
Validation of Bos taurus SNPs for Milk Productivity of Sahiwal Breed (Bos indicus), Pakistan
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. DNA Measurement and Dilution
2.3. Primer Designing
2.4. PCR Protocol
2.5. Statistical Analysis
2.5.1. Linkage Disequilibrium
2.5.2. Haplotype Analysis and Association
3. Results
3.1. Single Locus-Based Analysis
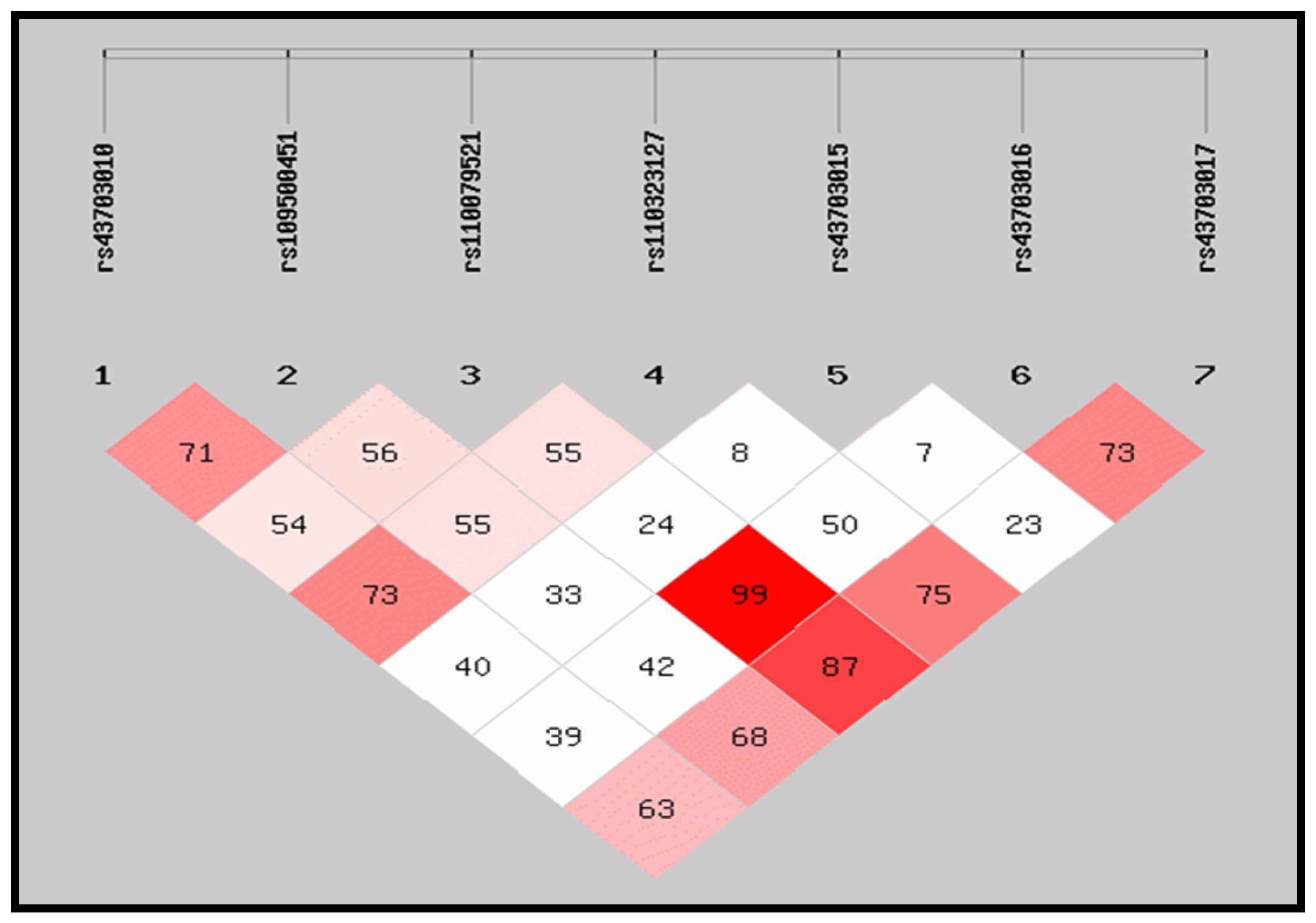
3.2. Pairwise Linkage Disequilibrium
3.3. Haplotype Frequencies and Association with Milk Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bilal, M.Q.; Suleman, M.; Raziq, A. Buffalo: Black gold of Pakistan. Livest. Res. Rural. Dev. 2006, 18, 140–151. [Google Scholar]
- Rehman, A.; Jingdong, L.; Chandio, A.A.; Hussain, I. Livestock production and population census in Pakistan: Determining their relationship with agricultural GDP using econometric analysis. Inf. Process. Agric. 2017, 4, 168–177. [Google Scholar] [CrossRef]
- Singh, P.K.; Kumar, D.; Varma, S.K. Genetic studies and development of prediction equations in Jersey × Sahiwal and Holstein-Friesian × Sahiwal half breds. Asian-Australas. J. Anim. Sci. 2005, 18, 179–184. [Google Scholar] [CrossRef]
- Illa, S.K.; Mukherjee, S.; Nath, S.; Mukherjee, A. Genome-wide scanning for signatures of selection revealed the putative genomic regions and candidate genes controlling milk composition and coat color traits in Sahiwal cattle. Front. Genet. 2021, 12, 699422. [Google Scholar] [CrossRef] [PubMed]
- Worku, D.; Gowane, G.; Alex, R.; Joshi, P.; Verma, A. Inputs for optimizing selection platform for milk production traits of dairy Sahiwal cattle. PLoS ONE 2022, 17, e0267800. [Google Scholar] [CrossRef]
- VanRaden, P.M. Symposium review: How to implement genomic selection. J. Dairy Sci. 2020, 103, 5291–5301. [Google Scholar] [CrossRef] [PubMed]
- Garkovenko, A.V.; Radchenko, V.V.; Ilnitskaya, E.V.; Koshchaev, A.G.; Shchukina, I.V.; Bakharev, A.A.; Sukhanova, S.F. Polymorphism of cattle microsatellite complexes. J. Pharm. Sci. Res. 2018, 10, 1545–1551. [Google Scholar]
- Asylbekovich, B.D.; Kurmanbaiuly, A.S.; Beskempirovich, S.N.; Grigoryevich, S.V.; Erbosynovich, C.A. Productivity and estimated breeding value of the dairy cattle gene pool in the Republic of Kazakhstan. Научныйжурнал «Вестник НАН РК» 2019, 1, 39–53. [Google Scholar]
