Effects of Partial Replacment of Dietary Forage Using Kelp Powder (Thallus laminariae) on Ruminal Fermentation and Lactation Performances of Dairy Cows

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Kelp Powder
2.2. Animals and Experimental Design
2.3. Feeding Management
2.4. Sampling and Parameters Measurement
2.5. DNA Extraction and Sequencing Process
2.6. Statistical Analysis
3. Results
3.1. Animal Production Performance and Rumen Fermentation Parameters
3.2. Sequencing Information
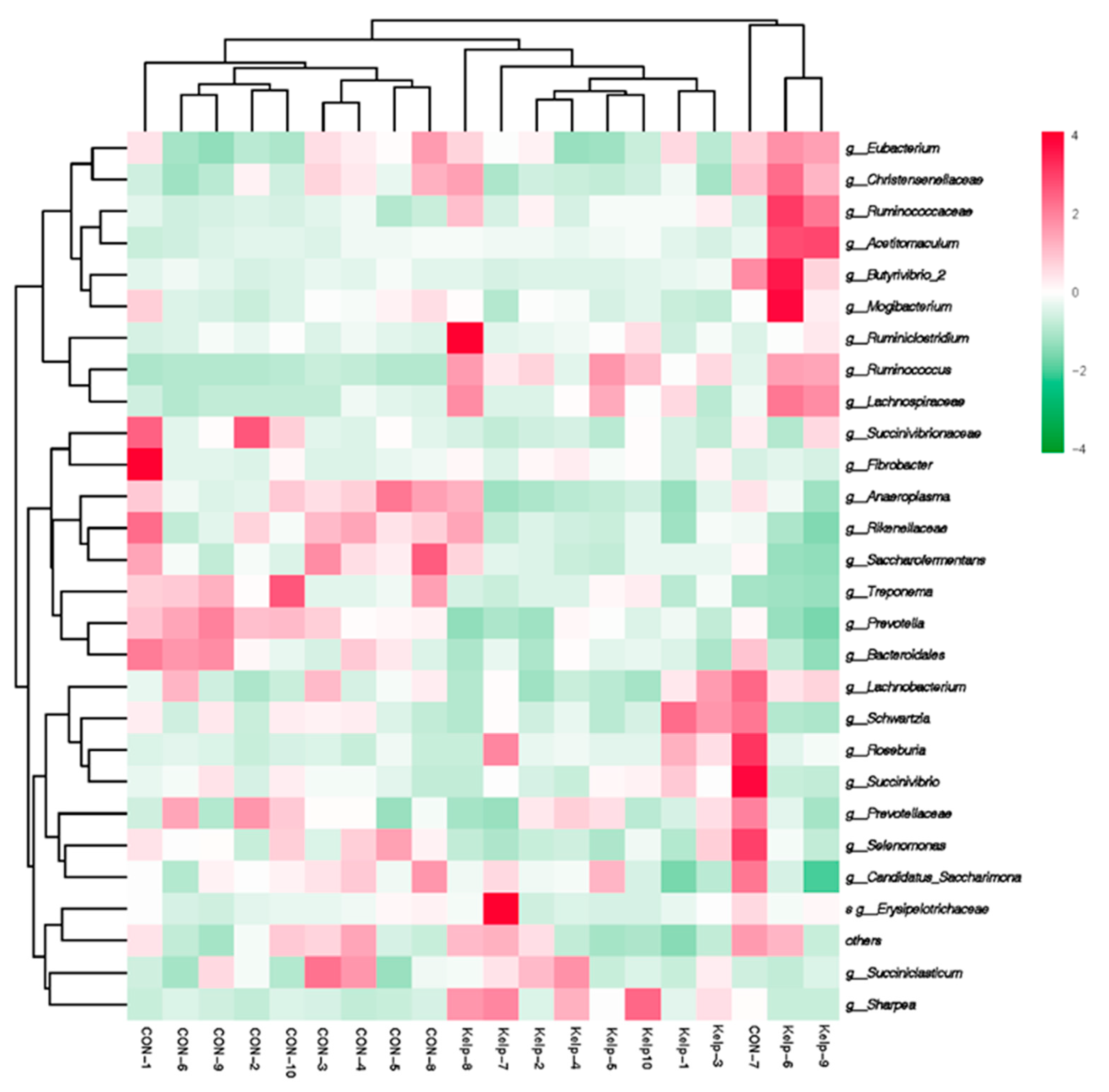
3.3. Effects of Kelp Replacing Diet on Ruminal Bacteria
3.3.1. α-Diversity
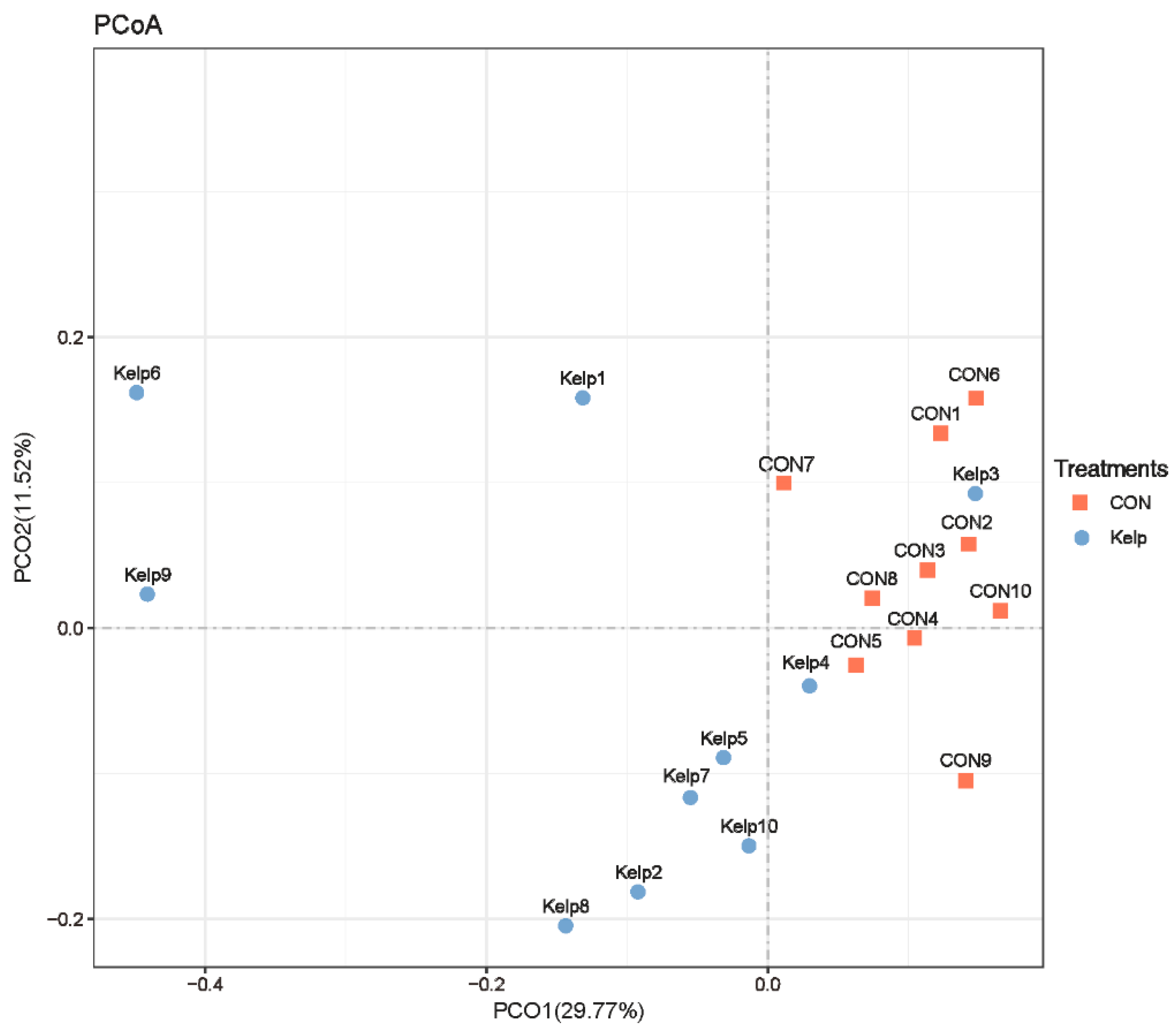
3.3.2. β-Diversity
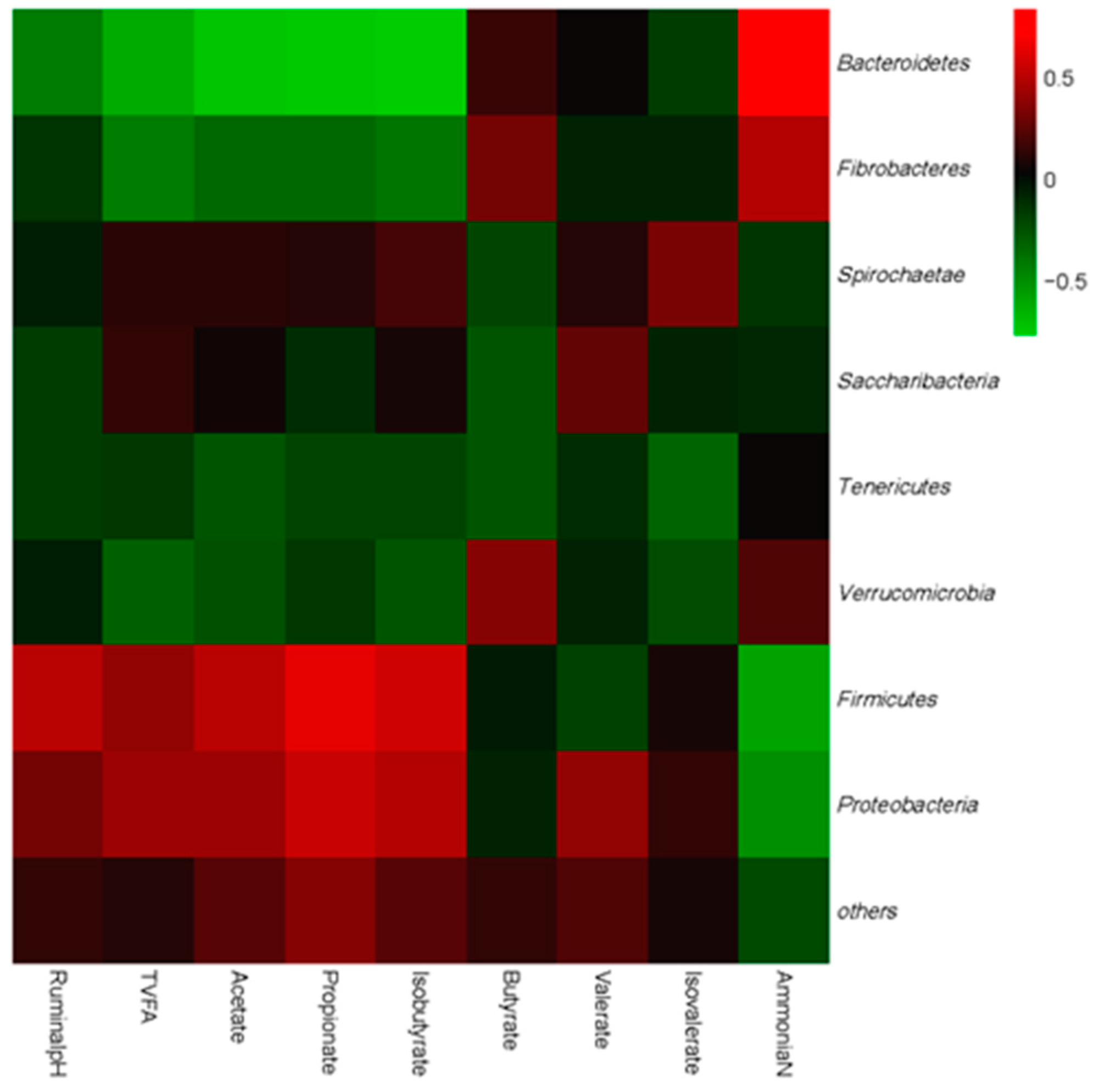
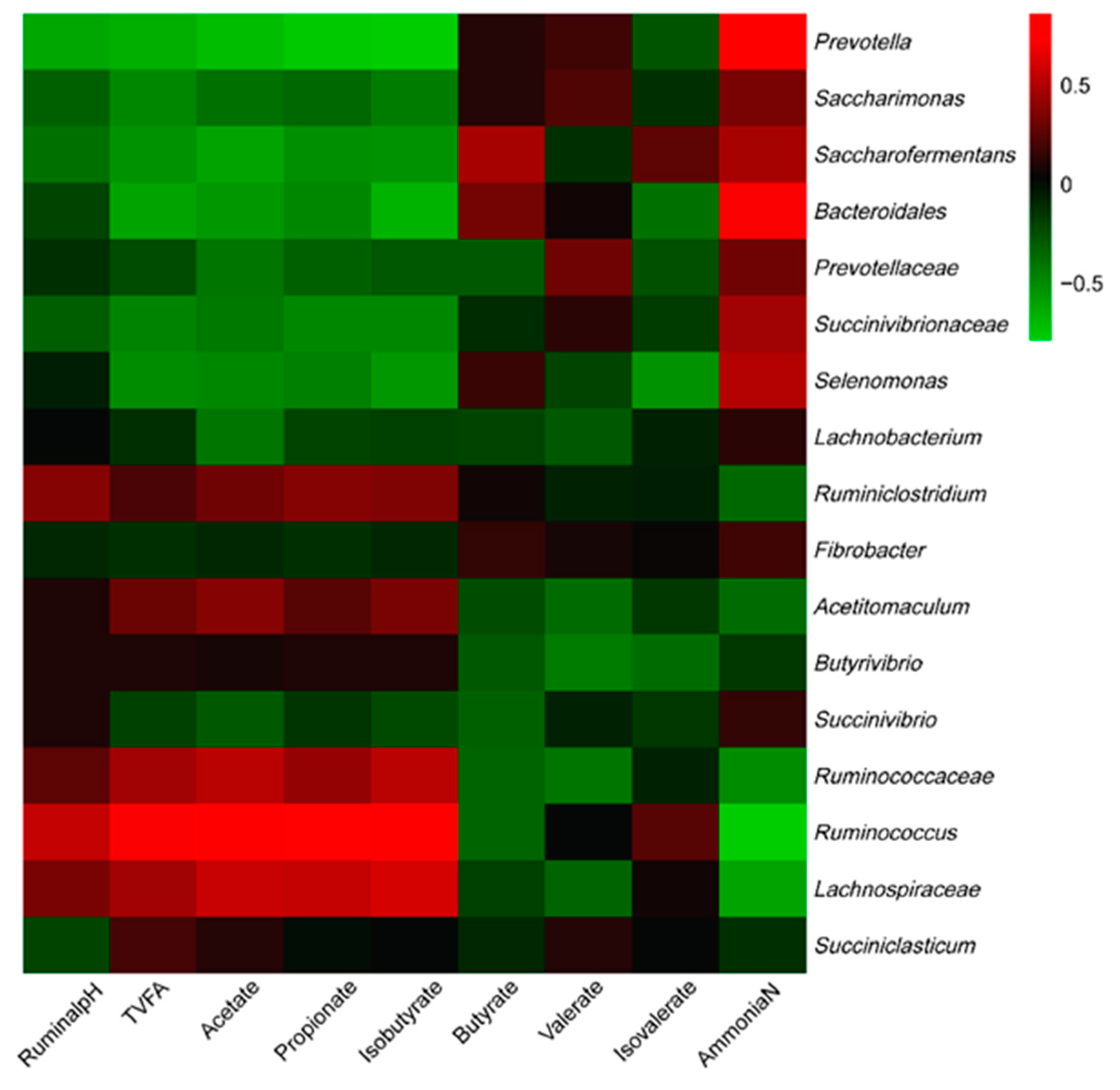
3.4. Correlations between Bacteria Communities and Ruminal Fermentation Parameters
4. Discussion
4.1. Effects of Kelp Feeding on Ruminal Carbohydrate Metabolism
4.2. Effects of Kelp Feeding on Ruminal Nitrogen Metabolism
4.3. Effect of Kelp Feeding on Milk Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Overton, T.R.; Yasui, T. Practical applications of trace minerals for dairy cattle. J. Anim. Sci. 2014, 92, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.L.; Browning, J.F.; Burdette, C.Q.; Caceres, G.C.; Chieh, K.D.; Davis, W.C.; Kassim, B.L.; Long, S.E.; Murphy, K.E.; Oflaz, R.; et al. Development of a kelp powder (Thallus laminariae) Standard Reference Material. Anal. Bioanal. Chem. 2018, 410, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.M.; Okab, A.B.; Aljumaah, R.S.; Samara, E.M.; Abdoun, K.A.; Alhaidary, A.A. Nutritional Value of Green Seaweed (Ulva Lactuca) for Broiler Chickens. Ital. J. Anim. Sci. 2013, 12, 612–620. [Google Scholar] [CrossRef]
