
A Review of Potential Feed Additives Intended for Carbon Footprint Reduction through Methane Abatement in Dairy Cattle

Abstract
:Simple Summary
Abstract
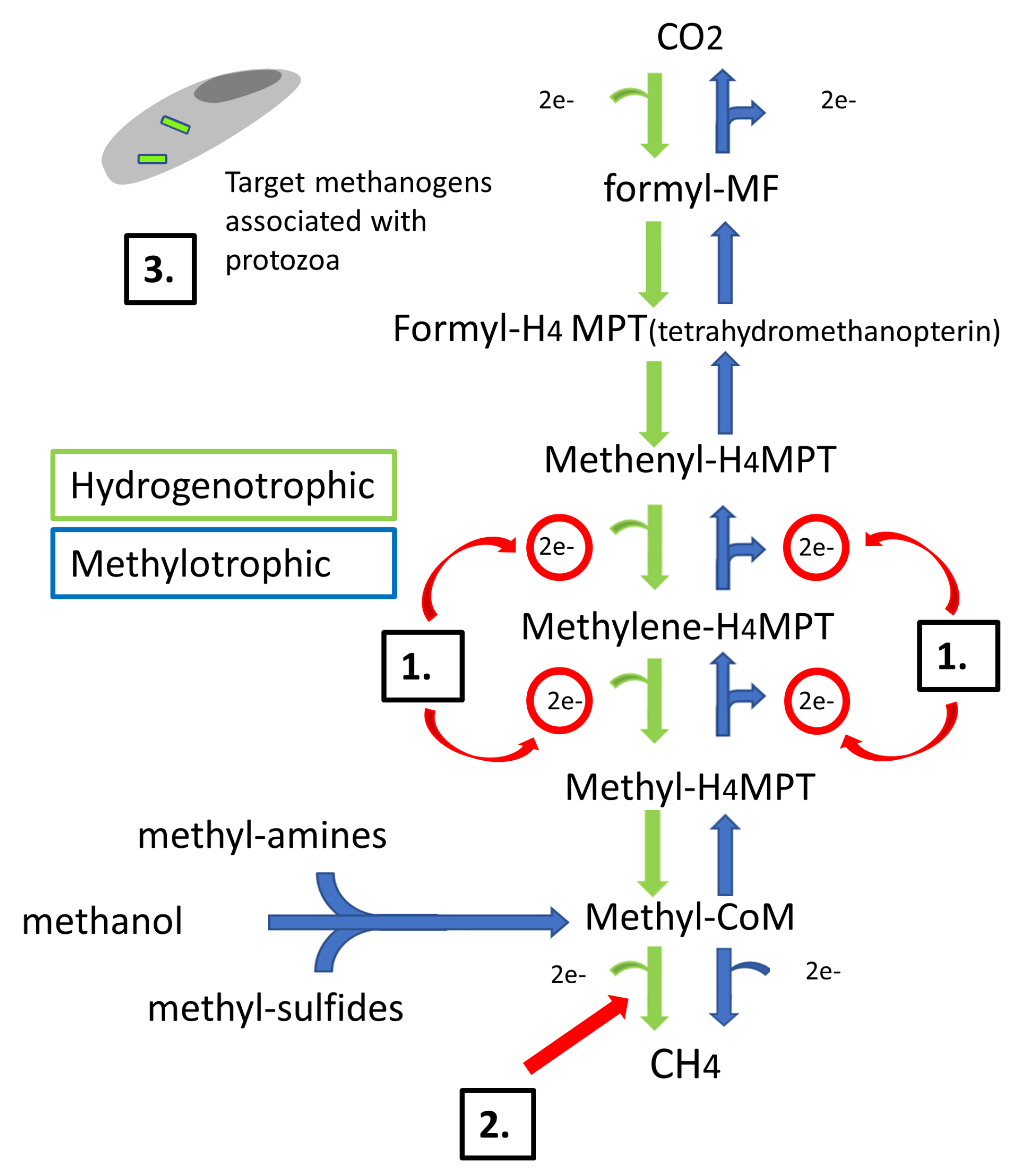
1. Introduction
1.1. Garlic Oil
1.2. Nitrate
1.3. Ascophyllum Nodosum
1.4. Asparagopsis
1.5. Lactic Acid Bacteria
1.6. Chitosan
1.7. Essential Oil Blends
1.8. 3-Nitrooxypropanol
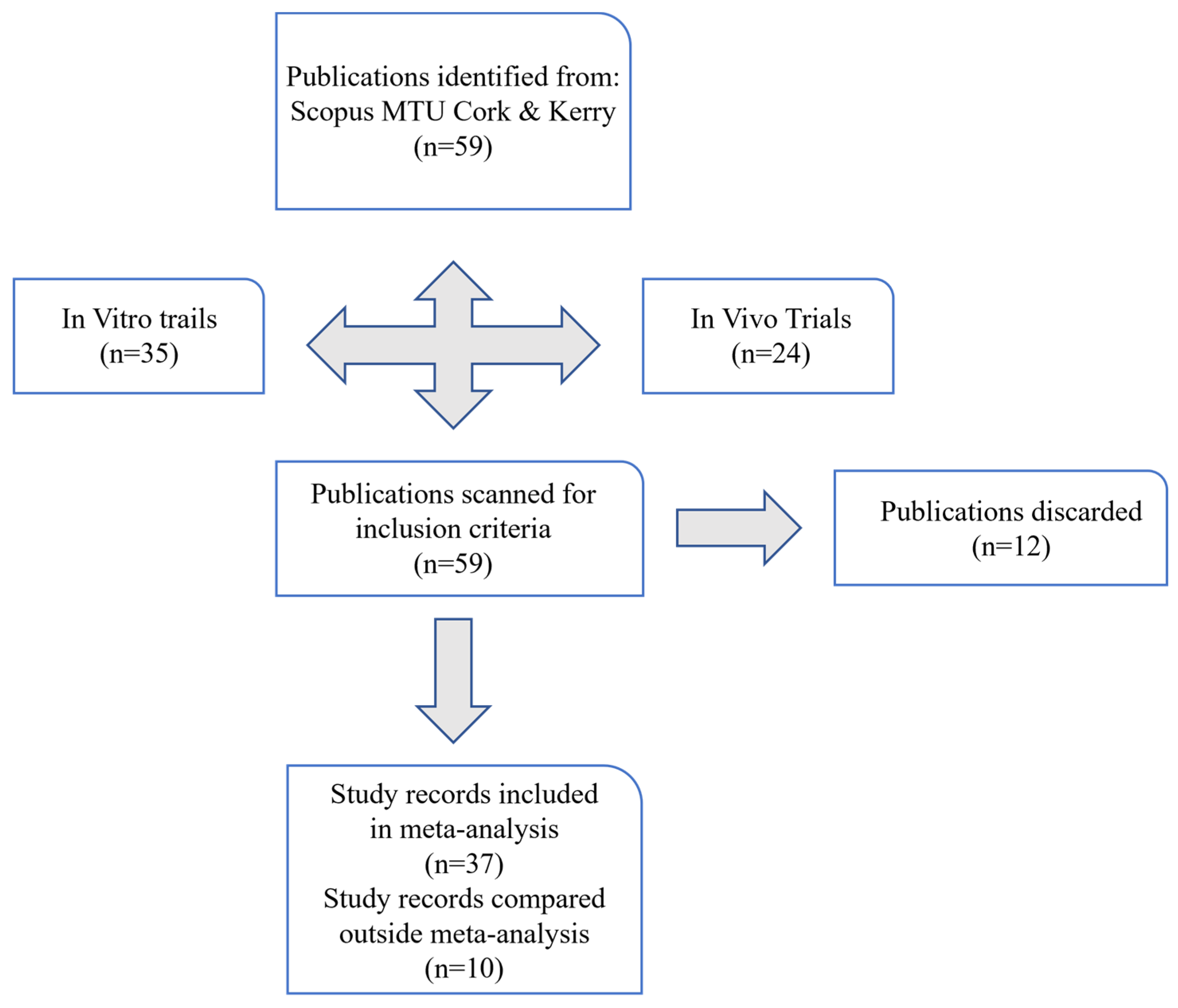
2. Materials and Methods
3. Results
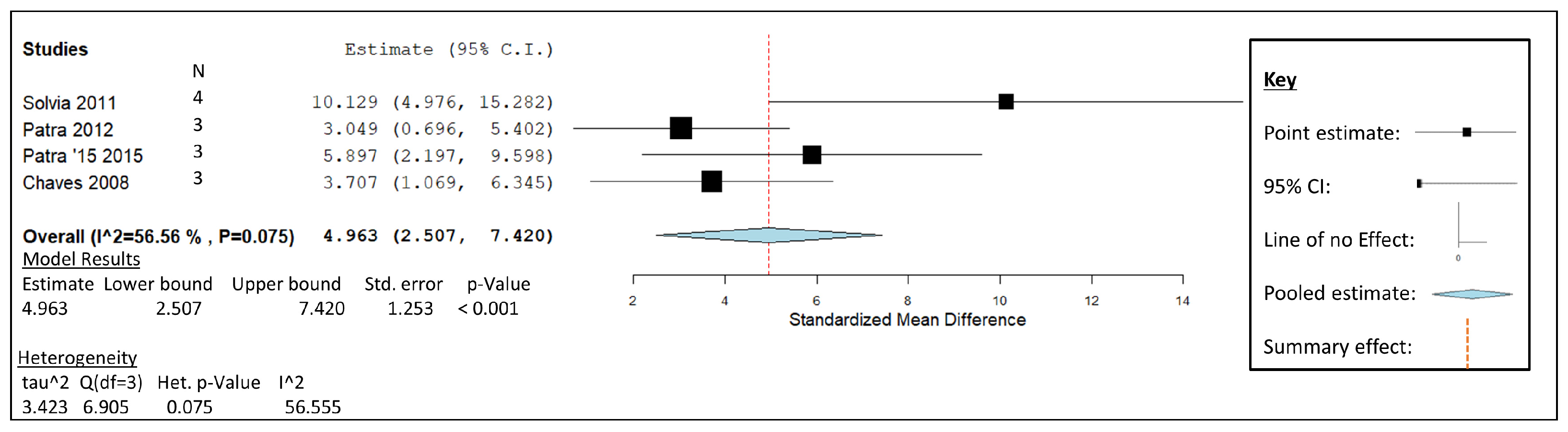
3.1. Garlic Oil
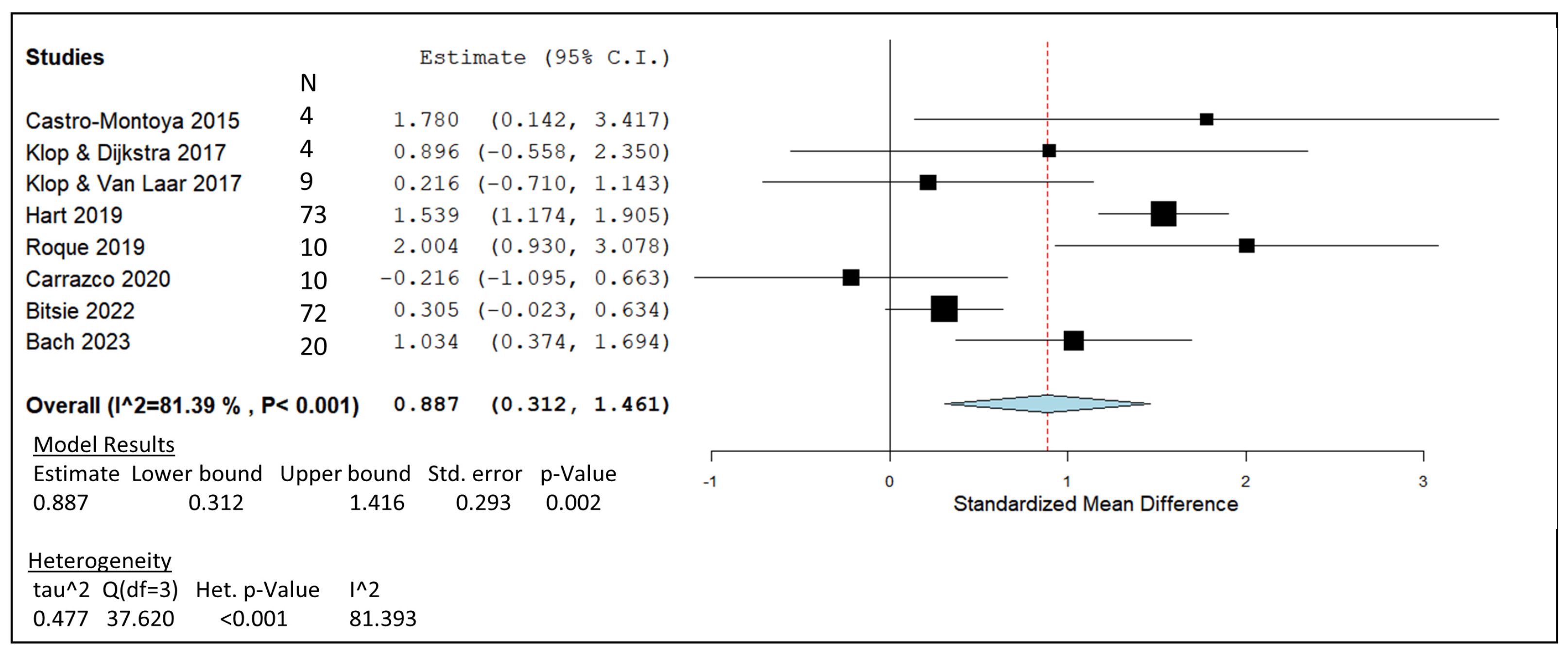
3.2. Essential Oil Blends
3.3. Nitrate
3.4. Chitosan
3.5. 3-Nitrooxypropanol
3.6. Ascophyllum Nodosum
3.7. Asparagopsis
3.8. Lactobacillus Plantarum
3.9. Comparison Meta-Analysis Plot
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lashof, D.A.; Ahuja, D.R. Relative Contributions of Greenhouse Gas Emissions to Global Warming. Nature 1990, 344, 529–531. [Google Scholar] [CrossRef]
- Andersen, S.O.; Davidson, O.; Kuijpers, L.; Metz, B. Safeguarding the Ozone Layer and the Global Climate System; IPCC: Geneva, Switzerland, 2005. [Google Scholar]
- United Nations United Nations Online Platform for Voluntary Cancellation of Certified Emission Reductions (CERs). Available online: https://offset.climateneutralnow.org/ (accessed on 2 December 2023).
- European Commission Communication Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions on an EU Strategy to Reduce Methane Emissions. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52020DC0663 (accessed on 29 January 2024).
- Food and Agriculture Organization of the United Nations Livestock Solutions for Climate Change; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017.
- Patra, A.K.; Yu, Z. Effects of Essential Oils on Methane Production and Fermentation by, and Abundance and Diversity of, Rumen Microbial Populations. Appl. Environ. Microbiol. 2012, 78, 4271–4280. [Google Scholar] [CrossRef] [PubMed]
- Duin, E.C.; Wagner, T.; Shima, S.; Prakash, D.; Cronin, B.; Yáñez-Ruiz, D.R.; Duval, S.; Rümbeli, R.; Stemmler, R.T.; Thauer, R.K.; et al. Mode of Action Uncovered for the Specific Reduction of Methane Emissions from Ruminants by the Small Molecule 3-Nitrooxypropanol. Proc. Natl. Acad. Sci. USA 2016, 113, 6172–6177. [Google Scholar] [CrossRef] [PubMed]
- Van Wesemael, D.; Vandaele, L.; Ampe, B.; Cattrysse, H.; Duval, S.; Kindermann, M.; Fievez, V.; De Campeneere, S.; Peiren, N. Reducing Enteric Methane Emissions from Dairy Cattle: Two Ways to Supplement 3-Nitrooxypropanol. J. Dairy Sci. 2019, 102, 1780–1787. [Google Scholar] [CrossRef] [PubMed]
- Soliva, C.R.; Amelchanka, S.L.; Duval, S.M.; Kreuzer, M. Ruminal Methane Inhibition Potential of Various Pure Compounds in Comparison with Garlic Oil as Determined with a Rumen Simulation Technique (Rusitec). Br. J. Nutr. 2011, 106, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Villar, M.L.; Hegarty, R.S.; Nolan, J.V.; Godwin, I.R.; McPhee, M. The Effect of Dietary Nitrate and Canola Oil Alone or in Combination on Fermentation, Digesta Kinetics and Methane Emissions from Cattle. Anim. Feed Sci. Technol. 2020, 259, 114294. [Google Scholar] [CrossRef]
- Kinley, R.D.; Martinez-Fernandez, G.; Matthews, M.K.; de Nys, R.; Magnusson, M.; Tomkins, N.W. Mitigating the Carbon Footprint and Improving Productivity of Ruminant Livestock Agriculture Using a Red Seaweed. J. Clean. Prod. 2020, 259, 120836. [Google Scholar] [CrossRef]
- Hook, S.E.; Wright, A.D.G.; McBride, B.W. Methanogens: Methane Producers of the Rumen and Mitigation Strategies. Archaea 2010, 2010, 11. [Google Scholar] [CrossRef]
- Ryle, M.; Ørskov, E.R.; Ryle, M.; Ørskov, E.R. Energy Nutrition of Rumen Micro-Organisms. In Energy Nutrition in Ruminants; Springer: Dordrecht, The Netherlands, 1990; pp. 10–27. [Google Scholar]
- McAllister, T.A.; Okine, E.K.; Mathison, G.W.; Cheng, K.J. Dietary, Environmental and Microbiological Aspects of Methane Production in Ruminants. Can. J. Anim. Sci. 1996, 76, 231–243. [Google Scholar] [CrossRef]
- Lee, S.-R.; Cho, Y.; Ju, H.K.; Kim, E. Theoretical Methane Emission Estimation from Volatile Fatty Acids in Bovine Rumen Fluid. Appl. Sci. 2021, 11, 7730. [Google Scholar] [CrossRef]
- Janssen, P.H. Influence of Hydrogen on Rumen Methane Formation and Fermentation Balances through Microbial Growth Kinetics and Fermentation Thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Leahy, S.C.; Janssen, P.H.; Attwood, G.T.; Mackie, R.I.; McAllister, T.A.; Kelly, W.J. Electron Flow: Key to Mitigating Ruminant Methanogenesis. Trends Microbiol. 2022, 30, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Krumholz, L.R.; Forsberg, C.W.; Veira, D.M. Association of Methanogenic Bacteria with Rumen Protozoa. Can. J. Microbiol. 1983, 29, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Klieve, A.V.; Hegarty, R.S. Opportunities for Biological Control of Ruminal Methanogenesis. Aust. J. Agric. Res. 1999, 50, 1315–1319. [Google Scholar] [CrossRef]
- Brul, S.; Stumm, C.K. Symbionts and Organelles in Ancrobic Protozoa and Fungi. Trends Ecol. Evol. 1994, 9, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Karekar, S.; Stefanini, R.; Ahring, B. Homo-Acetogens: Their Metabolism and Competitive Relationship with Hydrogenotrophic Methanogens. Microorganisms 2022, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, M.; Schwab, C.; Borg Jensen, B.; Engberg, R.M.; Spang, A.; Canibe, N.; Højberg, O.; Milinovich, G.; Fragner, L.; Schleper, C.; et al. Methylotrophic Methanogenic Thermoplasmata Implicated in Reduced Methane Emissions from Bovine Rumen. Nat. Commun. 2013, 4, 1428. [Google Scholar] [CrossRef]
- Wolin, M.J. The Rumen Fermentation: A Model for Microbial Interactions in Anaerobic Ecosystems; Springer: Boston, MA, USA, 1979; pp. 49–77. [Google Scholar]
- Bergman, E.N. Energy Contributions of Volatile Fatty Acids from the Gastrointestinal Tract in Various Species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Okine, E.K.; Mathison, G.W.; Hardin, R.T. Relations between Passage Rates of Rumen Fluid and Particulate Matter and Foam Production in Rumen Contents of Cattle Fed on Different Diets Ad Lib. Br. J. Nutr. 1989, 61, 387–395. [Google Scholar] [CrossRef]
