Two Novel SNPs in RET Gene Are Associated with Cattle Body Measurement Traits


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Samples
2.3. DNA Isolation and Genomic DNA Pools Construction
2.4. SNP Detection and Genotyping
2.5. Statistical Analysis
3. Results
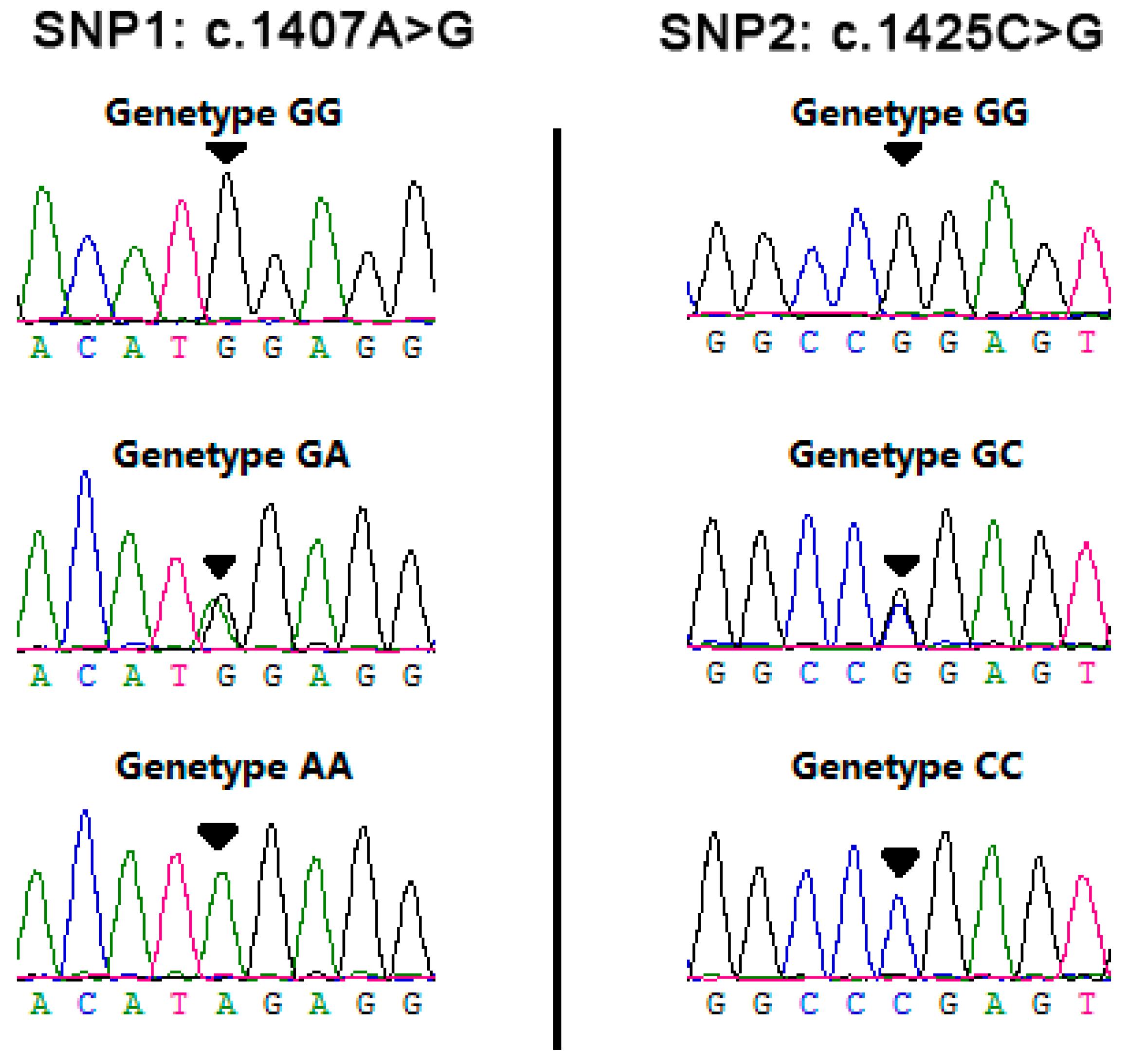
3.1. Identification of Genetic Variants in RET
3.2. Genetic Characteristics of SNPs Detected in Bovine RET Gene
3.3. Association Analysis between Polymorphisms and Body Measurement Traits
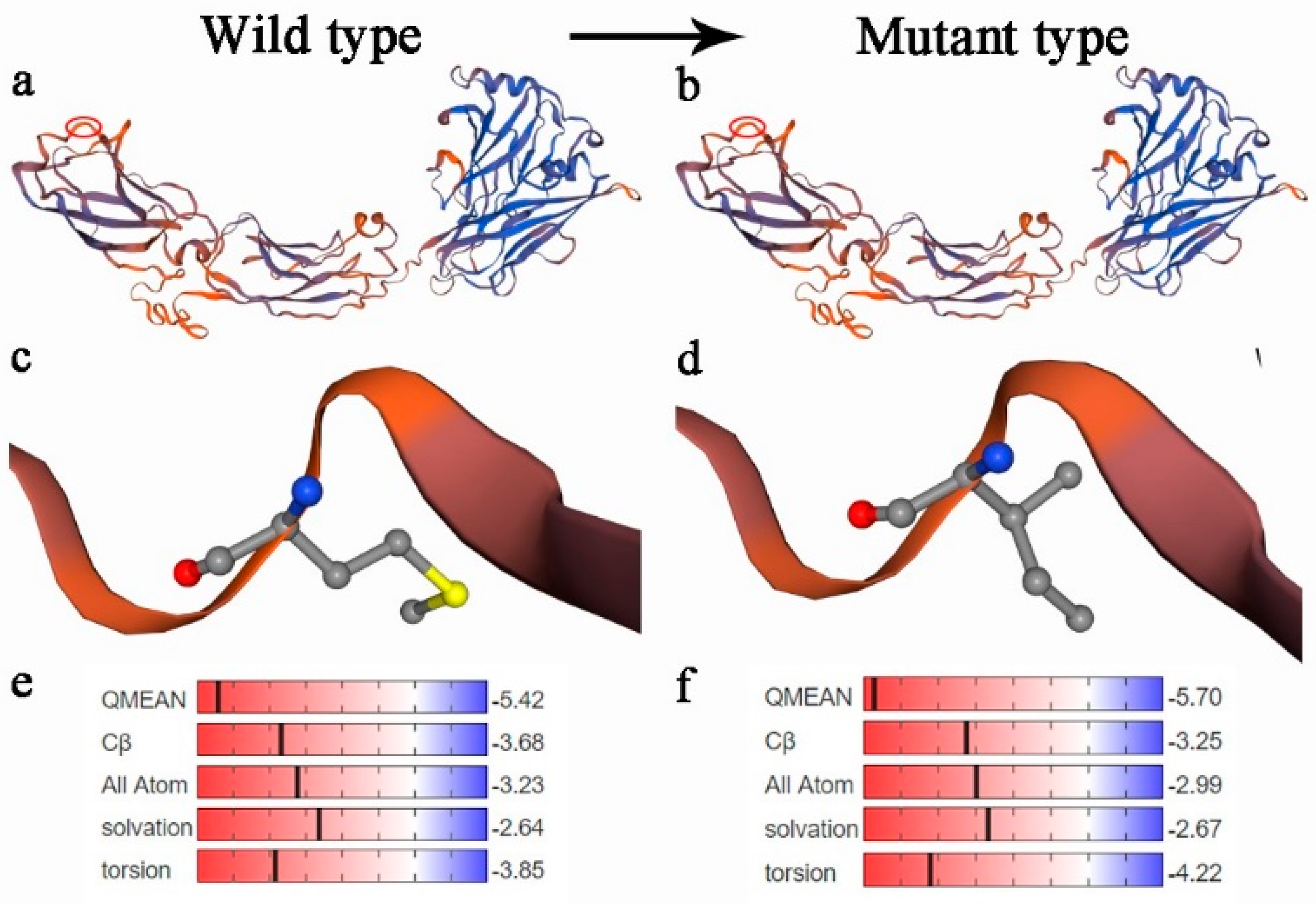
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanders, K.M.; Koh, S.D.; Ward, S.M. Interstitial cells of cajal as pacemakers in the gastrointestinal tract. Annu. Rev. Physiol. 2006, 68, 307–343. [Google Scholar] [CrossRef] [PubMed]
- Heanue, T.A.; Pachnis, V. Enteric nervous system development and hirschsprung’s disease: Advances in genetic and stem cell studies. Nat. Rev. Neurosci. 2007, 8, 466. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D. Genes and lineages in the formation of the enteric nervous system. Curr. Opin. Neurobiol. 1997, 7, 101–109. [Google Scholar] [CrossRef]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, F.R.; Kernohan, J.W. Myenteric plexus in congenital megacolon: Study of eleven cases. Arch. Intern. Med. 1948, 82, 75–111. [Google Scholar] [CrossRef]
- Ross, A.J. Surgery of Infants and Children: Scientific Principles and Practice. Ann. Surg. 1997, 226, 208–209. [Google Scholar] [CrossRef]
- Martucciello, G.; Bicocchi, M.; Dodero, P.; Lerone, M.; Cirillo, M.S.; Puliti, A.; Gimelli, G.; Romeo, G.; Jasonni, V. Total colonic aganglionosis associated with interstitial deletion of the long arm of chromosome 10. Pediatr. Surg. Int. 1992, 7, 308–310. [Google Scholar] [CrossRef]
- Chakravarti, A. Hirschsprung’s disease. In The Metabolic and Molecular Basis of Inherited Disease; McGraw-Hill Professional: New York, NY, USA, 2001; pp. 6231–6255. [Google Scholar]