- Raschia, M.A.; Nani, J.P.; Maizon, D.O.; Beribe, M.J.; Amadio, A.F.; Poli, M.A. Single nucleotide polymorphisms in candidate genes associated with milk yield in Argentinean Holstein and Holstein × Jersey cows. J. Anim. Sci. Technol. 2018, 60, 31. [Google Scholar] [CrossRef]
- Yao, S.; Liu, Y.; Liu, X.; Liu, G. Effects of SNPs in AANAT and ASMT Genes on Milk and Peripheral Blood Melatonin Concentrations in Holstein Cows (Bos taurus). Genes 2022, 13, 1196. [Google Scholar] [CrossRef]
- Asmarasari, S.A.; Sumantri, C.; Gunawan, A.; Taufik, E.; Anggraeni, A.; Hapsari AA, R.; Dewantoro, B. Kappa casein (CSN3) gene polymorphism and its effect on cumulative milk yields of Holstein Friesian dairy cattle. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; Volume 902, p. 012047. [Google Scholar]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited review: Milk protein polymorphisms in cattle: Effect on animal breeding and human nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef] [PubMed]
- Bech, A.M.; Kristiansen, K.R. Milk protein polymorphism in Danish dairy cattle and the influence of genetic variants on milk yield. J. Dairy Res. 1990, 57, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Mohan, G.; Kumar, A.; Khan, S.H.; Kumar, N.A.; Kapila, S.; Lathwal, S.S.; Niranjan, S.K. Casein (CSN) gene variants and parity affect the milk protein traits in crossbred (Bos taurus x Bos indicus) cows in sub-tropical climate. Trop. Anim. Health Prod. 2021, 53, 289. [Google Scholar] [CrossRef] [PubMed]
- Miranda, G.; Bianchi, L.; Krupova, Z.; Trossat, P.; Martin, P. An improved LC–MS method to profile molecular diversity and quantify the six main bovine milk proteins, including genetic and splicing variants as well as post-translationally modified isoforms. Food Chem. 2020, 5, 100080. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Korkuć, P.; Arends, D.; Brockmann, G.A. DNA sequence variants and protein haplotypes of casein genes in German Black Pied Cattle (DSN). Front. Genet. 2019, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Shah, T.; Sabara, P.; Bhatia, D.; Panchal, K.; Italiya, J.; Rank, D.N. Understanding functional implication of β-casein gene variants in four cattle breeds characterized using AmpliSeq approach. 3 Biotech 2020, 10, 414. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Nassiry, M.R.; Mosafer, J.; Mohammadabadi, M.R.; Sulimova, G.E. Distribution of BoLA-DRB3 allelic frequencies and identification of a new allele in the Iranian cattle breed Sistani (Bos indicus). Russ. J. Genet. 2009, 45, 198–202. [Google Scholar] [CrossRef]