- Eugène, M.; Klumpp, K.; Sauvant, D. Methane Mitigating Options with Forages Fed to Ruminants. Grass Forage Sci. 2021, 76, 196–204. [Google Scholar] [CrossRef]
- Dong, L.; Li, B.; Diao, Q. Effects of Dietary Forage Proportion on Feed Intake, Growth Performance, Nutrient Digestibility, and Enteric Methane Emissions of Holstein Heifers at Various Growth Stages. Animals 2019, 9, 725. [Google Scholar] [CrossRef]
- Boadi, D.A.; Wittenberg, K.M.; Scott, S.L.; Burton, D.; Buckley, K.; Small, J.A.; Ominski, K.H. Effect of Low and High Forage Diet on Enteric and Manure Pack Greenhouse Gas Emissions from a Feedlot. Can. J. Anim. Sci. 2004, 84, 445–453. [Google Scholar] [CrossRef]
- Hassanat, F.; Gervais, R.; Julien, C.; Massé, D.I.; Lettat, A.; Chouinard, P.Y.; Petit, H.V.; Benchaar, C. Replacing Alfalfa Silage with Corn Silage in Dairy Cow Diets: Effects on Enteric Methane Production, Ruminal Fermentation, Digestion, N Balance, and Milk Production. J. Dairy Sci. 2013, 96, 4553–4567. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane Emissions from Cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Danielsson, R.; Ramin, M.; Bertilsson, J.; Lund, P.; Huhtanen, P. Evaluation of a Gas In Vitro System for Predicting Methane Production In Vivo. J. Dairy Sci. 2017, 100, 8881–8894. [Google Scholar] [CrossRef] [PubMed]
- Hatew, B.; Cone, J.W.; Pellikaan, W.F.; Podesta, S.C.; Bannink, A.; Hendriks, W.H.; Dijkstra, J. Relationship between In Vitro and In Vivo Methane Production Measured Simultaneously with Different Dietary Starch Sources and Starch Levels in Dairy Cattle. Anim. Feed Sci. Technol. 2015, 202, 20–31. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Newbold, C.J.; van Nevel, C.J.; Demeyer, D.I. Manipulation of Ruminal Fermentation. In The Rumen Microbial Ecosystem; Springer: Dordrecht, The Netherlands, 1997; pp. 523–632. [Google Scholar]
- Russel, J.B.; Strobel, H.J. Effect of Lonophores on Ruminal Fermentation. Appl. Environ. Microbiol. 1989, 55, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.L.; Dijkstra, J.; Bannink, A.; Kebreab, E.; Hook, S.E.; Archibeque, S.; France, J. Quantifying the Effect of Monensin Dose on the Rumen Volatile Fatty Acid Profile in High-Grain-Fed Beef Cattle. J. Anim. Sci. 2012, 90, 2717–2726. [Google Scholar] [CrossRef] [PubMed]
- European Commission Food and Feed Information Portal Database _ FIP. Available online: https://ec.europa.eu/food/food-feed-portal/screen/feed-additives/search (accessed on 16 December 2023).
- Ankri, S.; Mirelman, D. Antimicrobial Properties of Allicin from Garlic. Microbes Infect. 1999, 1, 125–129. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.; Gambacorta, A.; Gliozzi, A. Structure, Biosynthesis, and Physicochemical Properties of Archaebacterial Lipids. Microbiol. Rev. 1986, 50, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.H.S.; Xu, S. Effects of Allyl Mercaptan and Various Allium-Derived Compounds on Cholesterol Synthesis and Secretion in Hep-G2 Cells. Comp. Biochem. Physiol.—C Pharmacol. Toxicol. Endocrinol. 2000, 126, 195–201. [Google Scholar] [CrossRef]
- Gebhardt, R.; Beck, H. Differential Inhibitory Effects of Garlic-Derived Organosulfur Compounds on Cholesterol Biosynthesis in Primary Rat Hepatocyte Cultures. Lipids 1996, 31, 1269–1276. [Google Scholar] [CrossRef]
- Ohene-Adjei, S.; Chaves, A.V.; McAllister, T.A.; Benchaar, C.; Teather, R.M.; Forster, R.J. Evidence of Increased Diversity of Methanogenic Archaea with Plant Extract Supplementation. Microb. Ecol. 2008, 56, 234–242. [Google Scholar] [CrossRef]
- Sari, N.F.; Ray, P.; Rymer, C.; Kliem, K.E.; Stergiadis, S. Garlic and Its Bioactive Compounds: Implications for Methane Emissions and Ruminant Nutrition. Animals 2022, 12, 2998. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.V.; He, M.L.; Yang, W.Z.; Hristov, A.N.; McAllister, T.A.; Benchaar, C. Effects of Essential Oils on Proteolytic, Deaminative and Methanogenic Activities of Mixed Ruminal Bacteria. Can. J. Anim. Sci. 2008, 88, 117–122. [Google Scholar] [CrossRef]
- Patra, A.K.; Yu, Z. Effects of Adaptation of In Vitro Rumen Culture to Garlic Oil, Nitrate, and Saponin and Their Combinations on Methanogenesis, Fermentation, and Abundances and Diversity of Microbial Populations. Front. Microbiol. 2015, 6, 1434. [Google Scholar] [CrossRef]
- Lawson, L.D.; Gardner, C.D. Composition, Stability, and Bioavailability of Garlic Products Used in a Clinical Trial. J. Agric. Food Chem. 2005, 53, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Carro, M.D.; Kamel, C. Effect of Garlic Oil and Four of Its Compounds on Rumen Microbial Fermentation. J. Dairy Sci. 2005, 88, 4393–4404. [Google Scholar] [CrossRef] [PubMed]
- Dan Gould and Frank Garry Sulfur Intake in Cattle. Available online: http://csu-cvmbs.colostate.edu/vth/livestock/integrated-livestock-management/Pages/sulfur-intake-in-cattle.aspx (accessed on 8 March 2021).
- Van Zijderveld, S.M.; Gerrits, W.J.J.; Dijkstra, J.; Newbold, J.R.; Hulshof, R.B.A.; Perdok, H.B. Persistency of Methane Mitigation by Dietary Nitrate Supplementation in Dairy Cows. J. Dairy Sci. 2011, 94, 4028–4038. [Google Scholar] [CrossRef]
- Jones, G.A. Dissimilatory Metabolism of Nitrate by the Rumen Microbiota. Can. J. Microbiol. 1972, 18, 1783–1787. [Google Scholar] [CrossRef]
- Lund, P.; Dahl, R.; Yang, H.J.; Hellwing, A.L.F.; Cao, B.B.; Weisbjerg, M.R. The Acute Effect of Addition of Nitrate on In Vitro and In Vivo Methane Emission in Dairy Cows. Anim. Prod. Sci. 2014, 54, 1432–1435. [Google Scholar] [CrossRef]
- van Zijderveld, S.M.; Gerrits, W.J.J.; Apajalahti, J.A.; Newbold, J.R.; Dijkstra, J.; Leng, R.A.; Perdok, H.B. Nitrate and Sulfate: Effective Alternative Hydrogen Sinks for Mitigation of Ruminal Methane Production in Sheep. J. Dairy Sci. 2010, 93, 5856–5866. [Google Scholar] [CrossRef] [PubMed]
- Alaboudi, A.R.; Jones, G.A. Effect of Acclimation to High Nitrate Intakes on Some Rumen Fermentation Parameters in Sheep. Can. J. Anim. Sci. 1985, 65, 841–849. [Google Scholar] [CrossRef]
