Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
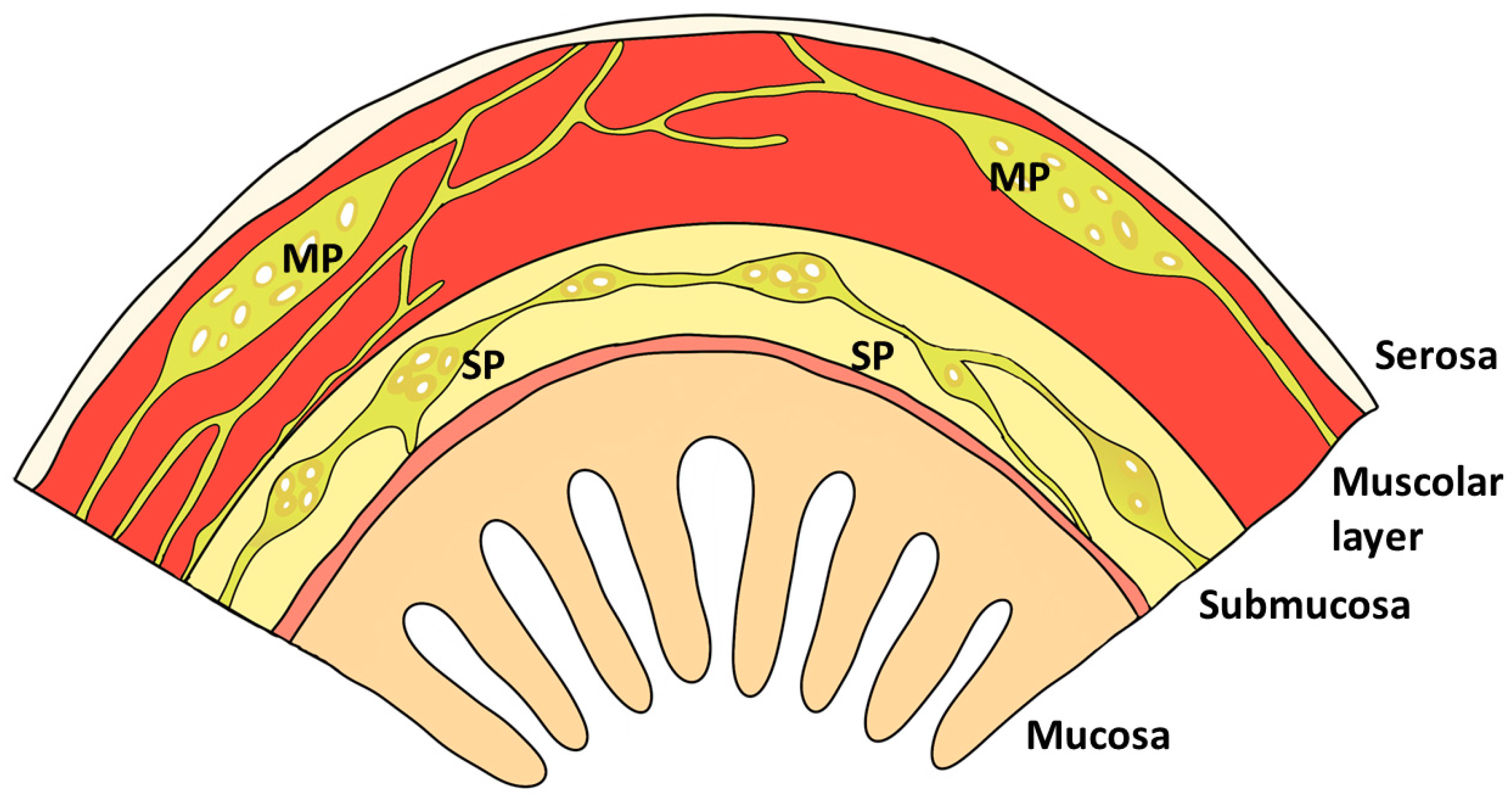
2. Gastrointestinal Barrier
2.1. Epithelial Cells
2.2. Immune System
2.3. Enteric Nervous System
3. GIT Barrier Disorders at Weaning
4. Additives
4.1. Functional Amino Acids
4.2. Phytochemicals
4.3. Organic Acids
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Worobec, E.K.; Duncan, I.J.H.; Widowski, T.M. The effects of weaning at 7, 14 and 28 days on piglet behavior. Appl. Anim. Behav. Sci. 1999, 62, 173–182. [Google Scholar] [CrossRef]
- Wiese, F.; Simon, O.; Weyrauch, K.D. Morphology of the small intestine of weaned piglets and a novel method for morphometric evaluation. Anat. Histol. Embryol. 2003, 32, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P. Role of intestinal epithelial cells in the innate immune defence of the pig intestine. Vet. Res. 2006, 37, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Edelblum, K.L.; Turner, J.R. The tight junction in inflammatory disease: Communication breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. 2010, 5, 119–144. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Turner, J.R.; Rill, B.K.; Carlson, S.L.; Carnes, D.; Kerner, R.; Mrsny, R.J. Physiological regulation of epithelial tight junctions is associated with myosin light-chain phosphorylation. Am. J. Physiol. 1997, 273, C1378–C1385. [Google Scholar] [CrossRef]
- Moeser, A.J.; Blikslager, A.T. Mechanisms of porcine diarrheal disease. J. Am. Vet. Med. Assoc. 2007, 231, 56–67. [Google Scholar] [CrossRef]
- Moeser, A.J.; Klok, C.V.; Ryan, K.A.; Wooten, J.G.; Little, D.; Cook, V.L. Stress signaling pathways activated by weaning mediate intestinal dysfunction in the pig. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G173–G181. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Sakar, Y.; Covasa, M. The modulatory role of high fat feeding on gastrointestinal signals in obesity. J. Nutr. Biochem. 2013, 24, 1663–1677. [Google Scholar] [CrossRef] [PubMed]
- Little, T.J.; Feinle-Bisset, C. Effects of dietary fat on appetite and energy intake in health and obesity–oral and gastrointestinal sensory contributions. Physiol. Behav. 2011, 104, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Mazzoni, M.; De Giorgio, R.; Latorre, R.; Vallorani, C.; Bosi, P.; Trevisi, P.; Barbara, G.; Stanghellini, V.; Corinaldesi, R.; Forni, M.; et al. Expression and regulation of α-transducin in the pig gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 466–474. [Google Scholar] [CrossRef]
- Rossi, R.; Corino, C.; Modina, S.; Di Giancamillo, A. Dietary Verbascoside Influences Gut Morphology and the expression of α-transducin and α-gustducin in the small intestine of weaned piglets exposed to n-6 polyunsaturated fatty acids-induced oxidative stress. Animals 2019, 9, 20. [Google Scholar] [CrossRef]
- Hershberg, R.; Blumberg, R.S. The lymphocyte-epithelial-bacterial interface. In Inflammatory Bowel Disease: From Bench to Bedside; Targan, S.R., Shanahan, F., Karp, L.C., Eds.; Springer: Boston, MA, USA, 2005; pp. 121–146. [Google Scholar]
- Pabst, R. The anatomical basis for the immune function of the gut. Anat. Embriol. 1987, 176, 135–144. [Google Scholar] [CrossRef]
- Bensussanm, C.N.; Guy-Grand, D. Intestinal intraepithelial lymphocytes. Gastroenterol. Clin. N. Am. 1991, 20, 549–576. [Google Scholar]