- Mohammadabadi, M.R. Tissue-specific mRNA expression profile of ESR2 gene in goat. Agric. Biotechnol. J. 2021, 12, 169–184. [Google Scholar]
- Mohammadabadi, M.R.; Nikbakhti, M.; Mirzaee, H.R.; Shandi, A.; Saghi, D.A.; Romanov, M.N.; Moiseyeva, I.G. Genetic variability in three native Iranian chicken populations of the Khorasan province based on microsatellite markers. Russ. J. Genet. 2010, 46, 505–509. [Google Scholar] [CrossRef]
- Mohammadabadi, M.R.; Jafari, A.H.D.; Bordbar, F. Molecular analysis of CIB4 gene and protein in Kermani sheep. Brazil. J. Med. Biol. Res. 2017, 50, e6177. [Google Scholar] [CrossRef]
- Mohammadabadi, M.; Bordbar, F.; Jensen, J.; Du, M.; Guo, W. Key genes regulating skeletal muscle development and growth in farm animals. Animals 2021, 11, 835. [Google Scholar] [CrossRef] [PubMed]
- Mohammadabadi, M.R.; Soflaei, M.; Mostafavi, H.; Honarmand, M. Using PCR for early diagnosis of bovine leukemia virus infection in some native cattle. Genet. Mol. Res. 2011, 10, 2658–2663. [Google Scholar] [CrossRef] [PubMed]
- Gooki, F.G.; Mohammadabadi, M.; Fozi, M.A.; Soflaei, M. Association of Biometric Traits with Growth Hormone Gene Diversity in Raini Cashmere Goats. Walailak J. Sci. Technol. (WJST) 2019, 16, 499–508. [Google Scholar] [CrossRef]
- Gholamhoseini Gooki, F.; Mohammadabadi, M.R.; Asadi Fozi, M. Polymorphism of the growth hormone gene and its effect on production and reproduction traits in goat. Iran. J. Appl. Anim. Sci. 2018, 8, 653–659. [Google Scholar]
- Pokorska, J.; Kułaj, D.; Dusza, M.; Żychlińska-Buczek, J.; Makulska, J. New rapid method of DNA isolation from milk somatic cells. Anim. Biotechnol. 2016, 27, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Darawi, M.N.; Ai-Vyrn, C.; Ramasamy, K.; Hua, P.P.J.; Pin, T.M.; Kamaruzzaman, S.B.; Majeed, A.B.A. Allele-specific polymerase chain reaction for the detection of Alzheimer’s disease-related single nucleotide polymorphisms. BMC Med. Genet. 2013, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast/ (accessed on 29 March 2019).
