Effects of Chromium-Loaded Chitosan Nanoparticles on the Intestinal Electrophysiological Indices and Glucose Transporters in Broilers

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Chromium-Loaded Chitosan Nanoparticles
2.2. Experimental Animals, Grouping, Diet, and Management
2.3. Sample Collection and Processing of Tissue
2.4. Measurement of Electrophysiological Indices
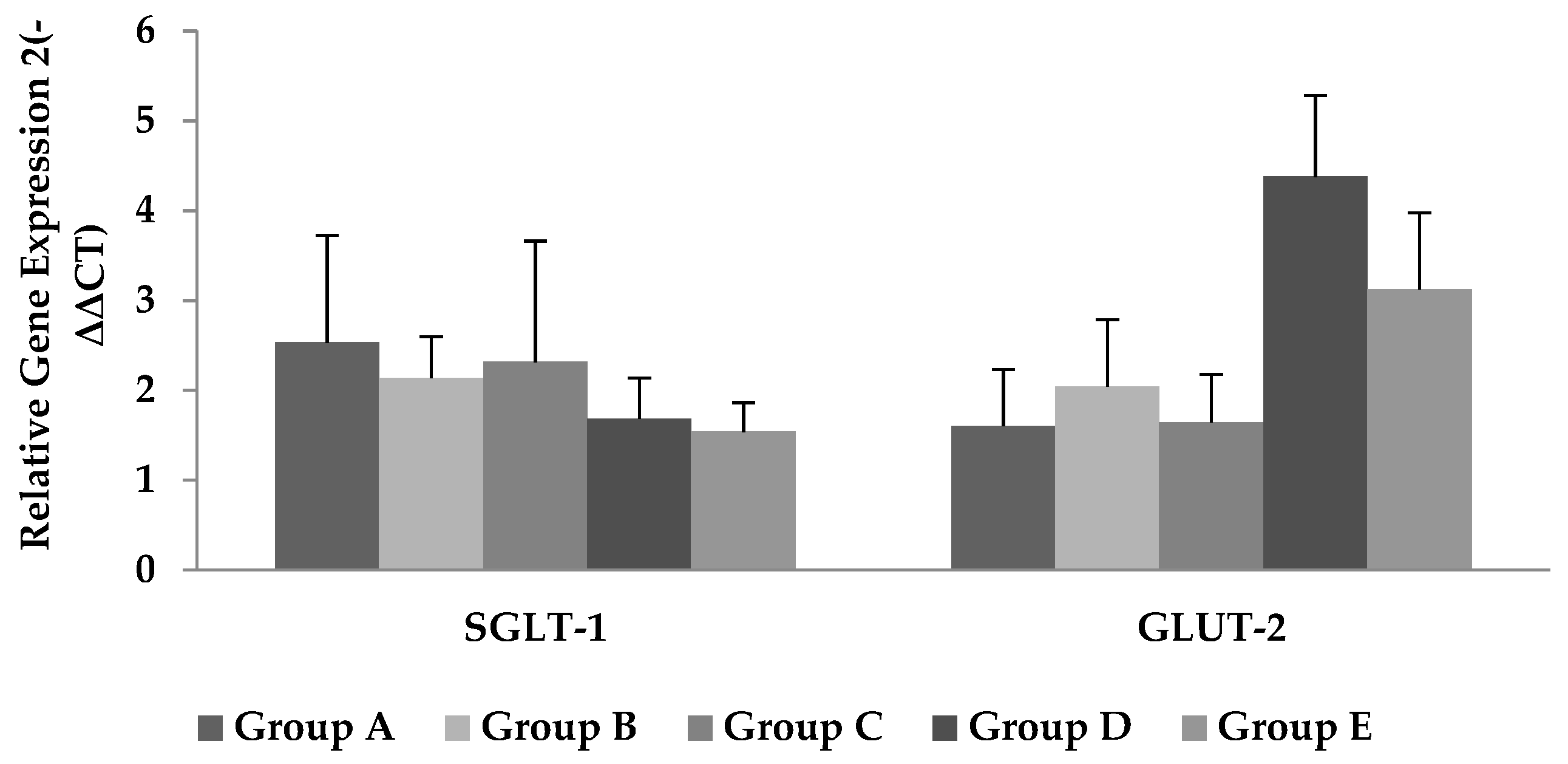
2.5. Extraction of RNA and Quantification of Glucose Transporters
2.6. Quantification of Tissues Glycogen and Serm Glucose
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brooks, M.A.; Grimes, J.L.; Lloyd, K.E.; Krafka, K.; Lamptey, A.; Spears, J.W. Chromium propionate in broilers: Effect on insulin sensitivity. Poult. Sci. 2016, 95, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G. Perspectives on the endocrinology of poultry growth and metabolism. Gen. Comp. Endocrinol. 2009, 163, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.B. The bioinorganic chemistry of chromium (III). Polyhedron 2001, 20, 1–26. [Google Scholar] [CrossRef]
- Lien, T.F.; Horng, Y.M.; Yang, K.H. Performance, serum characteristics, carcase traits and lipid metabolism of broilers as affected by supplement of chromium picolinate. Br. Poult. Sci. 1999, 40, 357–363. [Google Scholar] [CrossRef]
- Zheng, C.; Huang, Y.; Xiao, F.; Lin, X.; Lloyd, K. Effects of supplemental chromium source and concentration on growth, carcass characteristics, and serum lipid parameters of broilers reared under normal conditions. Biol. Trace Elem. Res. 2016, 169, 352–358. [Google Scholar] [CrossRef]
- Sahin, K.; Sahin, N.; Onderci, M.; Gursu, F.; Cikim, G. Optimal dietary concentration of chromium for alleviating the effect of heat stress on growth, carcass qualities, and some serum metabolites of broiler chickens. Biol. Trace Elem. Res. 2002, 89, 53–64. [Google Scholar] [CrossRef]