- McGhee, J.R.; Mestecky, J.; Dertzbaugh, M.T.; Eldrige, J.H.; Hirasawa, M.; Kiyono, H. The mucosal immune system: From fundamental concepts to vaccine development. Vaccine 1992, 10, 75–88. [Google Scholar] [CrossRef]
- Vega-Lopez, M.A.; Telemo, E.; Bailey, M.; Stevens, K.; Stokes, C.R. Immune cell distribution in the small intestine of the pig: Immunohistological evidence for an organised compartmentalisation in the lamina propria. Vet. Immunol. Immunopathol. 1993, 37, 49–60. [Google Scholar] [CrossRef]
- Pabst, R.; Rothkotter, H.J. Postnatal development of lymphocyte subsets in different compartments of the small intestine of piglets. Vet. Immunol. Immunopathol. 1999, 72, 167–173. [Google Scholar] [CrossRef]
- Vega-Lopez, M.A.; Bailey, M.; Telemo, E.; Stokes, C.R. Effect of early weaning on the development of immune cells in the pig small intestine. Vet. Immunol. Immunopathol. 1995, 44, 319–327. [Google Scholar] [CrossRef]
- Van Wijk, F.; Cheroutre, H. Intestinal T cells: Facing the mucosal immune dilemma with synergy and diversity. Semin. Immunol. 2009, 21, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Potten, C.S. Kinetics and possible regulation of crypt cell populations under normal and stress conditions. Bull. Cancer 1975, 62, 419–430. [Google Scholar] [PubMed]
- Barker, N. Adult intestinal stem cells: Critical drivers of epithelial homeostasis and regeneration. Nat. Rev. Mol. Cell Biol. 2014, 15, 19–33. [Google Scholar] [CrossRef]
- Qi, Z.; Chen, Y.G. Regulation of intestinal stem cell fate specification. Sci. China Life Sci. 2015, 58, 570–578. [Google Scholar] [CrossRef]
- Kraehenbuhl, J.P.; Neutra, M.R. Molecular and cellular basis of immune protection of mucosal surfaces. Physiol. Rev. 1992, 72, 853–879. [Google Scholar] [CrossRef]
- Neutra, M.R.; Frey, A.; Kraehenbuhl, J.P. Epithelial M cells: Gateways for mucosal infection and immunization. Cell 1996, 86, 345–348. [Google Scholar] [CrossRef]
- Neutra, M.R.; Pringault, E.; Kraehenbuhl, J.P. Antigen sampling across epithelial barriers and induction of mucosal immune responses. Annu. Rev. Immunol. 1996, 14, 275–300. [Google Scholar] [CrossRef]
- Jensen, V.B.; Harty, J.T.; Jones, B.D. Interactions of the invasive pathogens Salmonella typhimurium, Listeria monocytogenes, and Shigella flexneri with M cells and murine Peyer’s patches. Infect. Immun. 1998, 66, 3758–3766. [Google Scholar]
- Kerneis, S.; Bogdanova, A.; Kraehenbuhl, J.P.; Pringault, E. Conversion by Peyer’s patches lymphocytes of human enterocytes into M cells that transport bacteria. Science 1997, 277, 949–952. [Google Scholar] [CrossRef]
- Gebert, A.; Posselt, W. Glycoconjugate expression defines the origin and differentiation pathway of intestinal M cells. J. Histochem. Cytochem. 1997, 45, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Palus, K.; Makowska, K.; Całka, J. Acrylamide-induced alterations in the cocaine- and amphetamine-regulated peptide transcript (CART)-like immunoreactivity within the enteric nervous system of the porcine small intestines. Ann. Anat. 2018, 219, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, S.; Cailotto, C.; Harthoorn, L.F.; De Jonge, W.J. Cholinergic signalling in gut immunity. Life Sci. 2012, 91, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cabezudo, M.J.; Lorke, D.E.; Azimullah, S.; Mechkarska, M.; Hasan, M.Y.; Petroianu, G.A. Cholinergic stimulation of the immune system protects against lethal infection by Salmonella enterica serovar typhimurium. Immunology 2010, 30, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Price, L.D. Catecholamines and sympathomimetic drugs decrease early Salmonella Typhimurium uptake into porcine Peyer’s patches. FEMS Immunol. Med. Microbiol. 