- Pereira, M.; Tala, F.; Fernandez, M.; Subida, M.D. Effects of kelp phenolic compounds on the feeding-associated mobility of the herbivore snail Tegula tridentata. Mar. Environ. Res. 2015, 112, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Garima, K.; Bruce, R.; Glenn, S.; Nikhil, T.; Franklin, E.; Alan, C.; Jeff, H.; Balakrishnan, P. Feed supplementation with red seaweeds, Chondrus crispus and Sarcodiotheca gaudichaudii, affects performance, egg quality, and gut microbiota of layer hens. Poult. Sci. 2014, 93, 2991–3001. [Google Scholar]
- Heins, B.J.; Chester-Jones, H. Effect of feeding kelp on growth and profitability of group-fed calves in an organic production system. Prof. Ani. Sci. 2015, 31, 368–374. [Google Scholar] [CrossRef]
- Antaya, N.T.; Soder, K.J.; Kraft, J.; Whitehouse, N.L.; Guindon, N.E.; Erickson, P.S.; Conroy, A.B.; Brito, A.F. Incremental amounts of Ascophyllum nodosum meal do not improve animal performance but do increase milk iodine output in early lactation dairy cows fed high-forage diets. J. Dairy Sci. 2015, 98, 1991–2004. [Google Scholar] [CrossRef] [Green Version]
- Pitta, D.W.; Indugu, N.; Kumar, S.; Vecchiarelli, B.; Sinha, R.; Baker, L.D.; Bhukya, B.; Ferguson, J.D. Metagenomic assessment of the functional potential of the rumen microbiome in Holstein dairy cows. Anaerobe 2016, 38, 50–60. [Google Scholar] [CrossRef]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T. Metagenomic Discovery of Biomass-Degrading Genes and Genomes from Cow Rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Puniya, A.K.; Singh, R.; Kamra, D.N. Rumen Microbiology: From Evolution to Revolution; Springer: New Delhi, India, 2015. [Google Scholar]