- Debski, B.; Zalewski, W.; Gralak, M.A.; Kosla, T. Chromium yeast supplementation of broilers in an industrial farming system. J. Trace Elem. Med. Biol. 2004, 18, 47–51. [Google Scholar] [CrossRef]
- Toghyani, M.; Toghyani, M.; Shivazad, M.; Gheisari, A.; Bahadoran, R. Chromium supplementation can alleviate the negative effects of heat stress on growth performance, carcass traits, and meat lipid oxidation of broiler chicks without any adverse impacts on blood constituents. Biol. Trace. Elem. Res. 2012, 146, 171–180. [Google Scholar] [CrossRef]
- Orhan, C.; Tuzcu, M.; Deeh, P.B.D.; Sahin, N.; Komorowski, J.R.; Sahin, K. Organic Chromium Form Alleviates the Detrimental Effects of Heat Stress on Nutrient Digestibility and Nutrient Transporters in Laying Hens. Biol. Trace Elem. Res. 2019, 189, 529–537. [Google Scholar] [CrossRef]
- Amat, C.; Planas, J.M.; Moreto, M. Kinetics of hexose uptake by the small and large intestine of the chicken. Am. J. Physiol. 1996, 271, R1085–R1089. [Google Scholar] [CrossRef]
- Majeed, K.A.; Yousaf, M.S.; Zaneb, H.; Rabbani, I.; Rehman, Z.U.; Rehman, H. In vitro evaluation of the effect of dibutyl phthalate on electrogenic sodium-linked glucose transport in isolated rabbit ileum. Toxicol. Environ. Chem. 2017, 99, 1389–1396. [Google Scholar] [CrossRef]
- Wang, M.Q.; Wang, C.; Li, H.; Du, Y.J.; Tao, W.J.; Ye, S.S.; He, Y.D. Effects of chromium-loaded chitosan nanoparticles on growth, blood metabolites, immune traits and tissue chromium in finishing pigs. Biol. Trace Elem. Res. 2012, 149, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Zaneb, H.; Masood, S.; Yousaf, M.S.; Rehman, H.F.; Rehman, H. Effect of Moringa oleifera leaf