

Effects of Methionine Supplementation Levels in Normal or Reduced Protein Diets on the Body Composition and Femur Bone Characteristics of Broilers Challenged with Coccidia
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Bird Husbandry
2.2. Dual-Energy X-ray Absorptiometry for Body Composition Analysis
2.3. Microtomography Scanning for Microstructural Analysis of the Femur Bone
2.4. Statistical Analysis
3. Results
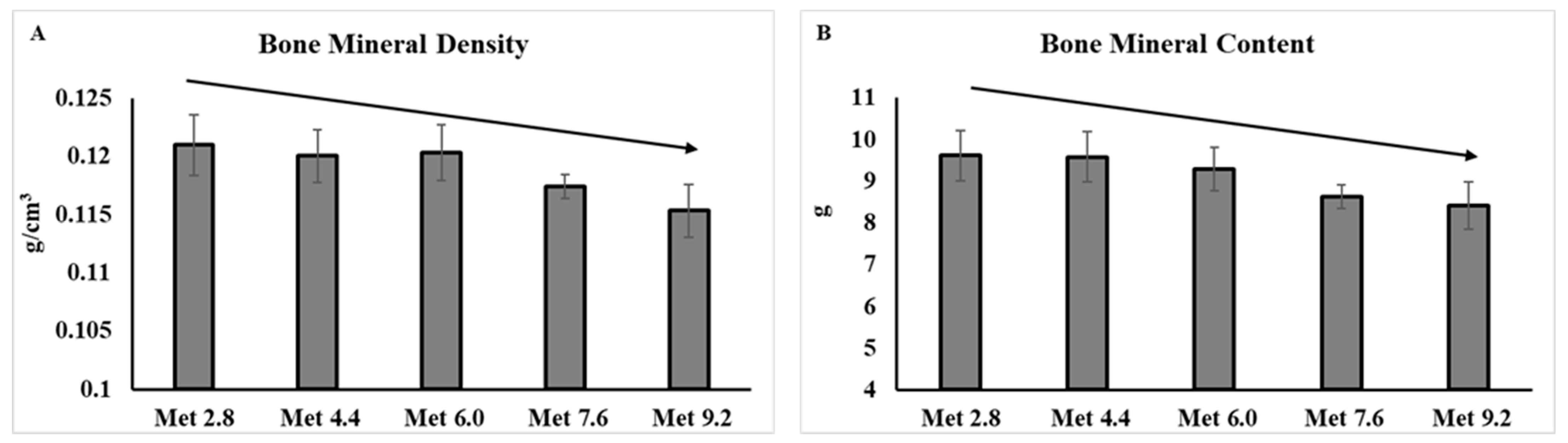
3.1. Body Composition Analyzed by Dual-Energy X-ray Absorptiometry
3.2. Femur Bone Microstructure Analyzed by Microtomography
3.2.1. Metaphysis Cortical Bone
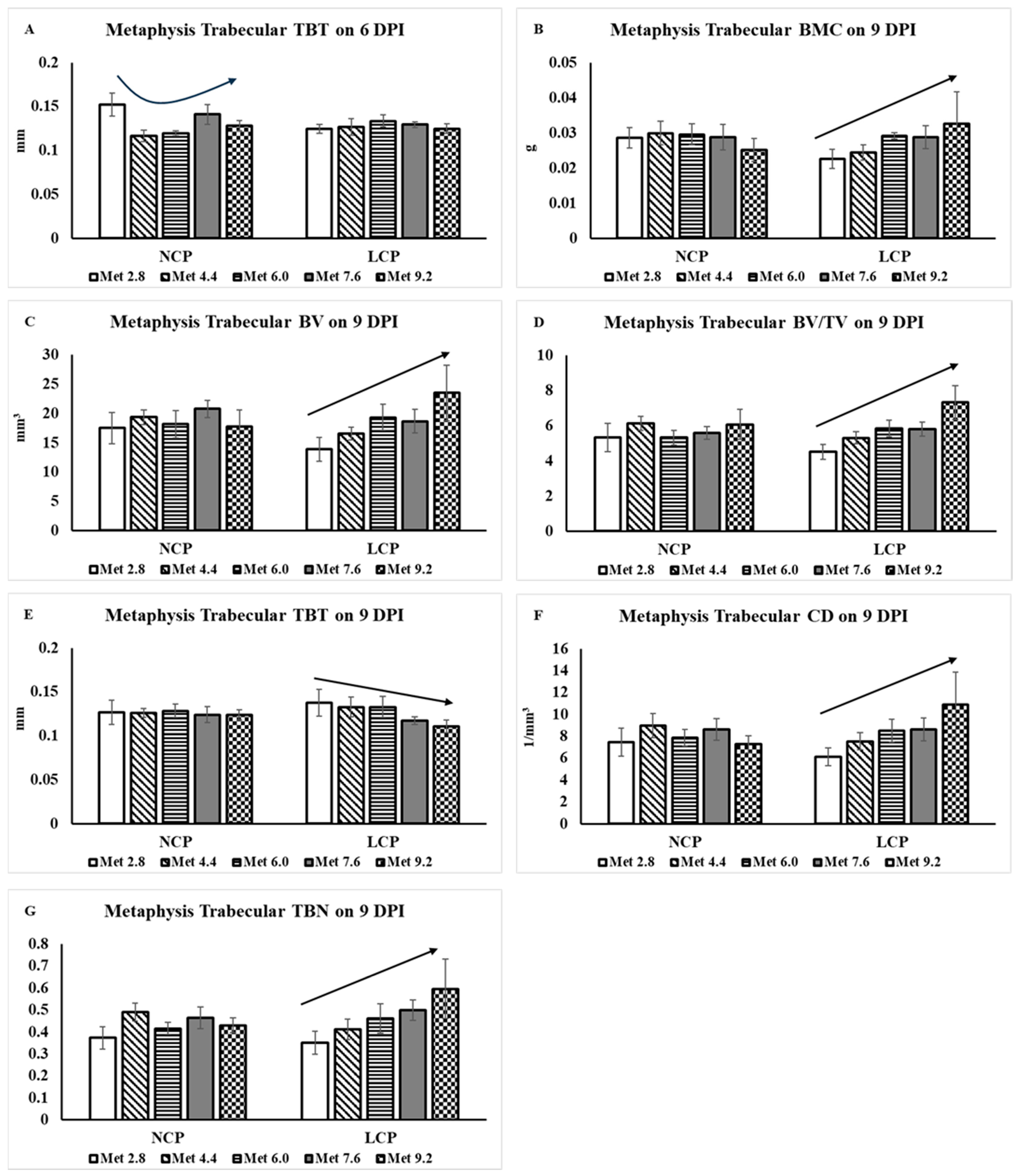
3.2.2. Metaphysis Trabecular Bone
3.2.3. Diaphysis Cortical Bone
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef]
- Liu, G.; Ajao, A.M.; Shanmugasundaram, R.; Taylor, J.; Ball, E.; Applegate, T.J.; Selvaraj, R.; Kyriazakis, I.; Olukosi, O.A.; Kim, W.K. The effects of arginine and branched-chain amino acid supplementation to reduced-protein diet on intestinal health, cecal short-chain fatty acid profiles, and immune response in broiler chickens challenged with Eimeria spp. Poult. Sci. 2023, 102, 102773. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Liu, G.; Choi, J.; Yadav, S.; Wei, F.; Kim, W.K. Effects of levels of methionine supplementations in forms of L- or DL-methionine on the performance, intestinal development, immune response, and antioxidant system in broilers challenged with Eimeria spp. Poult. Sci. 2023, 102, 102586. [Google Scholar] [CrossRef]
- Sharma, M.K.; Liu, G.; White, D.L.; Kim, W.K. Graded levels of Eimeria infection linearly reduced the growth performance, altered the intestinal health, and delayed the onset of egg production of Hy-Line W-36 laying hens when infected at the prelay stage. Poult. Sci. 2024, 103, 103174. [Google Scholar] [CrossRef]
- Taylor, J.; Sakkas, P.; Kyriazakis, I. Starving for nutrients: Anorexia during infection with parasites in broilers is affected by diet composition. Poult. Sci. 2022, 101, 101535. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, P.; Oikeh, I.; Blake, D.P.; Smith, S.; Kyriazakis, I. Dietary vitamin D improves performance and bone mineralisation, but increases parasite replication and compromises gut health in Eimeria-infected broilers. Br. J. Nutr. 2019, 122, 676–688. [Google Scholar] [CrossRef]
- Tompkins, Y.H.; Choi, J.; Teng, P.-Y.; Yamada, M.; Sugiyama, T.; Kim, W.K. Reduced bone formation and increased bone resorption drive bone loss in Eimeria infected broilers. Sci. Rep. 2023, 13, 616. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, H.M. Antibiotic-free poultry production: Is it sustainable? J. Appl. Poult. Res. 2015, 24, 91–97. [Google Scholar] [CrossRef]