- An, D.; Cai, S.; Dong, X. Actinomyces ruminicola sp. nov., isolated from cattle rumen. Int. J. Syst. Evol. Microbiol. 2006, 56, 2043–2048. [Google Scholar] [CrossRef]
- Hall, M.B.; Larson, C.C.; Wilcox, C.J. Carbohydrate source and protein degradability alter lactation, ruminal, and blood measures. J. Dairy Sci. 2010, 93, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bai, H.; Zheng, L.; Jiang, H.; Cui, H.; Cao, Y.; Yao, J. Bioactive polysaccharides and oligosaccharides as possible feed additives to manipulate rumen fermentation in rusitec fermenters. Int. J. Biol. Macromol. 2018, 109, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Vyas, D.; Beauchemin, K.A.; Koenig, K.M. Using organic acids to control subacute ruminal acidosis and fermentation in feedlot cattle fed a high-grain diet. J. Anim. Sci. 2015, 93, 3950. [Google Scholar] [CrossRef]
- Varki, A. Biological roles of oligosaccharides: All of the theories are correct. Glycobiology 1993, 3, 97–130. [Google Scholar] [CrossRef] [PubMed]
- Ransom-Jones, E.; Jones, D.L.; McCarthy, A.J.; McDonald, J.E. Thefibrobacteres: An important phylum of cellulose-degrading bacteria. Microb. Ecol. 2012, 63, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Prado, M.; Calsamiglia, S.; Ferret, A. Effects of Fiber Content and Particle Size of Forage on the Flow of Microbial Amino Acids from Continuous Culture Fermenters. J. Anim. Sci. 2004, 87, 1413–1424. [Google Scholar] [CrossRef] [Green Version]
- Naas, A.E.; Mackenzie, A.K.; Mravec, J.; Schuckel, J.; Willats, W.G.T.; Eijsink, V.G.H.; Pope, P.B. Do rumen bacteroidetes utilize an alternative mechanism for cellulose degradation? MBIO 2014, 5, 1401–1414. [Google Scholar] [CrossRef]