powder supplementation on growth performance and intestinal morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2017, 101, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.; Rosenkranz, C.; Böhm, J.; Zentek, J. Dietary inulin affects the morphology but not the sodium-dependent glucose and glutamine transport in the jejunum of broilers. Poult. Sci. 2007, 86, 118–122. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J. Intestinal structure and function of broiler chickens on diets supplemented with a synbiotic containing Enterococcus faecium and oligosaccharides. Int. J. Mol. Sci. 2008, 9, 2205–2216. [Google Scholar] [CrossRef]
- Shibata, M.; Takahashi, T.; Kozakai, T.; Kakudo, M.; Kasuga, S.; Azuma, Y.; Kurose, Y. Active transport of glucose across the jejunal epithelium decreases with age in broiler chickens. Poult. Sci. 2019, 98, 2570–2576. [Google Scholar] [CrossRef]
- Ebrahimi, R.; Faseleh-Jahromi, M.; Liang, J.B.; Soleimani-Farjam, A.; Shokryazdan, P.; Idrus, Z. Effect of dietary lead on intestinal nutrient transporters mRNA expression in broiler chickens. Bio. Med. Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Soleimani, A.F.; Zulkifli, I.; Omar, A.R.; Raha, A.R. Neonatal feed restriction modulates circulating levels of corticosterone and expression of glucocorticoid receptor and heat shock protein 70 in aged Japanese quail exposed to acute heat stress. Poult. Sci. 2011, 90, 1427–1434. [Google Scholar] [CrossRef]
- Bennett, L.W.; Keirs, R.W.; Peebles, E.D.; Gerard, P.D. Methodologies of tissue preservation and analysis of the glycogen content of the broiler chick liver. Poult. Sci. 2007, 86, 2653–2665. [Google Scholar] [CrossRef]
- Dreiling, C.E.; Brown, D.E.; Casale, L.; Kelly, L. Muscle glycogen: Comparison of iodine binding and enzyme digestion assays and application to meat samples. Meat Sci. 1987, 20, 167–177. [Google Scholar] [CrossRef]