2008, 52, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Lyte, M.; Stevens, M.P.; Vulchanova, L.; Brown, D.R. Mucosally-directed adrenergic nerves and sympathomimetic drugs enhance non-intimate adherence of Escherichia coli O157:H7 to porcine cecum and colon. Eur. J. Pharmacol. 2006, 539, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, K.L.; Brown, D.R. Adrenocorticotrophic hormone modulates Escherichia coli O157:H7 adherence to porcine colonic mucosa. Stress 2005, 8, 185–190. [Google Scholar] [CrossRef]
- Medland, J.E.; Pohl, C.S.; Edwards, L.L.; Frandsen, S.; Bagley, K.; Li, Y. Early life adversity in piglets induces long-term up regulation of the enteric cholinergic nervous system and heightened, sex-specific secretomotor neuron responses. Neurogastroenterol. Motil. 2016, 28, 1317–1329. [Google Scholar] [CrossRef] [Green Version]
- Pohl, C.S.; Medland, J.E.; Mackey, E.; Edwards, L.L.; Bagley, K.D.; De Wilde, M.P.; Williams, K.J.; Moeser, A.J. Early weaning stress induces chronic functional diarrhea, intestinal barrier defects, and increased mast cell activity in a porcine model of early life adversity. Neurogastroenterol. Motil. 2017, 29, e13118. [Google Scholar] [CrossRef]
- Wijtten, P.J.; Van der Meulen, J.; Verstegen, M.W. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef]
- Moeser, A.J.; Calvin, S.P.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Ryan, K.A.; Nighot, P.K.; Blikslager, A.T. Gastrointestinal dysfunction induced by early weaning is attenuated by delayed weaning and mast cell blockade in pigs. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G413–G421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.; Clark, J.E.; Overman, B.L.; Tozel, C.C.; Huang, J.H.; Rivier, J.E.F.; Blisklager, A.T.; Moeser, A.J. Early weaning stress impairs development of mucosal barrier function in the porcine intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G352–G363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, D.; Smyth, J.A.; MC Cracken, K.J. Digestive development of the early-weaned pig. Effect of level of food intake on digestive enzyme activity during the immediate post-weaning period. Br. J. Nutr. 1991, 65, 181–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Beers-Schreurs, H.M.G.; Nabuurs, M.J.A.; Vel-lenga, L.; Kalsbeck-van der Valk, H.J.; Wensing, T.; Breukin, H.J. Weaning and the weanling diet influence the villous height and crypt depth in the small intestine of pigs and alter the concentrations of short-chain fatty acids in the large intestine and blood. J. Nutr. 1998, 128, 947–953. [Google Scholar] [CrossRef]
- Hedemann, M.S.; Hojsgaard, S.; Jensen, B.B. Small intestinal morphology and activity of intestinal peptidases in piglets around weaning. J. Anim. Physiol. Anim. Nutr. 2003, 87, 32–41. [Google Scholar] [CrossRef]