- Ludwig, W.; Schleifer, K.H.; Whitman, W.B. Revised Road Map to the Phylum Firmicutes. In Bergey’s Manual® of Systematic Bacteriology; Springer: New York, NY, USA, 2009; pp. 1–13. [Google Scholar]
- Pan, X.H.; Yang, L.; Xue, F.G.; Xin, H.R.; Jiang, L.S.; Xiong, B.H.; Beckers, Y. Relationship between thiamine and subacute ruminal acidosis induced by a high-grain diet in dairy cows. J. Anim. Sci. 2016, 99, 8790–8801. [Google Scholar] [CrossRef] [Green Version]
- Xue, F.; Pan, X.; Jiang, L.; Guo, Y.; Xiong, B.H. GC–MS analysis of the ruminal metabolome response to thiamine supplementation during high grain feeding in dairy cows. Metabolomics 2018, 14, 67. [Google Scholar] [CrossRef]
- Pan, X.; Nan, X.; Yang, L.; Jiang, L.; Xiong, B. Thiamine status, metabolism and application in dairy cows: A review. Br. J. Nutr. 2018, 120, 491–499. [Google Scholar] [CrossRef]
- Whitelaw, F.G.; Milne, J.S.; Chen, X.B. The effect of a rumen microbial fermentation on urea and nitrogen metabolism of sheep nourished by intragastric infusion. Exp. Physiol. 1991, 76, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, F.N.; Zinn, R.A.; Kim, Y.K. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 1986, 63, 1634. [Google Scholar] [CrossRef] [PubMed]
- Devant, M.; Ferret, A.; Gasa, J.; Calsamiglia, S.; Casals, R. Effects of protein concentration and degradability on performance, ruminal fermentation, and nitrogen metabolism in rapidly growing heifers fed high-concentrate diets from 100 to 230 kg body weight. J. Anim. Sci. 2000, 78, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Pisulewski, P.M.; Okorie, A.U.; Buttery, P.J.; Haresign, W.; Lewis, D. Ammonia concentration and protein synthesis in the rumen. Proc. Nutr. Soc. 2010, 32, 759–766. [Google Scholar] [CrossRef]