- Emison, E.S.; Garcia-Barcelo, M.; Grice, E.A.; Lantieri, F.; Amiel, J.; Burzynski, G.; Fernández, R.M.; Hao, L.; Kashuk, C.; West, K.; et al. Differential Contributions of Rare and Common, Coding and Noncoding Ret Mutations to Multifactorial Hirschsprung Disease Liability. Am. J. Hum. Genet. 2010, 87, 60–74. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, A.; Jiang, Q.; Chatterjee, S.; Chakraborty, P.; Sosa, M.X.; Berrios, C.; Chakravarti, A. Population variation in total genetic risk of Hirschsprung disease from common RET, SEMA3 and NRG1 susceptibility polymorphisms. Hum. Mol. Genet. 2015, 24, 2997–3003. [Google Scholar] [CrossRef] [Green Version]
- Pachnis, V.; Mankoo, B.; Costantini, F. Expression of the c-ret proto-oncogene during mouse embryogenesis. Development 1993, 119, 1005. [Google Scholar]
- Eng, C. RET proto-oncogene in the development of human cancer. J. Clin. Oncol. 1999, 17, 380. [Google Scholar] [CrossRef] [PubMed]
- Ponder, B.A. The phenotypes associated with ret mutations in the multiple endocrine neoplasia type 2 syndrome. Cancer Res. 1999, 59, 1736–1742. [Google Scholar]
- Frank-Raue, K.; Rondot, S.; Raue, F. Molecular genetics and phenomics of RET mutations: Impact on prognosis of MTC. Mol. Cell. Endocrinol. 2010, 322, 2–7. [Google Scholar] [CrossRef] [PubMed]
- De Groot, J.W.B.; Links, T.P.; Plukker, J.T.M.; Lips, C.J.M.; Hofstra, R.M.W. RET as a Diagnostic and Therapeutic Target in Sporadic and Hereditary Endocrine Tumors. Endocr. Rev. 2006, 27, 535–560. [Google Scholar] [CrossRef] [Green Version]
- Borrego, S.; Wright, F.A.; Fernández, R.M.; Williams, N.; López-Alonso, M.; Davuluri, R.; Antiñolo, G.; Eng, C. A founding locus within the ret proto-oncogene may account for a large proportion of apparently sporadic hirschsprung disease and a subset of cases of sporadic medullary thyroid carcinoma. Am. J. Hum. Genet. 2003, 72, 88–100. [Google Scholar] [CrossRef]