- Vincent, J.B. The Biochemistry of Chromium. J. Nutr. 2000, 130, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Lukashi, H.C. Chromium as a supplement. Annu. Rev. Nutr. 1999, 19, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Amatya, J.; Haldar, S.; Ghosh, T. Effects of chromium supplementation from inorganic and organic sources on nutrient utilization, mineral metabolism and meat quality in broiler chickens exposed to natural heat stress. Anim. Sci. 2004, 79, 241–253. [Google Scholar] [CrossRef]
- Ahmed, N.; Haldar, S.; Pakhira, M.; Ghosh, T. Growth performances, nutrient utilization and carcass traits in broiler chickens fed with a normal and a low energy diet supplemented with inorganic chromium (as chromium chloride hexahydrate) and a combination of inorganic chromium and ascorbic acid. J. Agric. Sci. 2005, 143, 427–439. [Google Scholar] [CrossRef]
- Pappenheimer, J.R. On the coupling of membrane digestion with intestinal absorption of sugar and amino acids. Am. J. Physiol. 1993, 265, G409–G417. [Google Scholar] [CrossRef]
- Garriga, C.; Rovira, N.; Moreto´, M.; Planas, J.M. Expression of Na+-D-glucose cotransporter in brush-border membrane of the chicken intestine. Am. J. Physiol. 1999, 276, R627–R631. [Google Scholar] [CrossRef]
- Soriano-Garcia, J.F.; Torras-Llort, M.; Moreto´, M.; Ferrer, R. Regulation of L-methionine and L-lysine uptake in chicken jejunal brush border by dietary methionine. Am. J. Physiol. 1999, 77, R1654–R1661. [Google Scholar] [CrossRef]