- Allison, M.J. Biosynthesis of amino acids by ruminal microorganisms. J. Anim. Sci. 1969, 29, 797–807. [Google Scholar] [CrossRef]
- Wallace, R.J. Ruminal microbiology, biotechnology, and ruminant nutrition: Progress and problems. J. Anim. Sci. 1994, 72, 2992–3003. [Google Scholar] [CrossRef]
- Ryle, M.; Ørskov, E.R. Energy Nutrition of Rumen Micro-Organisms; Springer: Dordrecht, The Netherlands, 1990; pp. 10–27. [Google Scholar]
- Lock, A.L.; Bauman, D.E. Modifying milk fat composition of dairy cows to enhance fatty acids beneficial to human health. Lipids 2004, 39, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Storry, J.E.; Rook, J.A.F. The relationship in the cow between milk-fat secretion and ruminal volatile fatty acids. Br. J. Nutr. 1966, 20, 217–228. [Google Scholar] [CrossRef]
- Kaufmann, S.; Rambeck, W.A. Iodine supplementation in chicken, pig and cow feed. J. Anim. Physiol. Anim. Nutr. 2010, 80, 147–152. [Google Scholar]
- Davis, S.R.; Collier, R.J.; McNamara, J.P.; Head, H.H.; Sussman, W. Effects of thyroxine and growth hormone treatment of dairy cows on milk yield, cardiac output and mammary blood flow. J. Anim. Sci. 1988, 66, 70. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Kelp | Control |
|---|---|---|
| Ingredients (%) | ||
| Corn silage | 27.0 | 29.1 |
| Oat hay | 20.4 | 22.8 |
| Kelp powder | 5.0 | — |
| Alfalfa haylage | 7.2 | 7.2 |
| Ground corn | 3.6 | 3.6 |
| Extruded soybeans | 2.6 | 2.6 |
| Soybean meal | 9.9 | 9.9 |
| Rapeseed meal | 4.5 | 4.5 |
| Cottonseed meal | 2.6 | 2.6 |
| Pressed corn | 2.2 | 2.2 |
| Cottonseed | 2.7 | 2.7 |
| Corn hull | 9.6 | 9.6 |
| Fat powder | 1.2 | 1.2 |