- Emison, E.S.; McCallion, A.S.; Kashuk, C.S.; Bush, R.T.; Grice, E.; Lin, S.; Portnoy, M.E.; Cutler, D.J.; Green, E.D.; Chakravarti, A. A common sex-dependent mutation in a RET enhancer underlies Hirschsprung disease risk. Nature 2005, 434, 857–863. [Google Scholar] [CrossRef]
- Margawati, E.T. A global strategy of using molecular genetic information to improve genetics in livestock. Reprod. Domest. Anim. 2012, 47, 7–9. [Google Scholar] [CrossRef]
- Fang, X.; Lai, Z.; Liu, J.; Zhang, C.; Li, S.; Wu, F.; Zhou, Z.; Lei, C.; Dang, R. A Novel 13 bp Deletion within the NR6A1 Gene Is Significantly Associated with Growth Traits in Donkeys. Animals 2019, 9, 681. [Google Scholar] [CrossRef]
- Gilbert, R.P.; Bailey, D.R.; Shannon, N.H. Linear body measurements of cattle before and after 20 years of selection for postweaning gain when fed two different diets. J. Anim. Sci. 1993, 71, 1712–1720. [Google Scholar] [CrossRef]
- Nei, M.; Roychoudhury, A.K. Sampling Variances of Heterozygosity and Genetic Distance. Genetics 1974, 76, 379–390. [Google Scholar]
- Yong, Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Li, M.; Wang, S.; Xu, Y.; Lan, X.; Li, Z.; Lei, C.; Yang, D.; Jia, Y.; Chen, H. Association analysis of bovine Foxa2 gene single sequence variant and haplotype combinations with growth traits in Chinese cattle. Gene 2014, 536, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.B.; Ma, Y.; Yang, T.; Lin, F.; Fu, W.W.; Xu, Y.J.; Li, F.; Li, J.Y.; Gao, S.X. Tissue expression and predicted protein structures of the bovine ANGPTL3 and association of novel SNPs with growth and meat quality traits. Animals 2015, 9, 1285–1297. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chen, N.; Li, X.; An, S.; Zhao, M.; Sun, T.; Hao, R.; Ma, Y. Two novel snps of pparγ significantly affect weaning growth traits of nanyang cattle. Anim. Biotechnol. 2018, 29, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, S.; Zhang, G.; Fan, Y.; Gao, Y.; Huang, Y.; Lan, X.; Lei, C.; Ma, Y.; Dang, R. Exploring insertions and deletions (indels) of MSRB3 gene and their association with growth traits in four Chinese indigenous cattle breeds. Arch. Anim. Breed. 