- Cervantes, H.; McDougald, L. Raising broiler chickens without ionophore anticoccidials. J. Appl. Poult. Res. 2023, 32, 100347. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Iqbal, Z.; Khan, A.; Sindhu, Z.-U.-D.; Khan, J.A.; Khan, M.N.; Raza, A. Options for integrated strategies for the control of avian coccidiosis. Int. J. Agric. Biol. 2012, 14, 1014–1020. [Google Scholar]
- Dozier, W.; Kidd, M.T.; Corzo, A. Dietary Amino Acid Responses of Broiler Chickens1. J. Appl. Poult. Res. 2008, 17, 157–167. [Google Scholar] [CrossRef]
- Liu, G.; Sharma, M.K.; Tompkins, Y.H.; Teng, P.-Y.; Kim, W.K. Impacts of varying methionine to cysteine supplementation ratios on growth performance, oxidative status, intestinal health, and gene expression of immune response and methionine metabolism in broilers under Eimeria spp. challenge. Poult. Sci. 2024, 103, 103300. [Google Scholar] [CrossRef]
- Castro, F.L.S.; Teng, P.Y.; Yadav, S.; Gould, R.L.; Craig, S.; Pazdro, R.; Kim, W.K. The effects of L-Arginine supplementation on growth performance and intestinal health of broiler chickens challenged with Eimeria spp. Poult. Sci. 2020, 99, 5844–5857. [Google Scholar] [CrossRef] [PubMed]
- Khatlab, A.d.S.; Del Vesco, A.P.; de Oliveira Neto, A.R.; Fernandes, R.P.M.; Gasparino, E. Dietary supplementation with free methionine or methionine dipeptide mitigates intestinal oxidative stress induced by Eimeria spp. challenge in broiler chickens. J. Anim. Sci. Biotechnol. 2019, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Kim, W.K. The Functional Roles of Methionine and Arginine in Intestinal and Bone Health of Poultry: Review. Animals 2023, 13, 2949. [Google Scholar] [CrossRef]
- Lee, J.T.; Rochell, S.J.; Kriseldi, R.; Kim, W.K.; Mitchell, R.D. Functional properties of amino acids: Improve health status and sustainability. Poult. Sci. 2023, 102, 102288. [Google Scholar] [CrossRef] [PubMed]
- Teng, P.-Y.; Choi, J.; Yadav, S.; Tompkins, Y.H.; Kim, W.K. Effects of low-crude protein diets supplemented with arginine, glutamine, threonine, and methionine on regulating nutrient absorption, intestinal health, and growth performance of Eimeria-infected chickens. Poult. Sci. 2021, 100, 101427. [Google Scholar] [CrossRef] [PubMed]
- Lugata, J.K.; Ortega, A.D.S.V.; Szabó, C. The Role of Methionine Supplementation on Oxidative Stress and Antioxidant Status of Poultry-A Review. Agriculture 2022, 12, 1701. [Google Scholar] [CrossRef]
- Tompkins, Y.H.; Liu, G.; Kim, W.K. Impact of exogenous hydrogen peroxide on osteogenic differentiation of broiler chicken compact bones derived mesenchymal stem cells. Front. Physiol. 2023, 14, 1124355. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, Y.; Teng, P.; Pazdro, R.; Kim, W. Long bone mineral loss, bone microstructural changes and oxidative stress after Eimeria challenge in broilers. Front. Physiol. 2022, 13, 945740. [Google Scholar] [CrossRef]