- Vente-Spreeuwenberg, M.A.M.; Vedonk, J.M.A.J.; Verstegen, M.W.A.; Beyen, A.C. Villus height and gut development in weaned piglets receiving diets containing either glucose, lactose or starch. Br. J. Nutr. 2003, 90, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xiong, X.; Wang, X.; Li, T.; Yin, Y. Effects of weaning on intestinal crypt epithelial cells in piglets. Sci. Rep. 2016, 6, 36939. [Google Scholar] [CrossRef] [Green Version]
- Pie, S.; Lalles, J.P.; Blazy, F.; Laffitte, J.; Seve, B.; Oswald, I.P. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Mc Cracken, B.A.; Spurlock, M.E.; Roos, M.A.; Zuckermann, F.A.; Gaskins, H.R. Weaning anorexia may contribute to local inflammation in the piglet small intestine. J. Nutr. 1999, 129, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domeneghini, C.; Di Giancamillo, A.; Arrighi, S.; Bosi, G. Gut-trophic feed additives and their effects upon the gut structure and intestinal metabolism. State of the art in the pig, and perspectives towards humans. Histol. Histopathol. 2006, 21, 273–283. [Google Scholar] [PubMed]
- Lallès, J.P.; Bosi, P.; Janczyk, P.; Koopmans, S.J.; Torrallardona, D. Impact of bioactive substances on the gastrointestinal tract and performance of weaned piglets: A review. Animal 2009, 3, 1625–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Espinosab, C.D.; Abelillab, J.J.; Casasb, G.A.; Lagos, L.V.; Lee, S.A.; Kwon, W.B.; Mathai, J.K.; Navarro, D.M.; Jaworski, N.W.; et al. Non-antibiotic feed additives in diets for pigs: a review. Anim. Nutr. 2018, 4, 113–125. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Hanczakowska, E.; Niwińska, B. Glutamine as a feed supplement for piglets: A review. Ann. Anim. Sci. 2013, 13, 5–15. [Google Scholar] [CrossRef]
- He, J.; Feng, G.D.; Ao, X.; Li, Y.; Qian, H.X.; Liu, J.B.; He, Z.Z. Effects of L-glutamine on growth performance, antioxidant ability, immunity and expression of genes related to intestinal health in weanling pigs. Livest. Sci. 2016, 189, 102–109. [Google Scholar] [CrossRef]
- Achamrah, N.; Déchelotte, P.; Coëffier, M. Glutamine and the regulation of intestinal permeability: From bench to bedside. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 86–91. [Google Scholar] [CrossRef]
- Rose, W.C.; Oesterling, M.J.; Womack, M. Comparative growth on diets containing ten and 19 amino acids, with further observations upon the role of glutamic and aspartic acids. J. Biol. Chem. 1948, 176, 753–762. [Google Scholar]
- Rose, W.C.; Smith, L.C.; Womack, M.; Shane, M.J. The utilization of the nitrogen of ammonium salts, urea, and certain other compounds in the synthesis of non-essential amino acid in vivo. J. Biol. Chem. 1949, 181, 307–316. [Google Scholar]
- Watford, M. Glutamine and glutamate: Nonessential or essential amino acids? Anim. Nutr. 2015, 1, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Turpin, D.L.; Kim, J.C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Meier, S.A.; Knabe, D.A. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J. Nutr. 1996, 126, 2578–2584. [Google Scholar] [CrossRef] [PubMed]