2019, 62, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, X.; Wang, H.; He, H.; Li, Z.; Chen, L. Expression analysis, single nucleotide polymorphisms and combined genotypes in candidate genes and their associations with growth and carcass traits in qinchuan cattle. Mol. Biol. Rep. 2013, 40, 2335–2346. [Google Scholar] [CrossRef]
- Edery, P.; Lyonnet, S.; Mulligan, L.M.; Pelet, A.; Dow, E.; Abel, L.; Holder, S.; Nihoul-Fékété, C.; Ponder, B.A.; Munnich, A. Mutations of the ret proto-oncogene in hirschsprung’s disease. Nature 1994, 367, 378. [Google Scholar] [CrossRef]
- Ruiz-Ferrer, M.; Fernández, R.M.; Antiñolo, G.; López-Alonso, M.; Eng, C.; Borrego, S. A complex additive model of inheritance for Hirschsprung disease is supported by both RET mutations and predisposing RET haplotypes. Genet. Med. 2006, 8, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Fitze, G.; Cramer, J.; Ziegler, A.; Schierz, M.; Schreiber, M.; Kuhlisch, E.; Roesner, D.; Schackert, H.K. Association between c135g/a genotype and ret proto-oncogene germline mutations and phenotype of hirschsprung’s disease. Lancet 2002, 359, 1200–1205. [Google Scholar] [CrossRef]
- Sancandi, M.; Griseri, P.; Pesce, B.; Patrone, G.; Puppo, F.; Lerone, M.; Martucciello, G.; Romeo, G.; Ravazzolo, R.; Devoto, M. Single nucleotide polymorphic alleles in the 5′ region of the ret proto-oncogene define a risk haplotype in hirschsprung’s disease. J. Med. Genet. 2003, 40, 714–718. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5’–3’) | Tm (°C) | Length (bp) | Location |
|---|---|---|---|---|
| R1 | F:CGGCTCTGGTCTCCTAAC R: GCCACACAACCCCACT | 51 | 609 | Exon1 |
| R2 | F:GCTGGGAAAGCTGATCTGC R:AAGGAGGGACAAGGAGAGG | 56 | 440 | Exon2 |
| R3 | F:TGTGAGGACAAGGGAGCA R:AATGGATGATGGGGTGGT | 63 | 633 | Exon3 |
| R4 | F:CTGTGCTTGGAGGCTGTG R:ATGGAGGGACCCAGATGC | 63 | 477 | Exon4 |
| R5 | F: CCTATGGGCTCTGGCTTTC R:AAGGCTGGGAAGAGGGTT | 63 | 396 | Exon5 |
