia Albenzio, Matteo Francavilla, Gianluca Neglia, Luigi Esposito, and Mariangela Caroprese. 2019. "Extracts from Microalga Chlorella sorokiniana Exert an Anti-Proliferative Effect and Modulate Cytokines in Sheep Peripheral Blood Mononuclear Cells" Animals 9, no. 2: 45. https://doi.org/10.3390/ani9020045