- Ewtushick, A.L.; Bertolo, R.F.P.; Ball, R.O. Intestinal development of early-weaned piglets receiving diets supplemented with selected amino acids or polyamines. Can. J. Anim. Sci. 2000, 80, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Domeneghini, C.; Di Giancamillo, A.; Savoini, G.; Paratte, R.; Bontempo, V.; Dell’Orto, V. Structural patterns of swine ileal mucosa following L-glutamine and nucleotide administration during the weaning period. An histochemical and histometrical study. Histol. Histopathol. 2004, 19, 49–58. [Google Scholar]
- Wang, H.; Zhang, C.; Wu, G.; Sun, Y.; Wang, B.; He, B.; Dai, Z.; Wu, Z. Glutamine enhances tight junction protein expression and modulates corticotropin-releasing factor signaling in the jejunum of weanling piglets. J. Nutr. 2015, 45, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Fürst, P.; Pogan, K.; Stehle, P. Glutamine dipeptides in clinical nutrition. Nutrition 1997, 13, 731–737. [Google Scholar] [CrossRef]
- Miguel, J.C.; Maxwell, A.A.; Hsieh, J.J.; Harnisch, L.C.; Alam, D.A.; Polk, B.; Lien, C.L.; Watson, A.J.M.; Frey, M.R. Epidermal growth factor suppresses intestinal epithelial cell shedding through a MAPK-dependent pathway. J. Cell Sci. 2017, 130, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Zhang, B.; Lin, M.; Zhou, P.; Li, J.; Zhang, L.; Gao, F.; Zhou, G. Effects of alanyl-glutamine supplementation on the small intestinal mucosa barrier in weaned piglets Asian-Australas. J. Anim. Sci. 2017, 30, 236–245. [Google Scholar]
- Lopez-Pedrosa, J.M.; Torres, M.I.; Fernandez, M.I.; Rios, A.; Gill, A. Severe malnutrition alters lipid composition and fatty acid profile on the small intestine in newborn piglets. J. Nutr. 1998, 128, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.C.; Maxwell, C.V.; Erf, G.F.; Davis, M.E.; Singh, S.; Johnson, Z.B. The influence of different management systems and age on intestinal morphology, immune cell numbers and mucin production from goblet cells in post-weaning pigs. Vet. Immunol. Immunopathol. 2006, 111, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, M.C.; Bueno, J.D.; Ayudarte, M.V.; Allmendros, A.; Rios, A.; Suarez, M.D.; Gil, A. Dietary restriction induces biochemical and morphological changes in the small intestine of nursing piglets. J. Nutr. 1996, 126, 933–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Knabe, D.A.; Kim, S.W. Arginine nutrition in neonatal pigs. J. Nutr. 2004, 134, 2783S–2790S. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Tang, H.; Ren, P.; Kong, X.; Wu, G.; Yin, Y.; Wang, Y. Dietary Supplementation with L-Arginine partially counteracts serum metabolome induced by weaning stress in piglets. J. Proteome Res. 2011, 10, 5214–5221. [Google Scholar] [CrossRef] [PubMed]
- Tong, B.C.; Barbul, A. Cellular and physiological effects of arginine. Mini Rev. Med. Chem. 2004, 4, 823–832. [Google Scholar] [CrossRef]
- Kim, S.W.; Wu, G. Dietary arginine supplementation enhances the growth of milk-fed young pigs. J. Nutr. 2004, 134, 625–630. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Jaeger, L.A.; Johnson, G.A.; Sung Woo Kim, S.W.; Knabe, D.A.; Meininger, C.J.; Spencer, T.E.; Yin, Y.L. Important roles for the arginine family of amino acids in swine nutrition and production. Livest. Sci. 2007, 112, 8–22. [Google Scholar] [CrossRef]
- Cynober, L. Can arginine and ornithine support gut functions? Gut 1994, 35, S42–S45. [Google Scholar] [CrossRef]