- Sinclair, L.V.; Howden, A.J.M.; Brenes, A.; Spinelli, L.; Hukelmann, J.L.; Macintyre, A.N.; Liu, X.; Thomson, S.; Taylor, P.M.; Rathmell, J.C.; et al. Antigen receptor control of methionine metabolism in T cells. eLife 2019, 8, e44210. [Google Scholar] [CrossRef]
- Elias, R.J.; McClements, D.J.; Decker, E.A. Antioxidant Activity of Cysteine, Tryptophan, and Methionine Residues in Continuous Phase β-Lactoglobulin in Oil-in-Water Emulsions. J. Agric. Food Chem. 2005, 53, 10248–10253. [Google Scholar] [CrossRef]
- Atmaca, G. Antioxidant effects of sulfur-containing amino acids. Yonsei Med. J. 2004, 45, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.L.S.; Kim, Y.; Xu, H.; Kim, W.K. The effect of total sulfur amino acid levels on growth performance and bone metabolism in pullets under heat stress. Poult. Sci. 2020, 99, 5783–5791. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Macelline, S.P.; Chrystal, P.V.; Selle, P.H. Progress towards reduced-crude protein diets for broiler chickens and sustainable chicken-meat production. J. Anim. Sci. Biotechnol. 2021, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Barekatain, R.; Chalvon-Demersay, T.; McLaughlan, C.; Lambert, W. Intestinal Barrier Function and Performance of Broiler Chickens Fed Additional Arginine, Combination of Arginine and Glutamine or an Amino Acid-Based Solution. Animals 2021, 11, 2416. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Shafi, M.E.; Abdulsalam, N.M.; Nagadi, S.A.; Wang, J.; Kim, W.K. Amino Acids Supplementation Affects Sustainability of Productive and Meat Quality, Survivability and Nitrogen Pollution of Broiler Chickens during the Early Life. Life 2022, 12, 2100. [Google Scholar] [CrossRef]
- Attia, Y.A.; Bovera, F.; Wang, J.; Al-Harthi, M.A.; Kim, W.K. Multiple Amino Acid Supplementations to Low-Protein Diets: Effect on Performance, Carcass Yield, Meat Quality and Nitrogen Excretion of Finishing Broilers under Hot Climate Conditions. Animals 2020, 10, 973. [Google Scholar] [CrossRef]
- Lambert, W.; Berrocoso, J.D.; Swart, B.; van Tol, M.; Bruininx, E.; Willems, E. Reducing dietary crude protein in broiler diets positively affects litter quality without compromising growth performance whereas a reduction in dietary electrolyte balance further improves litter quality but worsens feed efficiency. Anim. Feed. Sci. Technol. 2023, 297, 115571. [Google Scholar] [CrossRef]
- Cobb-Vantress. Cobb500 Broiler Performance & Nutrition Supplement. 2018. Available online: https://www.cobb-vantress.com/assets/5a88f2e793/Broiler-Performance-Nutrition-Supplement.pdf (accessed on 23 February 2023).
- Cobb-Vantress. Cobb Broiler Management Guide. 2018. Available online: https://www.cobb-vantress.com/assets/Cobb-Files/045bdc8f45/Broiler-Guide-2021-min.pdf (accessed on 23 February 2023).