- Yong, Y.O.N.G.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97. [Google Scholar]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed]
- Khatkar, M.S.; Thomson, P.C.; Tammen, I.; Raadsma, H.W. Quantitative trait loci mapping in dairy cattle: Review and meta-analysis. Genet. Sel. Evol. 2004, 36, 163–190. [Google Scholar] [CrossRef]
- Nilsen, H.; Olsen, H.G.; Hayes, B.; Sehested, E.; Svendsen, M.; Nome, T.; Meuwissen, T.; Lien, S. Casein haplotypes and their association with milk production traits in Norwegian Red cattle. Genet. Sel. Evol. 2009, 41, 21. [Google Scholar] [CrossRef]
- Asim, M.; Saif-ur Rehman, M.; Hassan, F.U.; Awan, F.S. Genetic variants of CSN1S1, CSN2, CSN3, and BLG genes and their association with dairy production traits in Sahiwal cattle and Nili-Ravi buffaloes. Anim. Biotechnol. 2022, 34, 2951–2962. [Google Scholar] [CrossRef]
- Morammazi, S.; Masoudi, A.A.; Torshizi, R.V.; Pakdel, A. Differential expression of the alpha S1 casein and beta-lactoglobulin genes in different physiological stages of the Adani goats mammary glands. Iran. J. Biotechnol. 2016, 14, 278. [Google Scholar] [CrossRef]
- Huang, W.; Peñagaricano, F.; Ahmad, K.R.; Lucey, J.A.; Weigel, K.A.; Khatib, H. Association between milk protein gene variants and protein composition traits in dairy cattle. J. Dairy Sci. 2012, 95, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhou, J.; Chen, C.J.; Zhang, J.; Wen, W.; Tian, J.; Zhang, Z.; Gu, Y. GWAS-based identification of new loci for milk yield, fat, and protein in Holstein cattle. Animals 2020, 10, 2048. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, C.; Cai, W.; Liu, S.; Yin, H.; Shi, S.; Zhang, S. Genome-wide association study for milk protein composition traits in a Chinese Holstein population using a single-step approach. Front. Genet. 2019, 10, 72. [Google Scholar] [CrossRef]
- Selvaggi, M.; Laudadio, V.; Dario, C.; Tufarelli, V. Investigating the genetic polymorphism of sheep milk proteins: A useful tool for dairy production. J. Sci. Food Agric. 2014, 94, 3090–3099. [Google Scholar] [CrossRef]
- Ganguly, I.; Kumar, S.; Gaur, G.K.; Singh, U.; Kumar, A.; Kumar, S.; Sharma, A. Status of β-casein (CSN2) polymorphism in Frieswal (HF X Sahiwal Crossbred) cattle. Int. J. Biotechnol. Bioeng. Res. 2013, 4, 249–256. [Google Scholar]
- Comin, A.; Cassandro, M.; Chessa, S.; Ojala, M.; Dal Zotto, R.; De Marchi, M.; Carnier, P.; Gallo, L.; Pagnacco, G.; Bittante, G. Effects of composite β-and κ-casein genotypes on milk coagulation, quality, and yield traits in Italian Holstein cows. J. Dairy Sci. 2008, 91, 4022–4027. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, Z.; Qiu, L.; Zhang, Y.; Miao, Y. Polymorphisms of the kappa casein (CSN3) gene and inference of its variants in water buffalo (Bubalus bubalis). Arch. Anim. Breed. 2019, 62, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, J.; Matejicek, A.; Jandurová, O.M.; Sorensen, P.; Nemcova, E.; Stipkova, M.; Frelich, J. Milk protein genes CSN1S1, CSN2, CSN3, LGB and their relation to genetic values of milk production parameters in Czech Fleckvieh. Czech J. Anim. Sci. 2006, 51, 241. [Google Scholar] [CrossRef]
- Pizarro, M.G.; Landi, V.; Navas, F.J.; León, J.M.; Martínez, A.; Fernández, J.; Delgado, J.V. Nonparametric analysis of casein complex genes’ epistasis and their effects on phenotypic expression of milk yield and composition in Murciano-Granadina goats. J. Dairy Sci. 2020, 103, 8274–8291. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.animalgenome.org/qtldb/export/ksui8gfhot6/cattle_qtldata.txt (accessed on 1 April 2019).