- Lin, M.; Schaefer, D.M.; Zhao, G.Q.; Meng, Q.X. Effects of Nitrate Adaptation by Rumen Inocula Donors and Substrate Fiber Proportion on in Vitro Nitrate Disappearance, Methanogenesis, and Rumen Fermentation Acid. Animal 2013, 7, 1099–1105. [Google Scholar] [CrossRef]
- Nolan, J.V.; Godwin, I.R.; De Raphélis-Soissan, V.; Hegarty, R.S. Managing the Rumen to Limit the Incidence and Severity of Nitrite Poisoning in Nitrate-Supplemented Ruminants. Anim. Prod. Sci. 2016, 56, 1317–1329. [Google Scholar] [CrossRef]
- Lee, C.; Beauchemin, K.A. Une Revue de l’ajout de Nitrate Dans l’alimentation Des Ruminants: Toxicité Aux Nitrates, Émissions de Méthane et Performance de Production. Can. J. Anim. Sci. 2014, 94, 557–570. [Google Scholar] [CrossRef]
- Zhao, L.; Meng, Q.; Ren, L.; Liu, W.; Zhang, X.; Huo, Y.; Zhou, Z. Effects of Nitrate Addition on Rumen Fermentation, Bacterial Biodiversity and Abundance. Asian-Australasian J. Anim. Sci. 2015, 28, 1433–1441. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.K.; Li, Y.; Li, Y.X. Phlorotannins as Bioactive Agents from Brown Algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar]
- Pal Singh, I.; Bharate, S.B. Phloroglucinol Compounds of Natural Origin. Nat. Prod. Rep. 2006, 23, 558–591. [Google Scholar] [CrossRef]
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Gall, E.A. Interspecific and Temporal Variation in Phlorotannin Levels in an Assemblage of Brown Algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Bach, S.J.; McAllister, T.A. Effects of Phlorotannins from Ascophyllum Nodosum (Brown Seaweed) on in Vitro Ruminal Digestion of Mixed Forage or Barley Grain. Anim. Feed Sci. Technol. 2008, 145, 375–395. [Google Scholar] [CrossRef]
- Belanche, A.; Ramos-Morales, E.; Newbold, C.J. In Vitro Screening of Natural Feed Additives from Crustaceans, Diatoms, Seaweeds and Plant Extracts to Manipulate Rumen Fermentation. J. Sci. Food Agric. 2016, 96, 3069–3078. [Google Scholar] [CrossRef]
- Eason, C.T.; Fennessy, P. Methane Reduction, Health and Regulatory Considerations Regarding Asparagopsis and Bromoform for Ruminants. N. Zeal. J. Agric. Res. 2023, 66, 1–30. [Google Scholar] [CrossRef]
- Paul, N.A.; De Nys, R.; Steinberg, P.D. Chemical Defence against Bacteria in the Red Alga Asparagopsis Armata: Linking Structure with Function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef]
- Johnson, E.D.; Wood, A.S.; Stone, J.B.; Moran, E.T., Jr. Some Effects of Methane Inhibition in Ruminants (Steers). Can. J. Anim. Sci. 1972, 52, 703–712. [Google Scholar] [CrossRef]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis Armata in Lactating Dairy Cows’ Diet Reduces Enteric Methane Emission by over 50 Percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Brooke, C.G.; Roque, B.M.; Shaw, C.; Najafi, N.; Gonzalez, M.; Pfefferlen, A.; De Anda, V.; Ginsburg, D.W.; Harden, M.C.; Nuzhdin, S.V.; et al. Methane Reduction Potential of Two Pacific Coast Macroalgae During in Vitro Ruminant Fermentation. Front. Mar. Sci. 2020, 7, 561. [Google Scholar] [CrossRef]
- Roque, B.M.; Venegas, M.; Kinley, R.D.; De Nys, R.; Duarte, T.L.; Yang, X.; Kebreab, E. Red Seaweed (Asparagopsis Taxiformis) Supplementation Reduces Enteric Methane by over 80 Percent in Beef Steers. PLoS ONE 2021, 16, e0247820. [Google Scholar] [CrossRef]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and Diversity Analysis of Ruminal Methanogenic Populations in Response to the Antimethanogenic Compound Bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Kim, D.H.; Jiang, Y.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Control of Escherichia Coli O157:H7 in Contaminated Alfalfa Silage: Effects of Silage Additives. J. Dairy Sci. 2016, 99, 4427–4436. [Google Scholar] [CrossRef]
- Cao, Y.; Cai, Y.; Takahashi, T.; Yoshida, N.; Tohno, M.; Uegaki, R.; Nonaka, K.; Terada, F. Effect of Lactic Acid Bacteria Inoculant and Beet Pulp Addition on Fermentation Characteristics and in Vitro Ruminal Digestion of Vegetable Residue Silage. J. Dairy Sci. 2011, 94, 3902–3912. [Google Scholar] [CrossRef]
- Huyen, N.T.; Martinez, I.; Pellikaan, W. Using Lactic Acid Bacteria as Silage Inoculants or Direct-Fed Microbials to Improve in Vitro Degradability and Reduce Methane Emissions in Dairy Cows. Agronomy 2020, 10, 1482. [Google Scholar] [CrossRef]
- Jalč, D.; Lauková, A.; Váradyová, Z.; Homolka, P.; Koukolová, V. Effect of Inoculated Grass Silages on Rumen Fermentation and Lipid Metabolism in an Artificial Rumen (RUSITEC). Anim. Feed Sci. Technol. 2009, 151, 55–64. [Google Scholar] [CrossRef]
- Doyle, N.; Mbandlwa, P.; Kelly, W.J.; Attwood, G.; Li, Y.; Ross, R.P.; Stanton, C.; Leahy, S. Use of Lactic Acid Bacteria to Reduce Methane Production in Ruminants, a Critical Review. Front. Microbiol. 2019, 10, 2207. [Google Scholar] [CrossRef]
- Navarro-Villa, A.; O’Brien, M.; López, S.; Boland, T.M.; O’Kiely, P. In Vitro Rumen Methane Output of Red Clover and Perennial Ryegrass Assayed Using the Gas Production Technique (GPT). Anim. Feed Sci. Technol. 2011, 168, 152–164. [Google Scholar] [CrossRef]
- Cao, Y.; Takahashi, T.; Horiguchi, K.; Yoshida, N.; Cai, Y. Methane Emissions from Sheep Fed Fermented or Non-Fermented Total Mixed Ration Containing Whole-Crop Rice and Rice Bran. Anim. Feed Sci. Technol. 2010, 157, 72–78. [Google Scholar] [CrossRef]
- Guo, G.; Shen, C.; Liu, Q.; Zhang, S.; Shao, T.; Wang, C.; Wang, Y.; Xu, Q.; Huo, W. The Effect of Lactic Acid Bacteria Inoculums on in Vitro Rumen Fermentation, Methane Production, Ruminal Cellulolytic Bacteria Populations and Cellulase Activities of Corn Stover Silage. J. Integr. Agric. 2020, 19, 838–847. [Google Scholar] [CrossRef]
- Aydin, R.; Yanar, M.; Kocyigit, R.; Diler, A.; Ozkilicci, T.Z. Effect of Direct-Fed Microbials plus Enzyme Supplementation on the Fattening Performance of Holstein Young Bulls at Two Different Initial Body Weights. African J. Agric. Res. 2009, 4, 548–552. [Google Scholar]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial Properties of Chitosan and Mode of Action: A State of the Art Review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.M.; Garcia-Rodriguez, A. Dose-Response Effects of Chitosans on in Vitro Rumen Digestion and Fermentation of Mixtures Differing in Forage-to-Concentrate Ratios. Anim. Feed Sci. Technol. 2009, 151, 215–227. [Google Scholar] [CrossRef]