- Tan, B.; Xiao, H.; Xiong, X.; Wang, J.; Li, G.; Yin, Y.; Huang, B.; Hou, Y.; Wu, G. L-arginine improves DNA synthesis in LPS-challenged enterocytes. Front. Biosci. 2015, 1, 989–1003. [Google Scholar] [CrossRef]
- Shan, Y.; Shan, A.; Li, J.; Zhou, C. Dietary supplementation of arginine and glutamine enhances the growth and intestinal mucosa development of weaned piglets. Livest. Sci. 2012, 150, 369–373. [Google Scholar] [CrossRef]
- Hampson, D.J.; Kidder, D.E. Influence of creep feeding and weaning on brush border enzyme activities in the piglet small intestine. Res. Vet. Sci. 1986, 40, 24–31. [Google Scholar] [CrossRef]
- Cao, S.T.; Wang, C.C.; Wu, H.; Zhang, Q.H.; Jiao, L.F.; Hu, C.H. Weaning disrupts intestinal antioxidant status, impairs intestinal barrier and mitochondrial function, and triggers mitophagy in piglets. J. Anim. Sci. 2018, 96, 1073–1108. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Foti, M.C.; Valgimili, L. Antioxidant activity of essential oils. J. Agric. Food Chem. 2013, 61, 10835–10847. [Google Scholar] [CrossRef] [PubMed]
- Frankič, T.; Levart, A.; Salobir, J. The effect of vitamin E and plant extract mixture composed of carvacrol, cinnamaldehyde and capsaicin on oxidative stress induced by high PUFA load in young pigs. Animal 2010, 4, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Che, T.M.; Song, M.; Lee, J.J.; Almeida, J.A.; Bravo, D.; Van Alstine, W.G.; Pettigrew, J.E. Dietary plant extracts improve immune responses and growth efficiency of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. 2013, 91, 5668–5679. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, M.; Che, T.M.; Almeida, J.A.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts alleviate diarrhea and alter immune responses of weaned pigs experimentally infected with a pathogenic Escherichia coli. J. Anim. Sci. 2013, 91, 5294–5306. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Xiang, Q.; Wang, J.; Peng, J.; Wei, H. Oregano essential oil improves intestinal morphology and expression of tight junction proteins associated with modulation of selected intestinal bacteria and immune status in a pig model. Bio Med. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Yuan, D.; Hussain, T.; Tan, B.; Liu, Y.; Ji, P.; Yin, Y. The evaluation of antioxidant and anti-Inflammatory effects of Eucommia ulmoides flavones using diquat-challenged piglet models. Oxidative Med. Cell. Longev. 2017. [Google Scholar] [CrossRef] [Green Version]
- Fiesel, A.; Gessner, D.K.; Most, E.; Ede, K. Effects of dietary polyphenol-rich plant products from grape or hop on pro-inflammatory gene expression in the intestine, nutrient digestibility and faecal microbiota of weaned pigs. BMC Vet. Res. 2014, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Sehm, J.; Lindermayer, H.; Dummer, C.; Treutter, D.; Pfaffl, M.W. The influence of polyphenol rich apple pomace or red-wine pomace diet on the gut morphology in weaning piglets. J. Anim. Physiol. Anim. Nutr. 2007, 91, 289–296. [Google Scholar] [CrossRef]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzanilla, E.G.; Nofrarías, M.; Anguita, M.; Castillo, M.; Pérez, J.; Martín-Orúe, S.M.; Kamel, C.; Gasa, J. Effects of butyrate, avilamycin, and a plant extract combination on the intestinal equilibrium of early-weaned pigs. J. Anim. Sci. 2006, 84, 2743–2751. [Google Scholar] [CrossRef] [PubMed]