- Pedrosa, V.B.; Schenkel, F.S.; Chen, S.Y.; Oliveira, H.R.; Casey, T.M.; Melka, M.G.; Brito, L.F. Genomewide association analyses of lactation persistency and milk production traits in Holstein cattle based on imputed whole-genome sequence data. Genes 2021, 12, 1830. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, M.; Kopuzlu, S.; Topal, M.; Bilgin, O.C. Relationships between milk protein polymorphisms and production traits in cattle: A systematic review and meta-analysis. Arch. Anim. Breed. 2018, 61, 197–206. [Google Scholar] [CrossRef]
- Kruchinin, A.G.; Illarionova, E.E.; Galstyan, A.G.; Turovskaya, S.N.; Bigaeva, A.V.; Bolshakova, E.I.; Strizhko, M.N. Effect of CSN3 Gene Polymorphism on the Formation of Milk Gels Induced by Physical, Chemical, and Biotechnological Factors. Foods 2023, 12, 1767. [Google Scholar] [CrossRef] [PubMed]
- Mancini, G.; Nicolazzi, E.L.; Valentini, A.; Chillemi, G.; AjmoneMarsan, P.; Santus, E.; Pariset, L. Association between single nucleotide polymorphisms (SNPs) and milk production traits in Italian Brown cattle. Livest. Sci. 2013, 157, 93–99. [Google Scholar] [CrossRef]
- Heck, J.M.L.; Schennink, A.; van Valenberg, H.J.F.; Bovenhuis, H.; Visker, M.H.P.W.; van Arendonk, J.A.M.; van Hooijdonk, A.C.M. Effects of milk protein variants on the protein composition of bovine milk. J. Dairy Sci. 2009, 92, 1192–1202. [Google Scholar] [CrossRef]
- Deb, R.; Singh, U.; Kumar, S.; Singh, R.; Sengar, G.; Sharma, A. Genetic polymorphism and association of kappa-casein gene with milk production traits among Frieswal (HF × Sahiwal) cross breed of Indian origin. Iran. J. Vet. Res. 2014, 15, 406. [Google Scholar] [PubMed]
- Alim, M.A.; Dong, T.; Xie, Y.; Wu, X.P.; Zhang, Y.; Zhang, S.; Sun, D.X. Effect of polymorphisms in the CSN3 (κ-casein) gene on milk production traits in Chinese Holstein Cattle. Mol. Biol. Rep. 2014, 41, 7585–7593. [Google Scholar] [CrossRef]
- Boettcher, P.J.; Caroli, A.; Stella, A.; Chessa, S.; Budelli, E.; Canavesi, F.; Ghiroldi, S.; Pagnacco, G. Effects of casein haplotypes on milk production traits in Italian Holstein and Brown Swiss cattle. J. Dairy Sci. 2004, 87, 4311–4317. [Google Scholar] [CrossRef]
- Penasa, M.; Cassandro, M.; Pretto, D.; De Marchi, M.; Comin, A.; Chessa, S.; Zotto, R.D.; Bittante, G. Short communication: Influence of composite casein genotypes on additive genetic variation of milk production traits and coagulation properties in Holstein-Friesian cows. J. Dairy Sci. 2010, 93, 3346–3349. [Google Scholar] [CrossRef]
- Ikonen, T.; Bovenhuis, H.; Ojala, M.; Ruottinen, O.; Georges, M. Associations between casein haplotypes and first lactation milk production traits in Finnish Ayrshire cows. J. Dairy Sci. 2001, 84, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Moioli, B.; D’Andrea, M.; Pilla, F.J.S.R.R. Candidate genes affecting sheep and goat milk quality. Small Rumin. Res. 2007, 68, 179–192. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sr# | SNP Ref No | Gene | 5 > 3 | |
|---|---|---|---|---|
| 1 | rs110672723 | CSN2 | Fwd | GCCTTTTGGCAAGAAA |