- Goiri, I.; Garcia-Rodriguez, A.; Oregui, L.M. Effect of Chitosans on in Vitro Rumen Digestion and Fermentation of Maize Silage. Anim. Feed Sci. Technol. 2009, 148, 276–287. [Google Scholar] [CrossRef]
- Goiri, I.; Garcia-Rodriguez, A.; Oregui, L.M. Effect of Chitosan on Mixed Ruminal Microorganism Fermentation Using the Rumen Simulation Technique (Rusitec). Anim. Feed Sci. Technol. 2009, 152, 92–102. [Google Scholar] [CrossRef]
- Russell, J.B. The Importance of PH in the Regulation of Ruminal Acetate to Propionate Ratio and Methane Production in Vitro. J. Dairy Sci. 1998, 81, 3222–3230. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Cardozo, P.W.; Kamel, C. Effects of Cinnamaldehyde and Garlic Oil on Rumen Microbial Fermentation in a Dual Flow Continuous Culture. J. Dairy Sci. 2005, 88, 2508–2516. [Google Scholar] [CrossRef]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The Effect of Bioflavex® and Its Pure Flavonoid Components on in Vitro Fermentation Parameters and Methane Production in Rumen Fluid from Steers given High Concentrate Diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef]
- Belanche, A.; Newbold, C.J.; Morgavi, D.P.; Bach, A.; Zweifel, B.; Yáñez-Ruiz, D.R. A Meta-Analysis Describing the Effects of the Essential Oils Blend Agolin Ruminant on Performance, Rumen Fermentation and Methane Emissions in Dairy Cows. Animals 2020, 10, 620. [Google Scholar] [CrossRef]
- Castro-Montoya, J.; Peiren, N.; Cone, J.W.; Zweifel, B.; Fievez, V.; De Campeneere, S. In Vivo and In Vitro Effects of a Blend of Essential Oils on Rumen Methane Mitigation. Livest. Sci. 2015, 180, 134–142. [Google Scholar] [CrossRef]
- Klop, G.; Dijkstra, J.; Dieho, K.; Hendriks, W.H.; Bannink, A. Enteric Methane Production in Lactating Dairy Cows with Continuous Feeding of Essential Oils or Rotational Feeding of Essential Oils and Lauric Acid. J. Dairy Sci. 2017, 100, 3563–3575. [Google Scholar] [CrossRef]
- Klop, G.; Van Laar-Van Schuppen, S.; Pellikaan, W.F.; Hendriks, W.H.; Bannink, A.; Dijkstra, J. Changes in In Vitro Gas and Methane Production from Rumen Fluid from Dairy Cows during Adaptation to Feed Additives In Vivo. Animal 2017, 11, 591–599. [Google Scholar] [CrossRef]
- Hart, K.J.; Jones, H.G.; Waddams, K.E.; Worgan, H.J.; Zweifel, B.; Newbold, C.J. An Essential Oil Blend Decreases Methane Emissions and Increases Milk Yield in Dairy Cows. Open J. Anim. Sci. 2019, 09, 259–267. [Google Scholar] [CrossRef]
- Carrazco, A.V.; Peterson, C.B.; Zhao, Y.; Pan, Y.; McGlone, J.J.; Depeters, E.J.; Mitloehner, F.M. The Impact of Essential Oil Feed Supplementation on Enteric Gas Emissions and Production Parameters from Dairy Cattle. Sustainable 2020, 12, 10347. [Google Scholar] [CrossRef]
- Bach, A.; Elcoso, G.; Escartín, M.; Spengler, K.; Jouve, A. Modulation of Milking Performance, Methane Emissions, and Rumen Microbiome on Dairy Cows by Dietary Supplementation of a Blend of Essential Oils. Animal 2023, 17, 100825. [Google Scholar] [CrossRef]
- Hao, Y.Y.; Brackett, R.E.; Doyle, M.P. Inhibition of Listeria Monocytogenes and Aeromonas Hydrophila by Plant Extracts in Refrigerated Cooked Beef. J. Food Prot. 1998, 61, 307–312. [Google Scholar] [CrossRef]
- Stecchini, M.L.; Sarais, I.; Giavedoni, P. Effect of Essential Oils on Aeromonas Hydrophila in a Culture Medium and in Cooked Pork. J. Food Prot. 1993, 56, 406–409. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Kamel, C. Plant Extracts Affect In Vitro Rumen Microbial Fermentation. J. Dairy Sci. 2006, 89, 761–771. [Google Scholar] [CrossRef]
- Vyas, D.; McGinn, S.M.; Duval, S.M.; Kindermann, M.K.; Beauchemin, K.A. Optimal Dose of 3-Nitrooxypropanol for Decreasing Enteric Methane Emissions from Beef Cattle Fed High-Forage and High-Grain Diets. Anim. Prod. Sci. 2018, 58, 1049–1055. [Google Scholar] [CrossRef]
- Haisan, J.; Sun, Y.; Guan, L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Kindermann, M.; Barreda, D.R.; Oba, M. The Effects of Feeding 3-Nitrooxypropanol at Two Doses on Milk Production, Rumen Fermentation, Plasma Metabolites, Nutrient Digestibility, and Methane Emissions in Lactating Holstein Cows. Anim. Prod. Sci. 2017, 57, 282–289. [Google Scholar] [CrossRef]
- Haisan, J.; Sun, Y.; Guan, L.L.; Beauchemin, K.A.; Iwaasa, A.; Duval, S.; Barreda, D.R.; Oba, M. The Effects of Feeding 3-Nitrooxypropanol on Methane Emissions and Productivity of Holstein Cows in Mid Lactation. J. Dairy Sci. 2014, 97, 3110–3119. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Humphries, D.J.; Kirton, P.; Kindermann, M.; Duval, S.; Steinberg, W. Effects of 3-Nitrooxypropanol on Methane Emission, Digestion, and Energy and Nitrogen Balance of Lactating Dairy Cows. J. Dairy Sci. 2014, 97, 3777–3789. [Google Scholar] [CrossRef]
- Melgar, A.; Harper, M.T.; Oh, J.; Giallongo, F.; Young, M.E.; Ott, T.L.; Duval, S.; Hristov, A.N. Effects of 3-Nitrooxypropanol on Rumen Fermentation, Lactational Performance, and Resumption of Ovarian Cyclicity in Dairy Cows. J. Dairy Sci. 2020, 103, 410–432. [Google Scholar] [CrossRef]
- Lopes, J.C.; de Matos, L.F.; Harper, M.T.; Giallongo, F.; Oh, J.; Gruen, D.; Ono, S.; Kindermann, M.; Duval, S.; Hristov, A.N. Effect of 3-Nitrooxypropanol on Methane and Hydrogen Emissions, Methane Isotopic Signature, and Ruminal Fermentation in Dairy Cows. J. Dairy Sci. 2016, 99, 5335–5344. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.O.; et al. An Inhibitor Persistently Decreased Enteric Methane Emission from Dairy Cows with No Negative Effect on Milk Production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef]
- Melgar, A.; Welter, K.C.; Nedelkov, K.; Martins, C.M.M.R.; Harper, M.T.; Oh, J.; Räisänen, S.E.; Chen, X.; Cueva, S.F.; Duval, S.; et al. Dose-Response Effect of 3-Nitrooxypropanol on Enteric Methane Emissions in Dairy Cows. J. Dairy Sci. 2020, 103, 6145–6156. [Google Scholar] [CrossRef]
- Elsevier Scopus—Document Search. Available online: https://www-scopus-com.cit.idm.oclc.org/search/form.uri?display=basic#basic (accessed on 11 February 2021).