| Fatty acid calcium | 1.2 | 1.7 |
| Vitamin/mineral premix | 0.3 | 0.3 |
| Nutrient composition (%, unless otherwise stated) | ||
| NEL (Mcal/kg) | 1.40 | 1.41 |
| CP | 12.81 | 12.96 |
| EE | 3.42 | 3.57 |
| NDF | 36.78 | 38.14 |
| ADF | 20.69 | 22.15 |
| Items | CON | Kelp | SEM | p-Value |
|---|---|---|---|---|
| DMI (kg/day) | 16.2 | 16.6 | 0.468 | 0.114 |
| Milk production (kg/day) | 21.1 | 22.7 | 1.894 | 0.235 |
| Milk fat (%) | 3.8 | 3.9 | 0.126 | 0.146 |
| Milk protein(%) | 3.41 | 3.42 | 0.061 | 0.106 |
| Milk iodine(mg/L) | 0.07 b | 0.12 a | 0.020 | 0.029 |
| Ruminal pH | 6.45 | 6.38 | 0.194 | 0.216 |
| Ammonia-N (mg/dL) | 17.8 a | 13.1 b | 1.933 | <0.001 |
| Acetate (mmol/L) | 65.7 b | 67.1 a | 0.773 | 0.038 |
| Propionate (mmol/L) | 17.2 b | 19.2 a | 0.632 | 0.027 |
| A/P | 3.83 | 3.57 | 0.098 | 0.001 |
| Butyrate (mmol/L) | 16.3 | 16.1 | 0.137 | 0.356 |
| Isobutyrate (mmol/L) | 1.6 b | 3.9 a | 0.115 | 0.041 |
| Valerate(mmol/L) | 2.21 | 2.14 | 0.064 | 0.241 |
| Isovalerate (mmol/L) | 2.31 | 2.50 | 0.160 | 0.216 |
| TVFA (mmol/L) | 104.5 b | 110.8 a | 1.836 | 0.016 |
| Items | CON | Kelp | FC | Log 2 FC | SE | p-Value |
|---|---|---|---|---|---|---|
| ACE | 1182.5 | 1170.2 | 0.99 | −0.02 | 0.204 | 0.853 |
| Chao1 | 1192.4 | 1189.9 | 1.00 | 0.00 | 0.076 | 0.97 |
| Shannon | 5.42 | 5.31 | 0.98 | −0.03 | 0.08 | 0.398 |
| Simpson | 0.014 | 0.018 | 1.29 | 0.36 | 0.009 | 0.216 |
| SOBs | 1035.3 | 1016 | 0.98 | −0.03 | 0.12 | 0.759 |
| Items | CON | Kelp | FC | log2 FC | SE | p-Value |
|---|---|---|---|---|---|---|
| p__Bacteroidetes | 14570 a | 10110 b | 0.69 | −0.53 | 0.10 | <0.001 |
| p__Firmicutes | 7007 b | 12098 a | 1.73 | 0.79 | 0.13 | 0.008 |