- Wang, J.; Patterson, R.; Kim, W. Effects of phytase and multicarbohydrase on growth performance, bone mineralization, and nutrient digestibility in broilers fed a nutritionally reduced diet. J. Appl. Poult. Res. 2021, 30, 100146. [Google Scholar] [CrossRef]
- Chen, C.; Kim, W. The application of micro-CT in egg-laying hen bone analysis: Introducing an automated bone separation algorithm. Poult. Sci. 2020, 99, 5175–5183. [Google Scholar] [CrossRef]
- Sharma, M.K.; Liu, G.; White, D.L.; Tompkins, Y.H.; Kim, W.K. Graded levels of Eimeria challenge altered the microstructural architecture and reduced the cortical bone growth of femur of Hy-Line W-36 pullets at early stage of growth (0–6 wk of age). Poult. Sci. 2023, 102, 102888. [Google Scholar] [CrossRef]
- Cooper, D.M.L.; Kawalilak, C.E.; Harrison, K.; Johnston, B.D.; Johnston, J.D. Cortical Bone Porosity: What Is It, Why Is It Important, and How Can We Detect It? Curr. Osteoporos. Rep. 2016, 14, 187–198. [Google Scholar] [CrossRef]
- Xiong, Y.; He, T.; Wang, Y.; Liu, W.V.; Hu, S.; Zhang, Y.; Wen, D.; Hou, B.; Li, Y.; Zhang, P.; et al. CKD Stages, Bone Metabolism Markers, and Cortical Porosity Index: Associations and Mediation Effects Analysis. Front. Endocrinol. 2021, 12, 775066. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, E.; Benavides-Reyes, C.; Torres, C.; Dominguez-Gasca, N.; Garcia-Ruiz, A.I.; Gonzalez-Lopez, S.; Rodriguez-Navarro, A.B. Changes with age (from 0 to 37 D) in tibiae bone mineralization, chemical composition and structural organization in broiler chickens. Poult. Sci. 2019, 98, 5215–5225. [Google Scholar] [CrossRef]
- Norman, T.L.; Little, T.M.; Yeni, Y.N. Age-related changes in porosity and mineralization and in-service damage accumulation. J. Biomech. 2008, 41, 2868–2873. [Google Scholar] [CrossRef]
- Ditscheid, B.; Fünfstück, R.; Busch, M.; Schubert, R.; Gerth, J.; Jahreis, G. Effect of L-methionine supplementation on plasma homocysteine and other free amino acids: A placebo-controlled double-blind cross-over study. Eur. J. Clin. Nutr. 2005, 59, 768–775. [Google Scholar] [CrossRef]
- Xie, M.; Hou, S.S.; Huang, W.; Fan, H.P. Effect of Excess Methionine and Methionine Hydroxy Analogue on Growth Performance and Plasma Homocysteine of Growing Pekin Ducks. Poult. Sci. 2007, 86, 1995–1999. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, Y.; Yang, J.; Wan, X.; Yang, H.; Wang, Z. Hyperhomocysteinemia Induced by Methionine Excess Is Effectively Suppressed by Betaine in Geese. Animals 2020, 10, 1642. [Google Scholar] [CrossRef]
- Herrmann, M.; Wildemann, B.; Claes, L.; Klohs, S.; Ohnmacht, M.; Taban-Shomal, O.; Hübner, U.; Pexa, A.; Umanskaya, N.; Herrmann, W. Experimental Hyperhomocysteinemia Reduces Bone Quality in Rats. Clin. Chem. 2007, 53, 1455–1461. [Google Scholar] [CrossRef]
- Herrmann, M.; Widmann, T.; Herrmann, W. Homocysteine—A newly recognised risk factor for osteoporosis. Clin. Chem. Lab. Med. (CCLM) 2005, 43, 1111–1117. [Google Scholar] [CrossRef]