- Wallace, B.; Dahabreh, I.; Trikalinos, T.; Lau, J.; Trow, P.; Schmid, C. Closing the Gap between Methodologists and End-Users: R as a Computational Back-End. J Stat Softw. 2012, 49, 1–15. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Bitsie, B.; Osorio, A.M.; Henry, D.D.; Silva, B.C.; Godoi, L.A.; Supapong, C.; Brand, T.; Schoonmaker, J.P. Enteric Methane Emissions, Growth, and Carcass Characteristics of Feedlot Steers Fed a Garlic-and Citrus-Based Feed Additive in Diets with Three Different Forage Concentrations. J. Anim. Sci. 2022, 100, 1–11. [Google Scholar] [CrossRef]
- Roque, B.M.; Van Lingen, H.J.; Vrancken, H.; Kebreab, E. Effect of Mootral—A Garlic- And Citrus-Extract-Based Feed Additive—And Enteric Methane Emissions in Feedlot Cattle. Transl. Anim. Sci. 2019, 3, 1383–1388. [Google Scholar] [CrossRef]
- Klop, G.; Hatew, B.; Bannink, A.; Dijkstra, J. Feeding Nitrate and Docosahexaenoic Acid Affects Enteric Methane Production and Milk Fatty Acid Composition in Lactating Dairy Cows. J. Dairy Sci. 2016, 99, 1161–1172. [Google Scholar] [CrossRef]
- van Wyngaard, J.D.V.; Meeske, R.; Erasmus, L.J. Effect of Dietary Nitrate on Enteric Methane Emissions, Production Performance and Rumen Fermentation of Dairy Cows Grazing Kikuyu-Dominant Pasture during Summer. Anim. Feed Sci. Technol. 2018, 244, 76–87. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.J. Effects and Mode of Action of Chitosan and Ivy Fruit Saponins on the Microbiome, Fermentation and Methanogenesis in the Rumen Simulation Technique. FEMS Microbiol. Ecol. 2016, 92, 160. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, H.; Wang, J.; Liu, Y.; Mao, S.; Xiong, B.; Jiang, L. Effects of Different Molecular Weights of Chitosan on Methane Production and Bacterial Community Structure in Vitro. J. Integr. Agric. 2020, 19, 1644–1655. [Google Scholar] [CrossRef]
- Melgar, A.; Lage, C.F.A.; Nedelkov, K.; Räisänen, S.E.; Stefenoni, H.; Fetter, M.E.; Chen, X.; Oh, J.; Duval, S.; Kindermann, M.; et al. Enteric Methane Emission, Milk Production, and Composition of Dairy Cows Fed 3-Nitrooxypropanol. J. Dairy Sci. 2021, 104, 357–366. [Google Scholar] [CrossRef]
- Yanibada, B.; Hohenester, U.; Pétéra, M.; Canlet, C.; Durand, S.; Jourdan, F.; Boccard, J.; Martin, C.; Eugène, M.; Morgavi, D.P.; et al. Inhibition of Enteric Methanogenesis in Dairy Cows Induces Changes in Plasma Metabolome Highlighting Metabolic Shifts and Potential Markers of Emission. Sci. Rep. 2020, 10, 15591. [Google Scholar] [CrossRef]
- Bikker, P.; Stokvis, L.; van Krimpen, M.M.; van Wikselaar, P.G.; Cone, J.W. Evaluation of Seaweeds from Marine Waters in Northwestern Europe for Application in Animal Nutrition. Anim. Feed Sci. Technol. 2020, 263, 114460. [Google Scholar] [CrossRef]
- Belanche, A.; Jones, E.; Parveen, I.; Newbold, C.J. A Metagenomics Approach to Evaluate the Impact of Dietary Supplementation with Ascophyllum Nodosum or Laminaria Digitata on Rumen Function in Rusitec Fermenters. Front. Microbiol. 2016, 7, 299. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S. Methane Mitigation from Ruminants Using Tannins and Saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef]
- Tan, H.Y.; Sieo, C.C.; Abdullah, N.; Liang, J.B.; Huang, X.D.; Ho, Y.W. Effects of Condensed Tannins from Leucaena on Methane Production, Rumen Fermentation and Populations of Methanogens and Protozoa in Vitro. Anim. Feed Sci. Technol. 2011, 169, 185–193. [Google Scholar] [CrossRef]