| p__Fibrobacteres | 91.4 | 36.8 | 0.40 | −1.31 | 0.46 | 0.088 |
| p__Proteobacteria | 424.6 b | 1787.3 a | 4.21 | 2.07 | 0.53 | 0.024 |
| p__Spirochaetae | 179.3 | 242.1 | 1.35 | 0.43 | 0.32 | 0.408 |
| p__Saccharibacteria | 133.5 | 143 | 1.07 | 0.099 | 0.32 | 0.765 |
| p__Tenericutes | 264 | 214.4 | 0.81 | −0.30 | 0.52 | 0.462 |
| p__Verrucomicrobia | 33.8 | 19 | 0.56 | −0.83 | 0.31 | 0.277 |
| others | 39.4 | 54.2 | 1.38 | 0.46 | 0.27 | 0.286 |
| Items | CON | Kelp | FC | Log2 FC | SE | p-Value |
|---|---|---|---|---|---|---|
| g__Prevotella | 9897.2 a | 6376.8 b | 0.64 | −0.63 | 0.08 | <0.001 |
| g__Ruminococcaceae | 1470.3 b | 3111.1 a | 2.12 | 1.08 | 0.17 | 0.021 |
| g__Ruminococcus | 539.8 b | 2603.8 a | 4.82 | 2.27 | 0.25 | 0.004 |
| g__Bacteroidales | 1969.7 a | 1064.1 b | 0.54 | −0.89 | 0.15 | 0.002 |
| g__Prevotellaceae | 1472 | 1199.2 | 0.81 | −0.30 | 0.15 | 0.185 |
| g__Succinivibrionaceae | 636.3 | 1238.8 | 1.95 | 0.96 | 0.56 | 0.247 |
| g__Lachnospiraceae | 566.4 b | 1069.2 a | 1.89 | 0.92 | 0.15 | 0.008 |
| g__Succiniclasticum | 626.6 | 653.8 | 1.04 | 0.06 | 0.33 | 0.876 |
| g__Eubacterium | 502.7 | 521.8 | 1.04 | 0.05 | 0.27 | 0.767 |
| g__Rikenellaceae | 600.5 a | 315.6 b | 0.53 | −0.93 | 0.13 | 0.016 |
| g__Christensenellaceae | 430.8 | 428.6 | 0.99 | −0.01 | 0.28 | 0.986 |
| g__Selenomonas | 437.1 | 240.3 | 0.55 | −0.86 | 0.28 | 0.012 |
| g__Acetitomaculum | 171.7 | 446.2 | 2.60 | 1.38 | 0.18 | 0.121 |
| g__Butyrivibrio | 177.1 | 255.1 | 1.44 | 0.53 | 0.31 | 0.615 |
| g__Treponema | 285.8a | 124 b | 0.43 | −1.20 | 0.42 | 0.022 |
| g__Erysipelotrichaceae | 140.8 | 204.2 | 1.45 | 0.54 | 0.51 | 0.539 |
| g__Anaeroplasma | 214.9 a | 113.3 b | 0.53 | −0.92 | 0.35 | 0.003 |