- Nofrarias, M.; Manzanilla, E.G.; Pujols, J.; Gibert, X.; Majo, N.; Segales, J.; Gasa, J. Effects of spray-dried porcine plasma and plant extracts on intestinal morphology and on leukocyte cell subsets of weaned pigs. J. Anim. Sci. 2006, 84, 2735–2742. [Google Scholar] [CrossRef] [PubMed]
- Di Giancamillo, A.; Rossi, R.; Vitari, F.; Carollo, V.; Deponti, D.; Corino, C.; Domeneghini, C. Evidence of nitrosative stress biomarkers changes in the swine intestine following dietary intervention with verbascoside. Histol. Histopathol. 2013, 28, 715–723. [Google Scholar] [PubMed]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef]
- Vigan, M. Essential oils: Renewal of interest and toxicity. Eur. J. Dermatol. 2010, 20, 685–692. [Google Scholar]
- Partanen, K.H.; Mroz, Z. Organic acids for performance enhancement in pig diets. Nutr. Res. Rev. 1999, 12, 117–145. [Google Scholar] [CrossRef] [Green Version]
- Mroz, Z.; Koopmans, S.J.; Bannink, A.; Partanen, A.K.; Krasucki, W.; Overland, M.; Radcliffe, S. Carboxylic acids as bioregulators and gut growth promoters in non-ruminants. In Biology of Nutrition in Growing Animals; Mosenthin, R., Zentek, J., Zebrowska, T., Eds.; Elsevier Limited: Amsterdam, The Netherlands, 2006; Volume 4, pp. 81–133. [Google Scholar]
- Suiryanrayna, V.M.; Ramana, V.J. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 45. [Google Scholar] [CrossRef] [Green Version]
- Papatsiros, V.; Katsoulos, P.-D.; Koutoulis, K.; Karatzia, M.; Dedousi, A.; Christodoulopoulos, G. Alternatives to antibiotics for farm animals. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2013, 8, 1–15. [Google Scholar] [CrossRef]
- Randy, C.E.; Armstrong, W.D.; Herman, D.L. The development of digestive capacity in young pigs: Effects of age and weaning system. J. Anim. Sci. 1982, 55, 1380–1387. [Google Scholar]
- Cranwell, P.D. The development of acid and pepsin (EC 3. 4. 23. 1) secretory capacity in the pig; the effects of age and weaning: 1. Studies in anaesthetized pigs. Br. J. Nutr. 1985, 54, 305–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, T. Stimulatory effect of short-chain fatty acids on epithelial cell proliferation in the rat intestine: A possible explanation for trophic effects of fermentable fibre, gut microbes and luminal trophic factors. Br. J. Nutr. 1987, 8, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, T.; Adachi, M.; Hashida, M.; Sato, N.; Kojima, T. Effect of n-butyric acid on epithelial cell proliferation of pig colonic mucosa in short-term culture. Dtsch. Tierarztl. Wochenschr. 1995, 102, 163–164. [Google Scholar] [PubMed]
- Gálfi, P.; Bokori, J. Feeding trial in pigs with a diet containing sodium n-butyrate. Acta Vet. Hung. 1990, 38, 3–17. [Google Scholar]
- Blank, R.; Mosenthin, R.; Sauer, W.C.; Huang, S. Effect of fumaric acid and dietary buffering capacity on ileal and fecal amino acid digestibilities in early-weaned pigs. J. Anim. Sci. 1999, 77, 2974–2984. [Google Scholar] [CrossRef] [Green Version]
- Diao, H.; Zheng, P.; Yu, B.; He, J.; Mao, X.B.; Yu, J.; Chen, D.W. Effects of dietary supplementation with benzoic acid on intestinal morphological structure and microflor in weaned piglets. Livest. Sci. 2014, 167, 249–256. [Google Scholar] [CrossRef]