- Behera, J.; Bala, J.; Nuru, M.; Tyagi, S.C.; Tyagi, N. Homocysteine as a Pathological Biomarker for Bone Disease. J. Cell Physiol. 2017, 232, 2704–2709. [Google Scholar] [CrossRef]
- Fratoni, V.; Brandi, M.L. B vitamins, homocysteine and bone health. Nutrients 2015, 7, 2176–2192. [Google Scholar] [CrossRef]
- Devignes, C.-S.; Carmeliet, G.; Stegen, S. Amino acid metabolism in skeletal cells. Bone Rep. 2022, 17, 101620. [Google Scholar] [CrossRef]
- Lorenzo, J.; Horowitz, M.; Choi, Y. Osteoimmunology: Interactions of the bone and immune system. Endocr. Rev. 2008, 29, 403–440. [Google Scholar] [CrossRef]
- Prisby, R.D. Mechanical, hormonal and metabolic influences on blood vessels, blood flow and bone. J. Endocrinol. 2017, 235, R77–R100. [Google Scholar] [CrossRef]
- Mori, G.; D’Amelio, P.; Faccio, R.; Brunetti, G. The Interplay between the bone and the immune system. Clin. Dev. Immunol. 2013, 2013, 720504. [Google Scholar] [CrossRef]
- Fang, C.C.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Kuang, S.Y.; Tang, L.; Liu, X.A.; Zhou, X.Q. Effects of dietary methionine on growth performance, muscle nutritive deposition, muscle fibre growth and type I collagen synthesis of on-growing grass carp (Ctenopharyngodon idella). Br. J. Nutr. 2021, 126, 321–336. [Google Scholar] [CrossRef]
- Barzel, U.S.; Massey, L.K. Excess dietary protein can adversely affect bone. J. Nutr. 1998, 128, 1051–1053. [Google Scholar] [CrossRef]
- Heaney, R.P.; Layman, D.K. Amount and type of protein influences bone health. Am. J. Clin. Nutr. 2008, 87, 1567S–1570S. [Google Scholar] [CrossRef]
- Cao, J.J. High Dietary Protein Intake and Protein-Related Acid Load on Bone Health. Curr. Osteoporos. Rep. 2017, 15, 571–576. [Google Scholar] [CrossRef]
- Maurer, M.; Riesen, W.; Muser, J.; Hulter, H.N.; Krapf, R. Neutralization of Western diet inhibits bone resorption independently of K intake and reduces cortisol secretion in humans. Am. J. Physiol. -Renal. Physiol. 2003, 284, F32–F40. [Google Scholar] [CrossRef]
- Krieger, N.S.; Frick, K.K.; Bushinsky, D.A. Mechanism of acid-induced bone resorption. Curr. Opin. Nephrol. Hypertens. 2004, 13, 423–436. [Google Scholar] [CrossRef]
- Sukumar, D.; Ambia-Sobhan, H.; Zurfluh, R.; Schlussel, Y.; Stahl, T.J.; Gordon, C.L.; Shapses, S.A. Areal and volumetric bone mineral density and geometry at two levels of protein intake during caloric restriction: A randomized, controlled trial. J. Bone Miner. Res. 2011, 26, 1339–1348. [Google Scholar] [CrossRef]
- Cao, J.J.; Pasiakos, S.M.; Margolis, L.M.; Sauter, E.R.; Whigham, L.D.; McClung, J.P.; Young, A.J.; Combs, G.F., Jr. Calcium homeostasis and bone metabolic responses to high-protein diets during energy deficit in healthy young adults: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 99, 400–407. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NCP 1 | LCP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ingredient | Met 2.8 | Met 4.4 | Met 6.0 | Met 7.6 | Met 9.2 | Met 2.8 | Met 4.4 | Met 6.0 | Met 7.6 | Met 9.2 |