| R6 | F:ATGTGTCCAGGGAATGCT R: CTGGCACTGCTCTTTCAT | 57 | 596 | Exon6 |
| R7 | F:CTGTCCTGGGACTCAAGCTG R:TACAGGACCGCACCCTTCTA | 64 | 438 | Exon7 |
| R8 | F: CGCAAAGCAGGTATTCAC R: GAGCACAGAGGAAAACT | 51 | 536 | Exon8 |
| R9 | F: AGCCTCGTTTGGTCTCC R: GAATGTGGGTCAAGCCG | 61 | 578 | Exon9 |
| R10 | F: GAGTGGGCTGCTGAGTG R: AGGGAGGCATGAGGATC | 56 | 212 | Exon10 |
| R11 | F: TGGCTCTGTTGGGAGTTGG R: TGAAAGGCAGCACGGTG | 56 | 523 | Exon11 |
| R12 | F: TGAGCATACGGAGTCCAGC R: CACACCAGCCCACATCAT | 58 | 591 | Exon12 |
| R13 | F: ATAGCCCAGCCCAGAAGG R:AGTAGGTGCCCAGGAGACA | 60 | 699 | Exon13 |
| R14-15 | F: AGGAACCTGGCGGGCTAT R: GGGGATGTGGACTGGAAG | 56 | 762 | Exon14- Exon 15 |
| R16 | F: TTGGGAAGTGGTGCGTG R: GGTCATTCTGGACGGTTGG | 58 | 454 | Exon16 |
| R17 | F: ACCGTGGGCACTTGGAC R: CCCCAGCATCTGCCATC | 62 | 465 | Exon17 |
| R18 | F:GCTTGGCAGGTGTTTGTG R:GAAGGATAAAGGCTCGTGC | 58 | 542 | Exon18 |
| R19 | F: GCGAGAGGTTGATAGGTGC R: CGCCCTCTCCTCTAACATCT | 56 | 544 | Exon19 |
| Name | Breeds (Sizes) | Genotypic Frequencies | Allelic Frequencies | Ne | He | PIC | χ2 (HWE) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| AA | AG | GG | A | G | ||||||
| c.1407A>G | QC/225 | 0.5200 | 0.4267 | 0.0533 | 0.7333 | 0.2667 | 1.6423 | 0.3911 | 0.3146 | 1.8595 |
| NY/117 | 0.7265 | 0.2564 | 0.0171 | 0.8547 | 0.1453 | 1.3304 | 0.2484 | 0.2175 | 0.1225 | |
| CC | CG | GG | C | G | ||||||
| c.1425C>G | QC/225 | 0.5289 | 0.4178 | 0.0533 | 0.7378 | 0.2622 | 1.6311 | 0.3869 | 0.3121 | 1.4308 |
| NY/117 | 0.8205 | 0.1538 | 0.0256 | 0.8974 | 0.1026 | 1.2256 | 0.1841 | 0.1671 | 3.1578 | |
| Breed | Haplotype | SNP1 | SNP2 | Frequency |
|---|---|---|---|---|
| QC | Hap1 | A | C | 0.733 |
| Hap2 | G | G | 0.262 | |
| NY | Hap1 | A | C | 0.850 |
| Hap2 | G | G | 0.098 | |
| Hap3 | G | C | 0.047 |
| Genotypes | BH (cm) | HH (cm) | BL (cm) | ChC (cm) | RL (cm) |
|---|---|---|---|---|---|
| AA | 129.92 ± 0.50 a | 129.30 ± 0.48 a | 154.18 ± 0.90 | 198.52 ± 1.27 a | 50.86 ± 0.31 |