- Halas, D.; Hansen, C.F.; Hampsona, D.J.; Mullan, B.P.; Kimb, J.C.; Wilson, R.H.; Pluske, J.R. Dietary supplementation with benzoic acid improves apparent ileal digestibility of total nitrogen and increases villous height and caecal microbial diversity in weaner pigs. Anim. Feed Sci. Technol. 2010, 160, 137–147. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acidscan improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef]
- Ma, X.; Fan, P.X.; Li, L.S.; Qiao, S.Y.; Zhang, G.L.; Li, D.F. Butyrate promotes the recovering of intestinal wound healing through its positive effect on the tight junctions. Anim. Sci. 2012, 90, 266–268. [Google Scholar] [CrossRef] [Green Version]
- Grilli, E.; Tugnoli, B.; Foerster, C.J.; Piva, A. Butyrate modulates inflammatory cytokines and tight junctions components along the gut of weaned pigs. J. Anim. Sci. 2016, 94, 433–436. [Google Scholar] [CrossRef]
- Le Gall, M.; Gallois, M.; Seve, B.; Louveau, I.; Holst, J.J.; Oswald, I.P.; Lalles, J.; Guilloteau, P. Comparative effect of orally administered sodium butyrate before or after weaning on growth and several indices of gastrointestinal biology of piglets. Br. J. Nutr. 2009, 102, 1285–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagi, G.; Piva, A.; Moschini, M.; Vezzali, E.; Roth, F.X. Performance, intestinal microflora, and wall morphology of weanling pigs fed sodium butyrate. J. Anim. Sci. 2007, 85, 1184–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrara, F.; Tedin, L.; Pieper, R.; Meyer, W.; Zentek, J. Influence of medium-chain fatty acids and short-chain organic acids on jejunal morphology and intra-epithelial immune cells in weaned piglets. J. Anim. Physiol. Anim. Nutr. 2016, 101, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Grecco, H.A.T.; Amorim, A.B.; Saleh, M.A.D.; Tse, M.L.P.; Telles, F.G.; Miassi, G.M.; Pimenta, G.M.; Berto, D.A. Evaluation of growth performance and gastro-intestinal parameters on the response of weaned piglets to dietary organic acids. An. Acad. Bras. Cienc. 2018, 90, 401–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Morphology | Enterocyte Proliferation | Goblet Cells | Epithelial Junctions | ||||
|---|---|---|---|---|---|---|---|
| Effect | Villi | Crypts | Villi:crypt Ratio | ||||
| Amino Acids | Positive | 53, 64–67, 69, 79, 81 | 69, 79, 81 | 67, 81 | 53, 66, 70, 80 | 89 | 69,69 |
| Null | 67 | 69 | |||||
| Negative | |||||||
| Phytochemicals | Positive | 88, 89 | 89, 91 | 88 | |||
| Null | 87, 90, 95 | 87, 90, 95 | 90, 95 | ||||
| Negative | 92 | ||||||
| Organic Acids | Positive | 107, 108, 109, 110 | 107 | 108, 110 | 106, 110 | 111, 112 | |
| Null | 114, 115 | 116 | 115 | ||||
| Negative | 113, 116 | 110 | 110, 112 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modina, S.C.; Polito, U.; Rossi, R.; Corino, C.; Di Giancamillo, A. Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets. Animals 2019, 9, 1045. https://doi.org/10.3390/ani9121045
Modina SC, Polito U, Rossi R, Corino C, Di Giancamillo A. Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets. Animals. 2019; 9(12):1045. https://doi.org/10.3390/ani9121045
Chicago/Turabian StyleModina, Silvia Clotilde, Umberto Polito, Raffaella Rossi, Carlo Corino, and Alessia Di Giancamillo. 2019. "Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets" Animals 9, no. 12: 1045. https://doi.org/10.3390/ani9121045