- Awad, W.A.; Rehman, H.; Bo¨hm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of luminal deoxynivalenol and L-proline on electrophysiological parameters in the jejunums of laying hens. Poult. Sci. 2005, 84, 928–932. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Amado, M.A.; Del Castillo, J.R.; Perez, M.E.; Dominguez-Bello, M.G. Intestinal D-glucose and L-alanine transport in Japanese quail (Coturnix coturnix). Poult. Sci. 2005, 84, 947–950. [Google Scholar] [CrossRef]
- Mehmood, W.; Rabbani, I.; Stumpff, F.; Yousaf, M.S.; Zaneb, H.; Majeed, K.A.; Tahir, S.K.; Rashid, M.A.; Rehman, H. In Vitro Effects of Zinc Oxide Nanoparticles on Electrophysiological Indices and Sodium-Dependent Glucose Transport Across Jejunal Mucosa in Laying Hens. J. Appl. Poult. Res. 2018, 28, 271–277. [Google Scholar] [CrossRef]
- Gammelgaard, B.; Jensen, K.; Steffansen, B. In vitro metabolism and permeation studies in rat jejunum: Organic chromium compared to inorganic chromium. J. Trace Elem. Med. Biol. 1999, 13, 82–88. [Google Scholar] [CrossRef]
- Gilbert, E.R.; Li, H.; Emmerson, D.A.; WebbJr, K.E.; Wong, E.A. Developmental regulation of nutrient transporter and enzyme mRNA abundance in the small intestine of broilers. Poult. Sci. 2007, 86, 1739–1753. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F. Homologous physiological effects of phenformin and chromium picolinate. Med. Hypotheses 1993, 41, 316–324. [Google Scholar] [CrossRef]
- Campbell, W.W.; Polansky, M.M.; Bryden, N.A.; Soares, J.H.; Anderson, R.A. Exercise training and dietary chromium effects on glycogen, glycogen synthase, phosphorylase and total protein in rats. J. Nutr. 1989, 119, 653–660. [Google Scholar] [CrossRef]
- Edwards, M.R.; McMurtry, J.P.; Vasilatos-Younken, R. Relative insensitivity of avian skeletal muscle glycogen to nutritive status. Domest. Anim. Endocrinol. 1999, 16, 239–247. [Google Scholar] [CrossRef]