| Rev | CCACATGACTCATTTCACATCT | |||
| Rev | CCACATGACTCATTTCACATCG | |||
| 2 | Rs43703011 | CSN2 | Fwd | GTTTTGTGGGAGGCTGTTAG |
| Fwd | GTTTTGTGGGAGGCTGTTAT | |||
| Rev | TCTTTCCAGGATGAACTCCAG | |||
| 3 | rs43703010 | CSN1S1 | Fwd | ATTACGTTCCACTAGGCAC |
| Rev | CAGTGGCATAGTAGTCTTTT | |||
| Re | CAGTGGCATAGTAGTCTTTC | |||
| 4 | rs109500451 | CSN1S2 | Fwd | GGTATAAAACTGATAG |
| Rev | GGCTGATACGTTAAGAAATC | |||
| Rev | GGCTGATACGTTAAGAAATT | |||
| 5 | rs110079521 | CSN3 | Fwd | AATCAACCAGATGGATGATA |
| Fwd | AATCAACCAGATGGATGATC | |||
| Rev | GTTTTGCCATACATCAACAT | |||
| 6 | rs110323127 | CSN3 | Fwd | CAAGAAGTGGAAGGAAG |
| Fwd | CAAGAAGTGGAAGGAAA | |||
| Rev | CAGTATTTGATGTTAAG | |||
| 7 | rs43703015 | CSN3 | Fwd | CCTACAAGTACACCTACCAT |
| Fwd | CCTACAAGTACACCTACCAC | |||
| Rev | CTGTGTTGATCTCAGGTG | |||
| 8 | rs43703016 | CSN3 | Fwd | TGTAGCTACTCTAGAAGC |
| Fwd | TGTAGCTACTCTAGAAGA | |||
| Rev | CCTTAGAGTATTTAGACCGC | |||
| 9 | rs43703017 | CSN3 | Fwd | GCTTCTCCAGAAGTTATTGAGA |
| Fwd | GCTTCTCCAGAAGTTATTGAGG | |||
| Rev | ACCTGCGTTGTCTTCTTTG |
| SNP No | Allele | Chromosome | Gene |
|---|---|---|---|
| rs43703010 | A/G | 6 | CSN1 |
| rs109500451 | A/G | 6 | CSN1S2 |
| rs110672723 | A/C | 6 | CSN2 |
| Rs43703011 | G/T | 6 | CSN2 |
| rs110079521 | C/A | 6 | CSN3 |
| rs110323127 | A/G | 6 | CSN3 |
| rs43703015 | C/T | 6 | CSN3 |
| rs43703016 | A/C | 6 | CSN3 |
| rs43703017 | A/G | 6 | CSN3 |
| Gene | Position | Locus | Genotype | N | Frequency | Allele | Frequency | Hardy-Weienberg Equilibrium X2 Test |
|---|---|---|---|---|---|---|---|---|
| CSN1 | Fwd | rs43703010 | AA AG GG | 37 13 30 | 0.38 0.16 0.46 | A G | 0.54 0.46 | p < 0.05 |
| CSN1S2 | Fwd | Rs109500451 | AA AG GG | 18 39 23 | 0.22 0.49 0.29 | G A | 0.53 0.47 | p > 0.05 |
| CSn3 | Fwd | Rs110079521 | AA AC CC | 32 28 20 | 0.4 0.35 0.25 | A C | 0.57 0.42 | p < 0.05 |
| CSN3 | Fwd | Rs110323127 | AA GA GG | 7 24 46 | 0.09 0.34 0.57 | G A | 0.74 0.26 | p > 0.05 |
| CSN3 | Fwd | Rs43703015 | CC TC TT | 6 9 55 | 0.08 0.24 0.69 | T C | 0.81 0.19 | p > 0.05 |
| CSN3 | Fwd | Rs43703016 | AA AC CC | 26 34 20 | 0.34 0.42 0.25 | A C | 0.54 0.45 | p > 0.05 |
| CSN3 | Fwd | Rs43703017 | AA GA GG | 17 29 34 | 0.21 0.36 0.42 | G A | 0.61 0.39 | p > 0.05 |
| Gene | Locus | Genotype | Milk Yield | p Value |
|---|---|---|---|---|
| CSN1 | rs43703010 | AA | 8.03 ± 0.18 | p < 0.05 |
| AG | 9.85 ± 0.13 | |||
| GG | 10.09 ± 0.1 | |||
| CSN1S2 | rs109500451 | AA | 7.55 ± 0.27 | p < 0.05 |
| AG | 9.21 ± 0.14 | |||
| GG | 10.14 ± 0.17 | |||
| CSN3 | rs110079521 | AA | 10.33 ± 0.05 | p < 0.05 |
| AC | 8.77 ± 0.11 | |||
| CC | 7.6 ± 0.27 | |||
| CSN3 | rs110323127 | AA | 9.14 ± 0.43 | p < 0.05 |
| GA | 7.61 ± 0.17 | |||
| GG | 9.97 ± 0.08 | |||
| CSN3 | rs43703015 | CC | 10.72 ± 0.06 | p < 0.05 |
| TC | 8.43 ± 0.45 | |||
| TT | 9.16 ± 0.12 | |||
| CSN3 | rs43703016 | AA | 10.29 ± 0.06 | p < 0.05 |
| AC | 9.07 ± 0.14 | |||
| CC | 7.6 ± 0.27 | |||
| CSN3 | rs43703017 | AA | 10.39 ± 0.06 | p < 0.05 |
| GA | 9.86 ± 0.09 | |||
| GG | 7.81 ± 0.15 |
| Haplotype | SNP1 | SNP2 | SNP3 | SNP4 | SNP5 | SNP6 | SNP7 | Frequency |