| g__Lachnobacterium | 170.3 | 143.4 | 0.84 | −0.25 | 0.18 | 0.453 |
| g__Candidatus Saccharimonas | 169.6 | 111 | 0.65 | −0.61 | 0.22 | 0.05 |
| g__Saccharofermentans | 146.9 a | 81.2 b | 0.55 | −0.86 | 0.31 | 0.016 |
| g__Sharpea | 31.2 b | 189.7 a | 6.08 | 2.60 | 0.22 | 0.016 |
| g__Roseburia | 84.6 | 123.2 | 1.46 | 0.54 | 0.6 | 0.535 |
| g__Ruminiclostridium | 50.5 | 150.9 | 2.99 | 1.58 | 0.49 | 0.159 |
| g__Schwartzia | 93.5 | 77.5 | 0.83 | −0.27 | 0.33 | 0.642 |
| g__Lachnospiraceae | 79.8 | 81.7 | 1.02 | 0.03 | 0.41 | 0.868 |
| g__Fibrobacter | 77.3 | 52.9 | 0.68 | −0.55 | 0.14 | 0.654 |
| g__Mogibacterium | 58 | 64.9 | 1.12 | 0.16 | 0.53 | 0.768 |
| g__Succinivibrio | 60 | 34 | 0.57 | −0.82 | 0.28 | 0.338 |
| others | 3665.1 | 3447.5 | 0.94 | −0.088 | 0.48 | 0.471 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, F.; Sun, F.; Jiang, L.; Hua, D.; Wang, Y.; Nan, X.; Zhao, Y.; Xiong, B. Effects of Partial Replacment of Dietary Forage Using Kelp Powder (Thallus laminariae) on Ruminal Fermentation and Lactation Performances of Dairy Cows. Animals 2019, 9, 852. https://doi.org/10.3390/ani9100852
Xue F, Sun F, Jiang L, Hua D, Wang Y, Nan X, Zhao Y, Xiong B. Effects of Partial Replacment of Dietary Forage Using Kelp Powder (Thallus laminariae) on Ruminal Fermentation and Lactation Performances of Dairy Cows. Animals. 2019; 9(10):852. https://doi.org/10.3390/ani9100852
Chicago/Turabian StyleXue, Fuguang, Fuyu Sun, Linshu Jiang, Dengke Hua, Yue Wang, Xuemei Nan, Yiguang Zhao, and Benhai Xiong. 2019. "Effects of Partial Replacment of Dietary Forage Using Kelp Powder (Thallus laminariae) on Ruminal Fermentation and Lactation Performances of Dairy Cows" Animals 9, no. 10: 852. https://doi.org/10.3390/ani9100852