| AG | 130.51 ± 0.48 ab | 129.64 ± 0.50 ab | 156.57 ± 0.91 | 202.67 ± 1.17 b | 50.50 ± 0.37 |
| GG | 133.67 ± 2.12 b | 132.89 ± 2.33 b | 159.44 ± 4.95 | 208.33 ± 2.55 b | 51.33 ± 0.97 |
| p vaule | 0.035 | 0.042 | 0.109 | 0.011 | 0.645 |
| CC | 129.72 ± 0.50 a | 129.18 ± 0.47 a | 153.67 ± 0.95 | 197.80 ± 1.34 a | 50.84 ± 0.31 |
| CG | 130.66 ± 0.49 ab | 129.71 ± 0.52 ab | 156.78 ± 0.85 | 203.09 ± 1.16 b | 50.53 ± 0.38 |
| GG | 133.67 ± 2.12 b | 132.88 ± 2.33 b | 159.44 ± 4.95 | 208.33 ± 2.55 b | 51.33 ± 0.97 |
| p vaule | 0.027 | 0.036 | 0.091 | 0.003 | 0.622 |
| Genotypes | BH (cm) | BL (cm) | ChC (cm) | AC (cm) | HW (cm) | HuW (cm) | HH (cm) | BW (cm) |
|---|---|---|---|---|---|---|---|---|
| AA | 128.95 ± 0.97 | 140.79 ± 1.30 | 176.06 ± 1.37 a | 221.95 ± 4.15 | 46.92 ± 0.76 | 27.67 ± 0.35 a | 131.27 ± 1.78 | 410.45 ± 11.33 |
| AG | 126.83 ± 1.92 | 137.33 ± 2.41 | 169.48 ± 3.07 b | 207.95 ± 6.57 | 43.84 ± 1.72 | 26.00 ± 0.63 b | 125.50 ± 3.29 | 382.61 ± 17.62 |
| GG | 127.50 ± 1.50 | 140.00 ± 9.00 | 175.50 ± 0.50 ab | 234.50 ± 1.50 | 46.00 ± 1.00 | 28.00 ± 0.41 ab | 130.00 ± 1.00 | 416.00 ± 11.00 |
| p vaule | 0.334 | 0.267 | 0.050 | 0.138 | 0.174 | 0.043 | 0.157 | 0.227 |
| CC | 127.97 ± 0.99 | 139.92 ± 1.30 | 174.85 ± 1.42 | 216.21 ± 4.29 a | 45.56 ± 0.90 | 27.37 ± 0.36 | 128.90 ± 1.86 | 403.81 ± 10.90 |
| CG | 123.88 ± 2.34 | 139.80 ± 3.24 | 172.41 ± 4.33 | 222.22 ± 4.96 ab | 47.50 ± 1.36 | 26.83 ± 0.66 | 131.55 ± 3.47 | 402.28 ± 25.32 |
| GG | 128.33 ± 1.20 | 139.83 ± 5.19 | 176.83 ± 1.36 | 234.50 ± 1.50 b | 46.00 ± 1.00 | 28.33 ± 0.33 | 130.00 ± 1.00 | 410.33 ± 8.51 |
| p vaule | 0.743 | 0.974 | 0.778 | 0.001 | 0.626 | 0.722 | 0.816 | 0.992 |
| Genotypes (Number) | BH (cm) | HH (cm) | BL (cm) | ChC (cm) | RL (cm) |
|---|---|---|---|---|---|
| AA-CC (113) | 129.88 ± 0.51 a | 129.36 ± 0.48 ab | 154.14 ± 0.93 | 198.44 ± 1.31 a | 50.87 ± 0.31 |
| AA-CG (4) | 131.00 ± 2.38 ab | 127.50 ± 3.61 ab | 155.25 ± 1.03 | 200.75 ± 3.85 ab | 50.75 ± 1.37 |