| Corn | 692 | 692 | 692 | 692 | 692 | 730 | 730 | 730 | 730 | 730 |
| Soybean Meal | 248 | 248 | 248 | 248 | 248 | 221 | 221 | 221 | 221 | 221 |
| Soybean Oil | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 1.10 | 1.10 | 1.10 | 1.10 | 1.10 |
| Common Salt | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 |
| Limestone | 11.8 | 11.8 | 11.8 | 11.8 | 11.8 | 11.9 | 11.9 | 11.9 | 11.9 | 11.9 |
| Dicalcium Phosphate | 7.90 | 7.90 | 7.90 | 7.90 | 7.90 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Vitamin Premix 2 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Mineral Premix 3 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 | 0.80 |
| DL-Methionine | 0.00 | 1.60 | 3.20 | 4.80 | 6.40 | 0.10 | 1.70 | 3.30 | 4.90 | 6.50 |
| L-Lysine HCl | 4.10 | 4.10 | 4.10 | 4.10 | 4.10 | 2.70 | 2.70 | 2.70 | 2.70 | 2.70 |
| L-Glutamate | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | |||||
| Threonine | 1.30 | 1.30 | 1.30 | 1.30 | 1.30 | 0.60 | 0.60 | 0.60 | 0.60 | 0.60 |
| Arginine | 1.10 | 1.10 | 1.10 | 1.10 | 1.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| L-Cystine | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | |||||
| Isoleucine | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | |||||
| Glycine | 10.0 | 9.20 | 8.40 | 7.60 | 6.80 | 5.00 | 4.30 | 3.50 | 2.60 | 1.80 |
| Sand | 7.60 | 6.80 | 6.00 | 5.20 | 4.60 | 14.5 | 13.6 | 12.80 | 12.10 | 11.30 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Calculated Nutrients (g/kg) and Energy | ||||||||||
| Protein | 200 | 200 | 200 | 200 | 200 | 170 | 170 | 170 | 170 | 170 |
| ME, kcal/kg | 3030 | 3030 | 3030 | 3030 | 3030 | 3030 | 3030 | 3030 | 3030 | 3030 |
| Ca | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Available P | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 | 2.90 |
| Lysine | 11.2 | 11.2 | 11.2 | 11.2 | 11.2 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| Methionine | 2.80 | 4.40 | 6.00 | 7.60 | 9.20 | 2.80 | 4.40 | 6.00 | 7.60 | 9.20 |
| TSAA | 5.30 | 6.90 | 8.50 | 10.1 | 11.7 | 5.30 | 6.90 | 8.50 | 10.1 | 11.7 |
| Threonine | 7.30 | 7.30 | 7.30 | 7.30 | 7.30 | 6.20 | 6.20 | 6.20 | 6.20 | 6.20 |
| Arginine | 11.8 | 11.8 | 11.8 | 11.8 | 11.8 | 10.0 | 10.0 | 10.0 | 10.0 | 10.0 |
| Analyzed Amino Acids, g/kg | ||||||||||
| Protein | 195 | 199 | 211 | 211 | 204 | 177 | 182 | 176 | 181 | 176 |
| Lysine | 13.1 | 12.5 | 12.7 | 13.0 | 13.2 | 10.5 | 11.4 | 11.4 | 11.6 | 11.3 |
| Methionine | 2.90 | 4.50 | 5.90 | 7.80 | 8.90 | 2.90 | 4.50 | 6.30 | 7.20 | 8.80 |
| TSAA | 5.90 | 7.40 | 9.10 | 10.9 | 11.5 | 5.80 | 7.60 | 9.20 | 10.3 | 11.8 |
| Threonine | 7.60 | 7.60 | 7.70 | 7.60 | 7.70 | 6.50 | 7.10 | 7.00 | 7.10 | 7.00 |