- Volek, J.S.; Silvestre, R.; Kirwan, J.P.; Sharman, M.J.; Judelson, D.A.; Spiering, B.A.; Vingren, J.L.; Maresh, C.M.; Vanheest, J.L.; Kraemer, W.J. Effects of chromium supplementation on glycogen synthesis after high-intensity exercise. Med. Sci. Sports Exerc. 2008, 38, 2102–2109. [Google Scholar] [CrossRef]
- Gardner, G.E.; Pethick, D.W.; Smith, G. Effect of chromium chelavite supplementation on the metabolism of glycogen and lipid in adult Merino sheep. Aust. J. Agric. Res. 1998, 49, 137–145. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Percentage |
|---|---|
| Corn | 58.50 |
| Soybean meal 44% | 25.00 |
| Sunflower meal | 3.50 |
| Canola meal | 8.00 |
| Vegetable oil | 1.50 |
| Dicalcium phosphate | 0.90 |
| Limestone | 1.51 |
| Common salt | 0.50 |
| DL-Methionine | 0.21 |
| L-Lysine HCl | 0.12 |
| Vitamin premix 1 | 0.13 |
| Micro mineral premix 2 | 0.13 |
| Total | 100.00 |
| Nutrient contents | |
| Crude protein | 20.72 |
| Metabolizable energy (MJ) | 12.2 |
| Calcium | 0.91 |
| Phosphorus | 0.61 |
| * Available P | 0.33 |
| Name of Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Annealing Temperature (°C) |
|---|---|---|---|
| SGLT-1 | GTCCTGGCAGTGGGAGTATG | AAGAGTGAAGCACCGATCGG | 61 |
| GLUT-2 | CACACTATGGGCGCATGCT | ATTGTCCCTGGAGGTGTTGGTG | 60 |
| β-Actin | ATGAAGCCCAGAGCAAAAGA | GGGGTGTTGAAGGTCTCAAA | 60 |
| Variables | Group A | Group B | Group C | Group D | Group E | SEM | p-Value | Linear | Quadratic | Cubic |
|---|---|---|---|---|---|---|---|---|---|---|
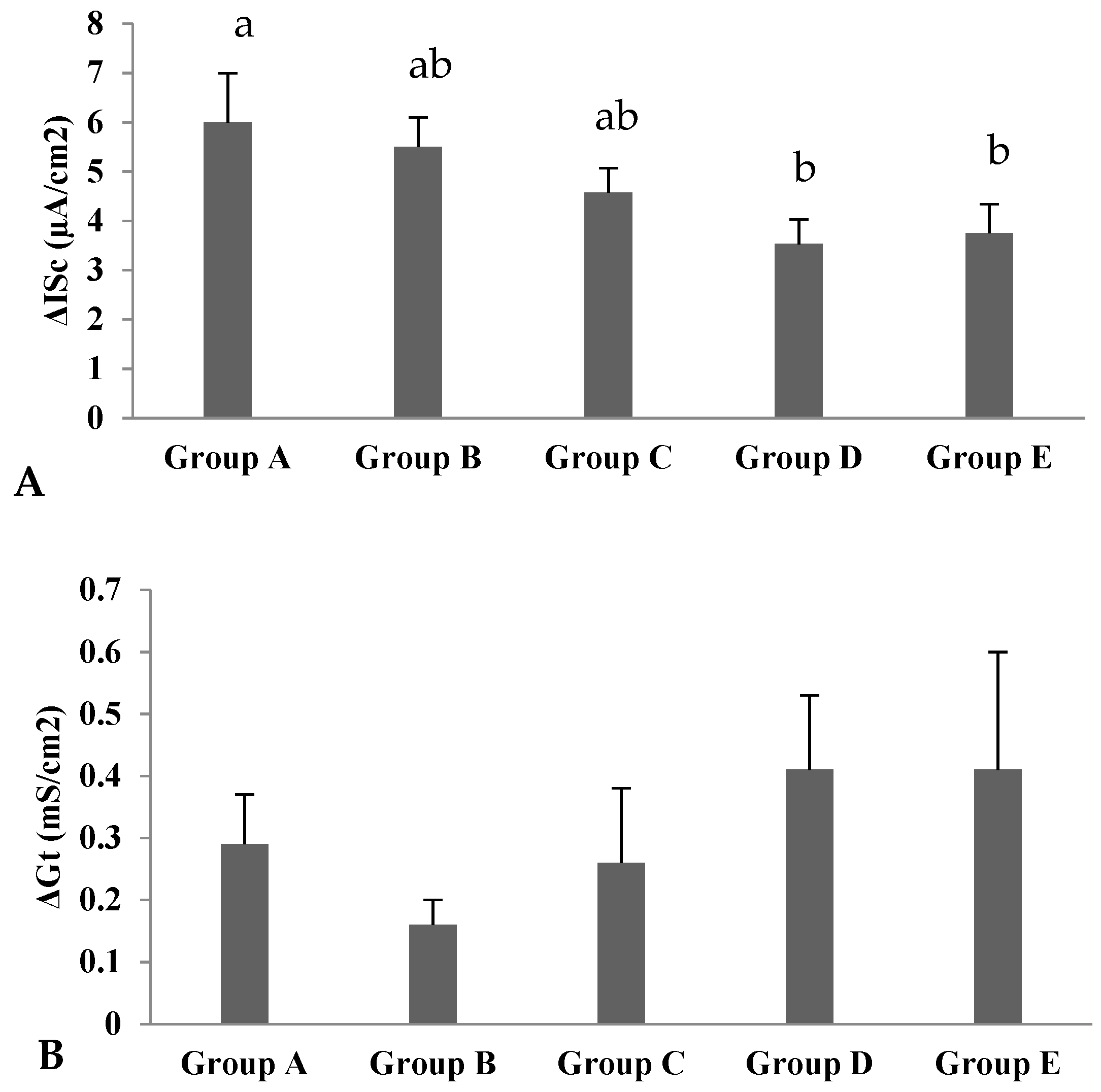
| ISci (µA/cm2) | 7.71 | 5.57 | 6.81 | 4.93 | 4.08 | 0.50 | 0.158 | 0.095 | 0.746 | 0.077 |
| Gti (mS/cm2) | 4.24 | 2.90 | 2.72 | 4.53 | 3.51 | 0.28 | 0.135 | 0.935 | 0.260 | 0.039 |
| Variables | Group A | Group B | Group C | Group D | Group E | SEM | p-Value | Linear | Quadratic | Cubic |