- Alvarez-Hess, P.S.; Jacobs, J.L.; Kinley, R.D.; Roque, B.M.; Neachtain, A.S.O.; Chandra, S.; Williams, S.R.O. Twice Daily Feeding of Canola Oil Steeped with Asparagopsis Armata Reduced Methane Emissions of Lactating Dairy Cows. Anim. Feed Sci. Technol. 2023, 297, 115579. [Google Scholar] [CrossRef]
- Kinley, R.D.; De Nys, R.; Vucko, M.J.; MacHado, L.; Tomkins, N.W. The Red Macroalgae Asparagopsis Taxiformis Is a Potent Natural Antimethanogenic That Reduces Methane Production during in Vitro Fermentation with Rumen Fluid. In Proceedings of the Animal Production Science; CSIRO: Canberra, Australia, 2016; Volume 56, pp. 282–289. [Google Scholar]
- Ellis, J.L.; Bannink, A.; Hindrichsen, I.K.; Kinley, R.D.; Pellikaan, W.F.; Milora, N.; Dijkstra, J. The Effect of Lactic Acid Bacteria Included as a Probiotic or Silage Inoculant on in Vitro Rumen Digestibility, Total Gas and Methane Production. Anim. Feed Sci. Technol. 2016, 211, 61–74. [Google Scholar] [CrossRef]
- Monteiro, H.F.; Laura, A.; Lelis, J.; Brandao, V.L.N.; Faccenda, A.; Avila, A.S.; Arce-Cordero, J.; Silva, L.G.; Dai, X.; Restelatto, R.; et al. In Vitro Evaluation of Lactobacillus Plantarum as Direct-Fed Microbials in High-Producing Dairy Cows Diets. Transl. Anim. Sci 2020, 4, 214–228. [Google Scholar] [CrossRef]
- O’Brien, M.; Hashimoto, T.; Senda, A.; Nishida, T.; Takahashi, J. The Impact of Lactobacillus Plantarum TUA1490L Supernatant Oninvitro Rumen Methanogenesis and Fermentation. Anaerobe 2013, 22, 137–140. [Google Scholar] [CrossRef]
- McCauley, J.I.; Labeeuw, L.; Jaramillo-Madrid, A.C.; Nguyen, L.N.; Nghiem, L.D.; Chaves, A.V.; Ralph, P.J. Management of Enteric Methanogenesis in Ruminants by Algal-Derived Feed Additives. Curr. Pollut. Rep. 2020, 6, 188–205. [Google Scholar] [CrossRef]
- Haque, M.N. Dietary Manipulation: A Sustainable Way to Mitigate Methane Emissions from Ruminants. J. Anim. Sci. Technol. 2018, 60, 15. [Google Scholar] [CrossRef]
- Kebreab, E.; Bannink, A.; Pressman, E.M.; Walker, N.; Karagiannis, A.; van Gastelen, S.; Dijkstra, J. A Meta-Analysis of Effects of 3-Nitrooxypropanol on Methane Production, Yield, and Intensity in Dairy Cattle. J. Dairy Sci. 2023, 106, 927–936. [Google Scholar] [CrossRef]
- Almeida, A.K.; Hegarty, R.S.; Cowie, A. Meta-Analysis Quantifying the Potential of Dietary Additives and Rumen Modifiers for Methane Mitigation in Ruminant Production Systems. Anim. Nutr. 2021, 7, 1219–1230. [Google Scholar] [CrossRef]
- Patra, A.K. Enteric Methane Mitigation Technologies for Ruminant Livestock: A Synthesis of Current Research and Future Directions. Environ. Monit. Assess. 2012, 184, 1929–1952. [Google Scholar] [CrossRef]
- Manzanilla-Pech, C.I.V.; Stephansen, R.B.; Difford, G.F.; Løvendahl, P.; Lassen, J. Selecting for Feed Efficient Cows Will Help to Reduce Methane Gas Emissions. Front. Genet. 2022, 13, 885932. [Google Scholar] [CrossRef]
- Weimer, P.J. Degradation of Cellulose and Hemicellulose by Ruminal Microorganisms. Microorganisms 2022, 10, 2345. [Google Scholar] [CrossRef]
- Czerkawski, J.W.; Breckenridge, G. Design and Development of a Long-Term Rumen Simulation Technique (Rusitec). Br. J. Nutr. 1977, 38, 371–384. [Google Scholar] [CrossRef]
- Ma, T.; Chen, D.; Tu, Y.; Zhang, N.; Si, B.; Deng, K.; Diao, Q. Effect of Supplementation of Allicin on Methanogenesis and Ruminal Microbial Flora in Dorper Crossbred Ewes. J. Anim. Sci. Biotechnol. 2016, 7, 1. [Google Scholar] [CrossRef]