| Arginine | 12.1 | 11.7 | 11.6 | 12.4 | 12.5 | 10.1 | 11.3 | 11.0 | 11.5 | 10.6 |
| Glutamate | 39.2 | 37.3 | 37.3 | 39.3 | 39.2 | 30.2 | 32.7 | 32.2 | 33.3 | 31.5 |
| Amino Acids to Lysine Ratios | ||||||||||
| Lysine | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Methionine | 22.2 | 34.1 | 46.5 | 59.8 | 67.4 | 27.1 | 39.5 | 54.8 | 62.1 | 77.9 |
| TSAA | 45.2 | 56.3 | 71.7 | 83.8 | 90.5 | 54.8 | 66.7 | 80.7 | 88.8 | 104 |
| Threonine | 57.9 | 60.4 | 60.6 | 58.3 | 58.0 | 61.4 | 62.3 | 60.9 | 61.2 | 61.5 |
| Arginine | 92.7 | 89.3 | 91.3 | 95.7 | 94.7 | 96.2 | 99.1 | 96.5 | 98.7 | 93.8 |
| Glutamate | 300 | 286 | 294 | 303 | 297 | 288 | 287 | 282 | 287 | 278 |
| Parameters (Abbreviation) | Description | Standard Unit |
|---|---|---|
| Bone mineral density (BMD) | Measure the bone mineral content per unit of volume | g/cm3 |
| Bone mineral content (BMC) | Measure the bone mineral content of the tissue. The value is calculated by BMD × TV | g |
| Tissue volume (TV) | The volume of the entire region of interest, including pores and cavity inside the bone | mm3 |
| Bone volume (BV) | Volume of the bone segments | mm3 |
| Bone volume fraction (BV/TV) | Ratio of the bone volume to the tissue volume | % |
| Number of closed pores (NP) | Number of closed pores within the bone segments | |
| Volume of closed pores (VP) | Total volume of the closed pores | mm3 |
| Closed pore percentage (PP) | The volume of closed pores as a percentage of the bone volume | % |
| Trabecular number (TBN) | The average number of trabeculae per unit length | 1/mm |
| Trabecular thickness (TBT) | Mean thickness of trabeculae, measured using 3D methods | mm |
| Trabecular separation (TBS) | Mean distance between trabeculae, measured using 3D methods | mm |
| Connectivity density (CD) | A measure of the degree of connectivity of trabeculae normalized by tissue volume | 1/mm3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Choppa, V.S.R.; Sharma, M.K.; Ko, H.; Choi, J.; Kim, W.K. Effects of Methionine Supplementation Levels in Normal or Reduced Protein Diets on the Body Composition and Femur Bone Characteristics of Broilers Challenged with Coccidia. Animals 2024, 14, 917. https://doi.org/10.3390/ani14060917
Liu G, Choppa VSR, Sharma MK, Ko H, Choi J, Kim WK. Effects of Methionine Supplementation Levels in Normal or Reduced Protein Diets on the Body Composition and Femur Bone Characteristics of Broilers Challenged with Coccidia. Animals. 2024; 14(6):917. https://doi.org/10.3390/ani14060917
Chicago/Turabian StyleLiu, Guanchen, Venkata Sesha Reddy Choppa, Milan Kumar Sharma, Hanseo Ko, Janghan Choi, and Woo Kyun Kim. 2024. "Effects of Methionine Supplementation Levels in Normal or Reduced Protein Diets on the Body Composition and Femur Bone Characteristics of Broilers Challenged with Coccidia" Animals 14, no. 6: 917. https://doi.org/10.3390/ani14060917