|---|---|---|---|---|---|---|---|---|---|---|
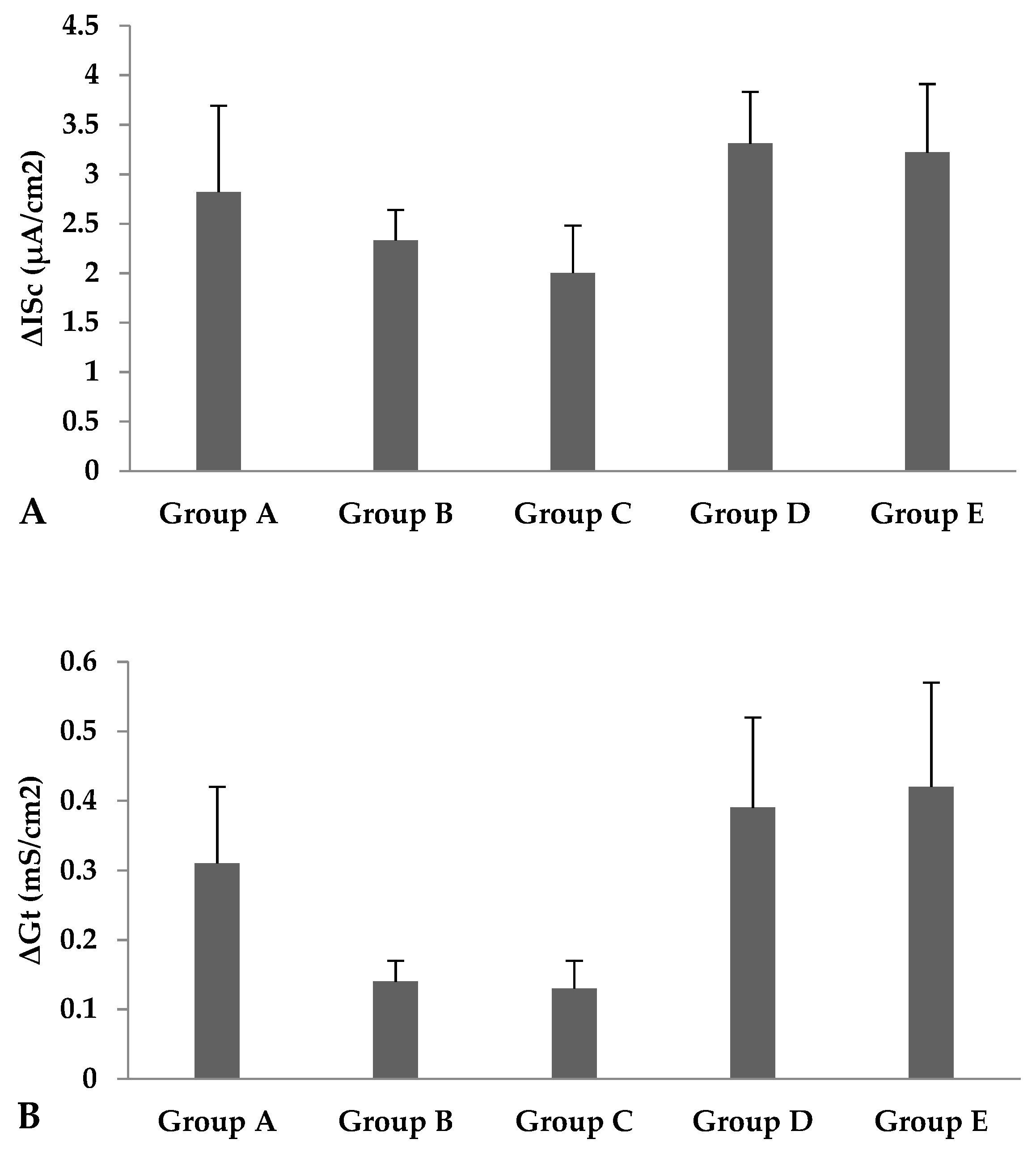
| ISci (µA/cm2) | 9.55 | 6.67 | 6.67 | 9.56 | 8.67 | 0.62 | 0.353 | 0.812 | 0.221 | 0.129 |
| Gti (mS/cm2) | 4.50 | 3.09 | 2.78 | 4.45 | 4.45 | 0.29 | 0.057 | 0.415 | 0.080 | 0.059 |
| Variables | Group A | Group B | Group C | Group D | Group E | SEM | p-Value | Linear | Quadratic | Cubic |
|---|---|---|---|---|---|---|---|---|---|---|
| Liver | 13.38 a | 13.89 a | 8.71 ab | 7.36 b | 6.71 b | 0.93 | 0.014 | 0.002 | 0.826 | 0.230 |
| Muscle | 3.37 | 3.75 | 3.11 | 3.24 | 3.05 | 0.17 | 0.756 | 0.382 | 0.807 | 0.592 |
| Blood Glucose | 252 a | 267 a | 229 a | 218 ab | 164 b | 14.5 | <0.001 | <0.001 | 0.046 | 0.796 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahir, S.K.; Yousaf, M.S.; Ahmad, S.; Shahzad, M.K.; Khan, A.F.; Raza, M.; Majeed, K.A.; Khalid, A.; Zaneb, H.; Rabbani, I.; et al. Effects of Chromium-Loaded Chitosan Nanoparticles on the Intestinal Electrophysiological Indices and Glucose Transporters in Broilers. Animals 2019, 9, 819. https://doi.org/10.3390/ani9100819
Tahir SK, Yousaf MS, Ahmad S, Shahzad MK, Khan AF, Raza M, Majeed KA, Khalid A, Zaneb H, Rabbani I, et al. Effects of Chromium-Loaded Chitosan Nanoparticles on the Intestinal Electrophysiological Indices and Glucose Transporters in Broilers. Animals. 2019; 9(10):819. https://doi.org/10.3390/ani9100819
Chicago/Turabian StyleTahir, Sajid Khan, Muhammad Shahbaz Yousaf, Sohrab Ahmad, Muhammad Khurram Shahzad, Ather Farooq Khan, Mohsin Raza, Khalid Abdul Majeed, Abia Khalid, Hafsa Zaneb, Imtiaz Rabbani, and et al. 2019. "Effects of Chromium-Loaded Chitosan Nanoparticles on the Intestinal Electrophysiological Indices and Glucose Transporters in Broilers" Animals 9, no. 10: 819. https://doi.org/10.3390/ani9100819