- Romero-Perez, A.; Okine, E.K.; McGinn, S.M.; Guan, L.L.; Oba, M.; Duval, S.M.; Kindermann, M.; Beauchemin, K.A. Sustained Reduction in Methane Production from Long-Term Addition of 3-Nitrooxypropanol to a Beef Cattle Diet. J. Anim. Sci. 2015, 93, 1780–1791. [Google Scholar] [CrossRef]
- Seo, S.; Lanzas, C.; Tedeschi, L.O.; Fox, D.G. Development of a Mechanistic Model to Represent the Dynamics of Liquid Flow out of the Rumen and to Predict the Rate of Passage of Liquid in Dairy Cattle. J. Dairy Sci. 2007, 90, 840–855. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Q.; Lan, X.; Shen, W.; Wan, F.; He, J.; Tang, S.; Tan, Z. Evaluation of Stirring Time through a Rumen Simulation Technique: Influences on Rumen Fermentation and Bacterial Community. Front. Microbiol. 2023, 14, 1103222. [Google Scholar] [CrossRef] [PubMed]
- Sudarshan, N.R.; Hoover, D.G.; Knorr, D. Antibacterial Action of Chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Allison, M.; Reddy, C. Adaptations of Gastrointestinal Bacteria in Response to Changes in Dietary Oxalate and Nitrate. In Proceedings of the Current Perspectives in Microbial Ecology—Proceedings of the Third International Symposium on Microbial Ecology, Michigan State University, East Lansing, MI, USA, 7–12 August 1983; American Society for Microbiology: Washington, DC, USA, 1984; pp. 248–256. [Google Scholar]
- Patra, A.K.; Saxena, J. A New Perspective on the Use of Plant Secondary Metabolites to Inhibit Methanogenesis in the Rumen. Phytochemistry 2010, 71, 1198–1222. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, F.M.; Williams, P.; Losa, R.; Wallace, R.J.; Beever, D.A.; Newbold, C.J. Effects of Essential Oils on Ruminal Microorganisms and Their Protein Metabolism. Appl. Environ. Microbiol. 2003, 69, 5011–5014. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Brand, T.; Tapio, I.; Bayat, A.R. Effect of a Garlic and Citrus Extract Supplement on Performance, Rumen Fermentation, Methane Production, and Rumen Microbiome of Dairy Cows. J. Dairy Sci. 2023, 106, 4608–4621. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Davis, C.L.; Frobish, R.A.; Sachan, D.S. Evaluation of Polyethylene Glycol Method in Determining Rumen Fluid Volume in Dairy Cows Fed Different Diets. J. Dairy Sci. 1971, 54, 928–930. [Google Scholar] [CrossRef]
- Xie, B.; Yang, X.; Yang, L.; Wen, X.; Zhao, G. Adding Polyethylene Glycol to Steer Ration Containing Sorghum Tannins Increases Crude Protein Digestibility and Shifts Nitrogen Excretion from Feces to Urine. Anim. Nutr. 2021, 7, 779–786. [Google Scholar] [CrossRef]
- Silanikove, N.; Perevolotsky, A.; Provenza, F.D. Use of Tannin-Binding Chemicals to Assay for Tannins and Their Negative Postingestive Effects in Ruminants. Anim. Feed Sci. Technol. 2001, 91, 69–81. [Google Scholar] [CrossRef]









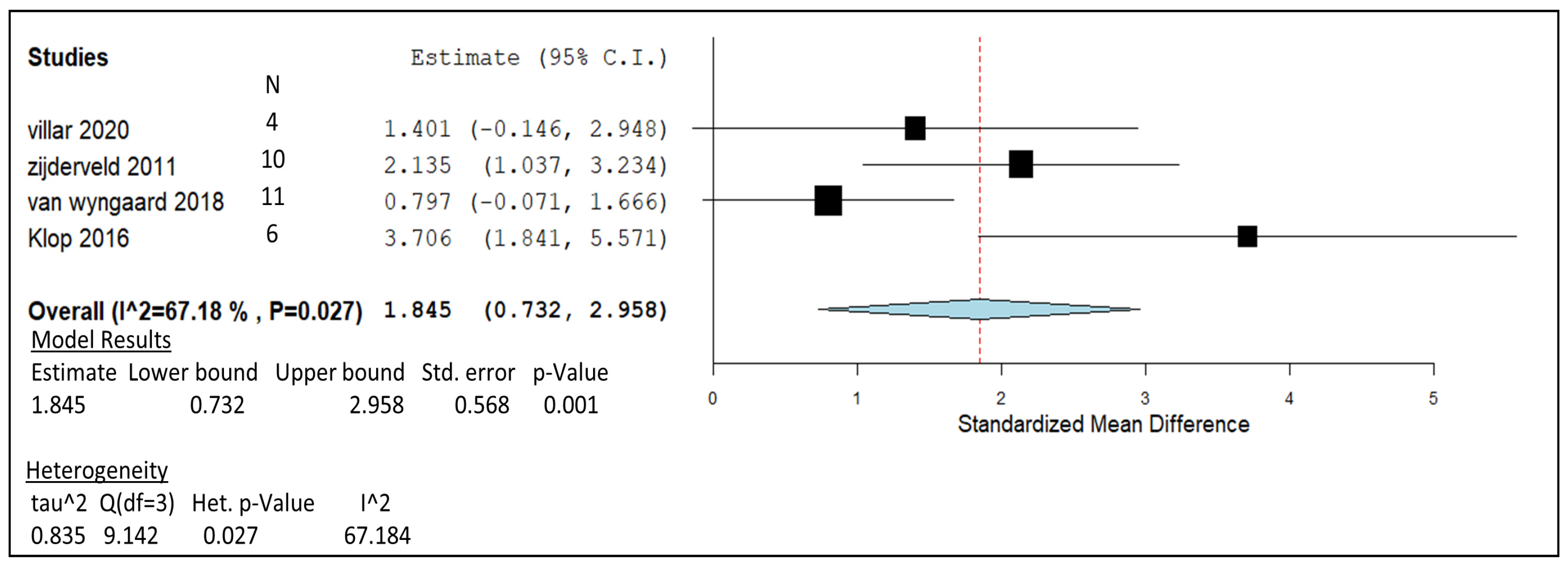
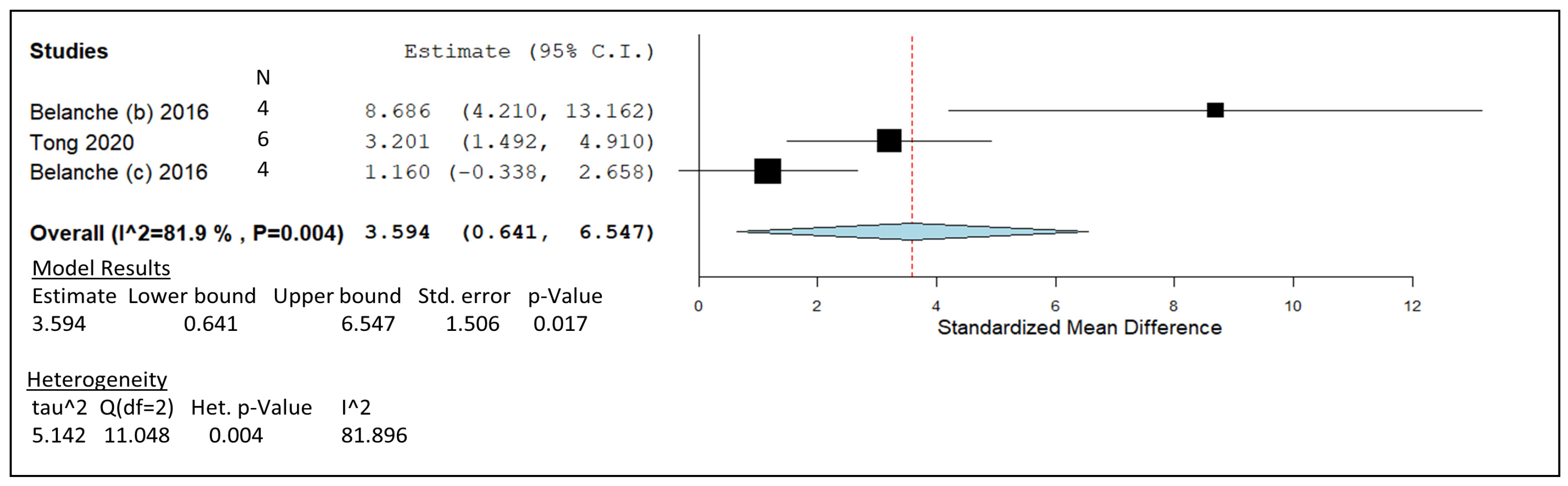
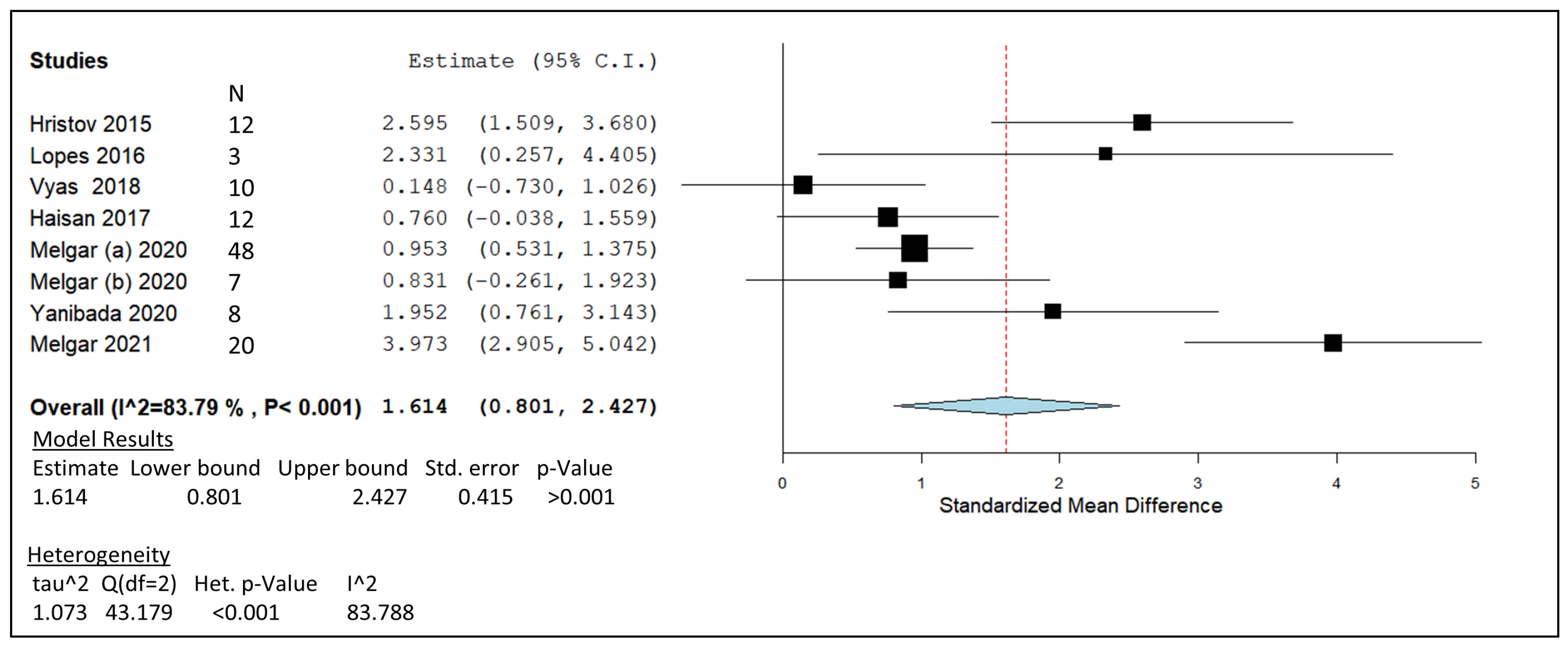
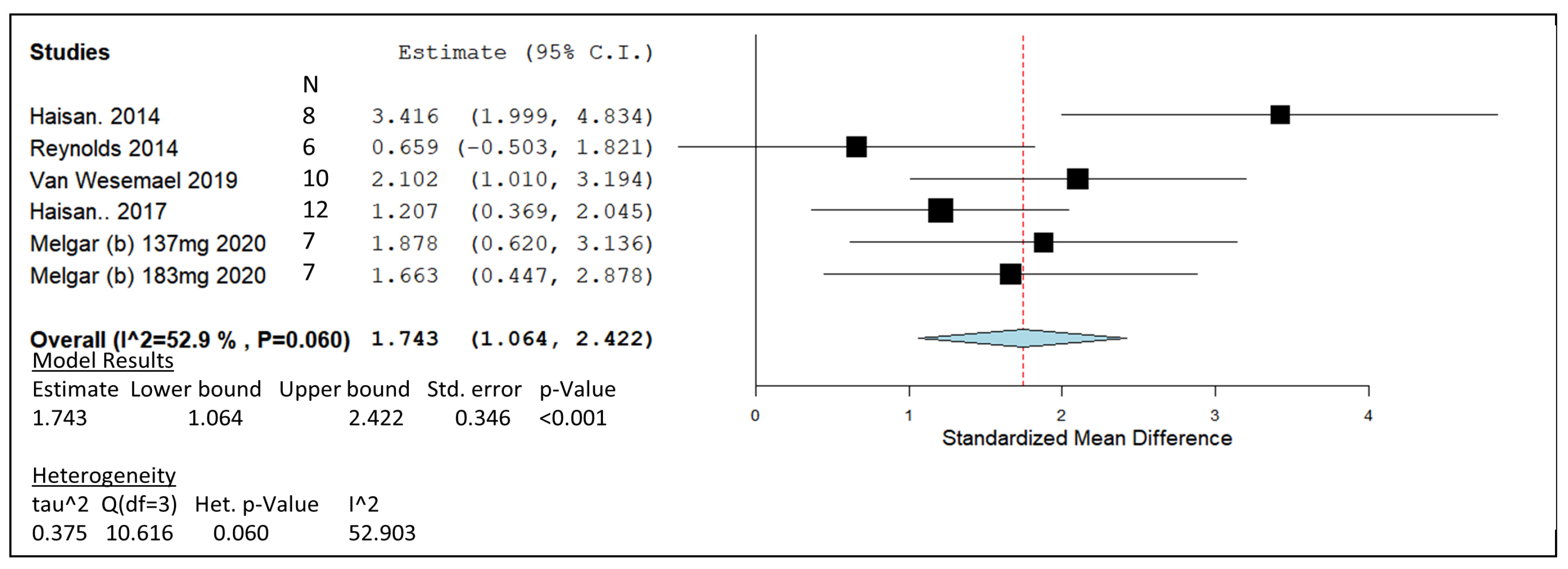
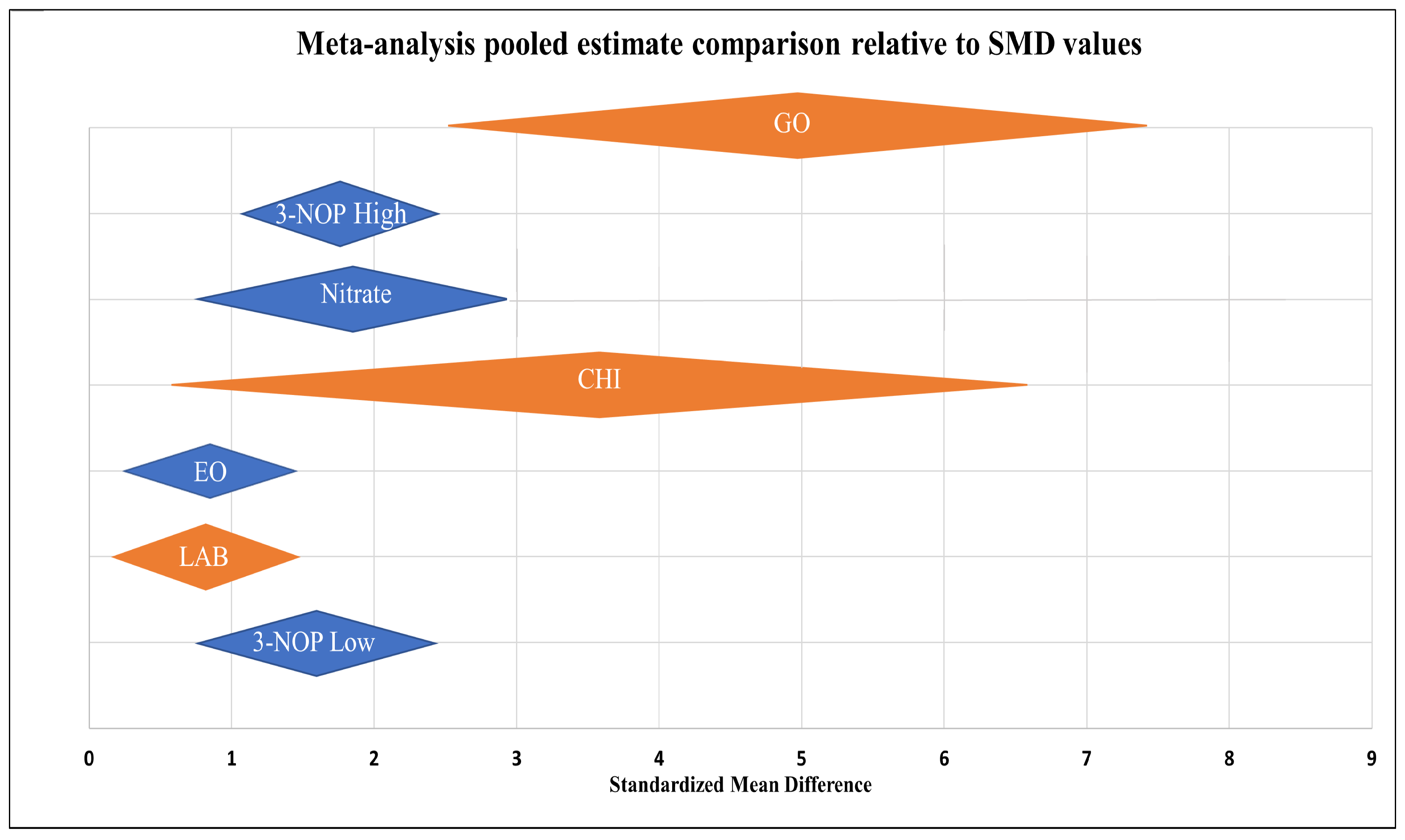
 ) including garlic oil (GO), L. plantarum (LAB), chi-tosan (CHI), and in vivo studies identified by blue diamonds (
) including garlic oil (GO), L. plantarum (LAB), chi-tosan (CHI), and in vivo studies identified by blue diamonds ( ) including nitrate, essential oil blends (EOs), and 3-nitrooxypropanol at high (3-NOP High), and low doses (3-NOP Low).