|---|---|---|---|---|---|---|---|---|
| GGAGTAA | G | G | A | G | T | A | A | 0.2006 |
| AACATCG | A | A | C | A | T | C | G | 0.1705 |
| AACGCCG | A | A | C | G | C | C | G | 0.0625 |
| AAAGTAA | A | A | A | G | T | A | A | 0.0619 |
| AAAGTAG | A | A | A | G | T | A | G | 0.0491 |
| AACGTCG | A | A | C | G | T | C | G | 0.0482 |
| AGAGTAG | A | G | A | G | T | A | G | 0.0472 |
| GGAGCAA | G | G | A | G | C | A | A | 0.0366 |
| GACGTCG | G | A | C | G | T | C | G | 0.0362 |
| AGCGTCG | A | G | C | G | T | C | G | 0.0333 |
| GGAGCCA | G | G | A | G | C | C | A | 0.0314 |
| GGCGTCG | G | G | C | G | T | C | G | 0.0313 |
| GGAGCAG | G | G | A | G | C | A | G | 0.0292 |
| GGAACAA | G | G | A | A | C | A | A | 0.025 |
| GGCGTCA | G | G | C | G | T | C | A | 0.0218 |
| AGCATCG | A | G | C | A | T | C | G | 0.0213 |
| AAAATAG | A | A | A | A | T | A | G | 0.0193 |
| GAAGTAG | G | A | A | G | T | A | G | 0.0181 |
| AGAGTAA | A | G | A | G | T | A | A | 0.0166 |
| AGAATAG | A | G | A | A | T | A | G | 0.0139 |
| GGAGTAG | G | G | A | G | T | A | G | 0.0109 |
| GGAATAG | G | G | A | A | T | A | G | 0.0062 |
| GGAGCCG | G | G | A | G | C | C | G | 0.0061 |
| GAAGCAG | G | A | A | G | C | A | G | 0.0029 |
| AACACCG | A | A | C | A | C | C | G | 0 |
| AGCGCCG | A | G | C | G | C | C | G | 0 |
| GGAGTAA | G | G | A | G | T | A | A | 0 |
| GACGTCA | G | A | C | G | T | C | A | 0 |
| Haplotype | SNP1 | SNP2 | SNP3 | SNP4 | SNP5 | SNP6 | SNP7 | Frequency | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| GGAGTAA | G | G | A | G | T | A | A | 0.1964 | <0.0001 |
| AACATCG | A | A | C | A | T | C | G | 0.1638 | <0.0001 |
| AACGCCG | A | A | C | G | C | C | G | 0.0625 | <0.0001 |
| AACGTCG | A | A | C | G | T | C | G | 0.0507 | <0.0001 |
| GACGTCG | G | A | C | G | T | C | G | 0.0383 | <0.0001 |
| GGAGCAA | G | G | A | G | C | A | A | 0.0382 | 0.01 |
| GGAGCCA | G | G | A | G | C | C | A | 0.0375 | 0.1 |
| GGCGTCG | G | G | C | G | T | C | G | 0.033 | <0.0001 |
| AGCATCG | A | G | C | A | T | C | G | 0.0328 | <0.0001 |
| GGAACAA | G | G | A | A | C | A | A | 0.025 | 0.0064 |
| GGCGTCA | G | G | C | G | T | C | A | 0.0188 | 0.0062 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younis, A.; Hussain, I.; Ahmad, S.N.; Shah, A.; Inayat, I.; Kanwal, M.A.; Suleman, S.; Kamran, M.A.; Matloob, S.; Ahmad, K.R. Validation of Bos taurus SNPs for Milk Productivity of Sahiwal Breed (Bos indicus), Pakistan. Animals 2024, 14, 1306. https://doi.org/10.3390/ani14091306
Younis A, Hussain I, Ahmad SN, Shah A, Inayat I, Kanwal MA, Suleman S, Kamran MA, Matloob S, Ahmad KR. Validation of Bos taurus SNPs for Milk Productivity of Sahiwal Breed (Bos indicus), Pakistan. Animals. 2024; 14(9):1306. https://doi.org/10.3390/ani14091306
Chicago/Turabian StyleYounis, Asma, Imtiaz Hussain, Syeda Nadia Ahmad, Amin Shah, Iram Inayat, Muhammad Ali Kanwal, Sadia Suleman, Muhammad Atif Kamran, Saima Matloob, and Khawaja Raees Ahmad. 2024. "Validation of Bos taurus SNPs for Milk Productivity of Sahiwal Breed (Bos indicus), Pakistan" Animals 14, no. 9: 1306. https://doi.org/10.3390/ani14091306