| AG-CC (6) | 126.66 ± 2.10 a | 125.83 ± 1.55 a | 146.00 ± 7.15 | 185.83 ± 8.73 a | 50.20 ± 1.31 |
| AG-CG (90) | 130.65 ± 0.51 ab | 129.82 ± 0.51 ab | 156.86 ± 0.89 | 203.21 ± 1.20 b | 50.51 ± 0.39 |
| GG-GG (12) | 133.67 ± 2.12 b | 132.89 ± 2.33 b | 159.44 ± 4.95 | 208.33 ± 2.55 b | 51.33 ± 0.97 |
| p vaule | 0.044 | 0.033 | 0.143 | 0.002 | 0.922 |
| Genotypes (Number) | BH (cm) | BL (cm) | ChC (cm) | AC (cm) | HW (cm) | HuW(cm) | HH (cm) | BW (cm) |
|---|---|---|---|---|---|---|---|---|
| AA-CC (85) | 128.95 ± 0.97 a | 140.79 ± 1.30 | 176.06 ± 1.37 a | 221.95 ± 4.15 a | 46.92 ± 0.76 a | 27.67 ± 0.35 a | 131.27 ± 1.78 a | 410.45 ± 11.33 |
| AG-CC (11) | 120.15 ± 3.99 b | 132.50 ± 5.05 | 164.55 ± 6.24 b | 194.12 ± 10.10 b | 39.14 ± 2.45 b | 24.25 ± 1.35 b | 119.62 ± 4.59 b | 345.23 ± 34.57 |
| AG-CG (18) | 129.88 ± 2.34 a | 139.80 ± 3.24 | 172.41 ± 4.33 ab | 222.22 ± 4.96 a | 47.50 ± 1.36 a | 26.83 ± 0.66 a | 131.55 ± 3.47 a | 402.28 ± 25.17 |
| GG-GG (2) | 127.50 ± 1.50 ab | 140.00 ± 9.00 | 175.50 ± 0.50 ab | 234.50 ± 1.50 a | 46.00 ± 1.00 ab | 28.00 ± 0.01 ab | 130.00 ± 1.00 ab | 416.00 ± 11.00 |
| p vaule | 0.043 | 0.282 | 0.018 | 0.008 | 0.004 | 0.013 | 0.025 | 0.359 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Huang, B.; Bai, F.; Wu, F.; Zhou, Z.; Lai, Z.; Li, S.; Qu, K.; Jia, Y.; Lei, C.; et al. Two Novel SNPs in RET Gene Are Associated with Cattle Body Measurement Traits. Animals 2019, 9, 836. https://doi.org/10.3390/ani9100836
Gao Y, Huang B, Bai F, Wu F, Zhou Z, Lai Z, Li S, Qu K, Jia Y, Lei C, et al. Two Novel SNPs in RET Gene Are Associated with Cattle Body Measurement Traits. Animals. 2019; 9(10):836. https://doi.org/10.3390/ani9100836
Chicago/Turabian StyleGao, Yuan, Bizhi Huang, Fuxia Bai, Fei Wu, Zihui Zhou, Zhenyu Lai, Shipeng Li, Kaixing Qu, Yutang Jia, Chuzhao Lei, and et al. 2019. "Two Novel SNPs in RET Gene Are Associated with Cattle Body Measurement Traits" Animals 9, no. 10: 836. https://doi.org/10.3390/ani9100836