) including garlic oil (GO), L. plantarum (LAB), chi-tosan (CHI), and in vivo studies identified by blue diamonds () including nitrate, essential oil blends (EOs), and 3-nitrooxypropanol at high (3-NOP High), and low doses (3-NOP Low).
) including nitrate, essential oil blends (EOs), and 3-nitrooxypropanol at high (3-NOP High), and low doses (3-NOP Low).
) including garlic oil (GO), L. plantarum (LAB), chi-tosan (CHI), and in vivo studies identified by blue diamonds () including nitrate, essential oil blends (EOs), and 3-nitrooxypropanol at high (3-NOP High), and low doses (3-NOP Low).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Search Number | Scopus Search Terms | Relevant (Total) Hits | Date |
|---|---|---|---|
| 1 | “Methane abatement” | too broad | . |
| 2 | “Methane abatement” AND “Rumen” | 7 (19) | 2007–2023 |
| 3 | “Methane reduction” AND “Rumen” | 6 (175) | 2011–2023 |
| 4 | “Methane production” AND “Rumen” | 19 (507) | 1997–2023 |
| 5 | “Methane production” AND “Rumen” AND “weight Gain” | 2 (9) | 2017–2018 |
| 6 | “Brown seaweed” AND “Rumen” | 7 (8) | 2015–2019 |
| 7 | “Agolin” AND “Rumen” | 6 (14) | 1948–2023 |
| 8 | “Garlic” AND “Citrus” AND “Rumen” | 4 (10) | 2018–2022 |
| 9 | “3-nitrooxypropanol” | 9 (15) | 2014–2023 |
| 10 | “Ascophyllum nodosum” AND “Rumen” | 7 (16) | 2004–2020 |
| 11 | “Laminaria digitata” AND “Rumen” | 1 (8) | 2018–2018 |
| 12 | ” Garlic oil” AND “Methane” | 12 (47) | 2005–2018 |
| 13 | “Lactic acid bacteria” AND “Methane” | 11 (19) | 1988–2020 |
| 14 | “Nitrate” AND “methane” AND “Rumen” | 11 (149) | 1972–2022 |
| 15 | “Subacute rumen acidosis” AND “Rumen” | 3 (240) | 2007–2008 |
| 16 | “Chitosan” AND “Rumen” | 9 (37) | 1998–2020 |
| 17 | “Asparagopsis” AND “Rumen” | 11 (19) | 2006–2023 |
| GO Study | Ruminant System | Concentration 3 | In Vitro/In Vivo 2 | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Soliva, 2011 | Lactating brown Swiss cow | 300 mg/L CF | In vitro (RUSITEC) | -GO caused almost complete inhibition of CH4. -Decreased protozoal numbers and increased bacterial counts. | 4 |
| Patra, 2012 | Lactating Jersey cow | 250 mg/L CF | In vitro | -Reduced CH4 emissions by around 23%. -Dose inhibitory to Archaea but did not affect feed digestibility. | 3 |
| Patra, 2015 | Lactating Jersey cow | 250 mg/L CF | In vitro | -Total VFA concentration decreased. -Suppressed enteric methane by 40% after 18 days. | 3 |
| Chaves, 2008 | Lactating Holstein cow | 250 mg/L CF | In vitro | -CH4 emissions from ruminal bacteria reduced 72%. -Proportion of propionate reduced. | 3 |
| Nitrate Study | Ruminant System | Concentration Given 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Villar, 2020 | Steers | 20 g/kg DM | in vivo | -Rumen protozoal concentration was reduced when including NO3. -Substantial decrease in CH4 peaks after 8 h in respiration chambers. | 4 |
| Van Zijderveld, 2011 | Holstein–Friesian dairy cows | 21 g/kg DM | in vivo | -Nitrate decreased methane production by 16%. -Milk protein content lowered and increased hydrogen emission. | 10 |
| Van Wyngaard, 2018 | Jersey cows | 23 g/kg DM | in vivo | -Milk yields decreased by 12%; concentrate DMI decreased linearly (5.5–3.7 kg/d). -CH4 production decreased linearly with increasing nitrate addition. | 11 |
| Klop, 2016 | Holstein dairy cows | 21 g/kg DM | in vivo | -Decreased enteric CH4 production by 23%. -Increased polyunsaturated fats and lower milk protein concentration. | 6 |
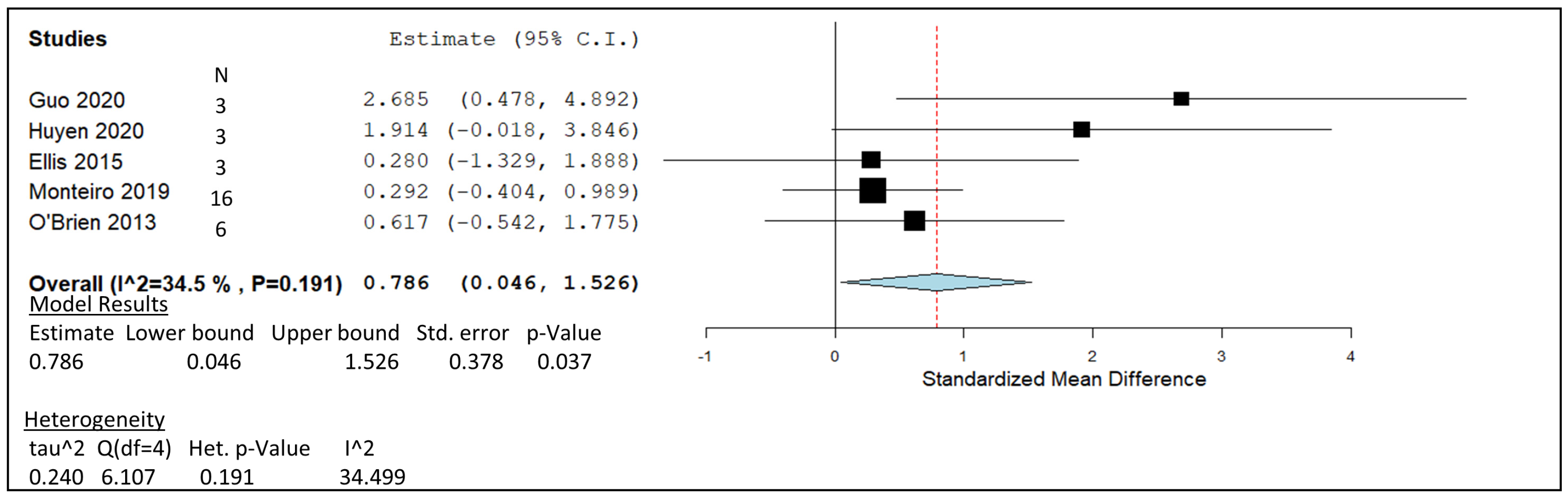
| LAB Study | Ruminant System | Concentration 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Guo, 2020 | Jinnan cattle | 1.0 × 106 cfu/g (fresh weight) | in vitro | -Lowered the ratio of CH4 output to total VFAs. -Increased acetate and propionate, total VFA, DM-D, and NDF-D as compared with that of the control after 72 h in vitro incubation. | 3 |
| Huyen, 2020 | Lactating Holstein–Friesian cow | 1.0 × 106 cfu/g (fresh weight) | in vitro | -CH4 production was lower for LAB when used as silage inoculants, compared to being used as directly fed microbials. -Increased the in vitro DM and organic matter (OM) degradability both in the fresh ration and rain treated ration. | 3 |
| Ellis, 2016 | Holstein–Friesian | ‘1.0 × 106 cfu/mL (In 60 mL of buffered rumen fluid) | in vitro | -No significant effect of LAB treatment on OM digestibility, cumulative gas or CH4 production. | 3 |
| Monteiro, 2020 | Lactating Holstein cow | 1.35 × 109 cfu/g DM | in vitro | -No significant changes on CH4 production. -Lower CO2 production linked with total VFA reduction over 24 and 48 h. | 16 |
| O’Brien, 2013 | Holstein cow | ‘1.0 × 108.3 cfu/mL (In 100 mL glucose-yeast medium) | in vitro | -Molar proportions of propionic acid increase and lower levels of acetic and butyric acid. -Decrease in total VFA concentration (17%) (mM) and CH4 output (68%) (ml 24 h−1). | 6 |
| Chitosan Study | Ruminant System | Concentration Given 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Belanche, 2016b | Holstein–Friesian | 50 mg/g DM | In vitro (RUSITEC) | -Decreased rumen methanogenesis by 42%. -Promoted shift in fermentation pattern towards propionate production. | 4 |
| Tong, 2020 | Lactating Holstein cow | 16 mg/g DM | In vitro | -Propionate concentration was significantly increased, and acetate proportion was decreased. -CH4 reduced by replacing fibrolytic bacteria with amylolytic bacteria. | 6 |
| Belanche, 2016c | Holstein–Friesian | 50 mg/g DM | In vitro | -Fermentation shifted towards propionate production. -Lower CH4 (23%) and protozoal activity (56%). | 4 |
| AR/MO Ruminant Study | Ruminant System | Concentration Given 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Klop, Vaan Laar-Van Shuppen, 2017b | Lactating Holstein cow | 0.05 g/kg DM | in vivo | -Average CH4 production was decreased by 8% when AR was supplemented to associated doner animals for 3 weeks. -No negative effects on dietary mass intake for cows receiving AR diet. | 9 |
| Klop, Dijkstra, 2017a | Lactating dairy cows | 0.17 g/kg DM | In vivo | -CH4 production lowered after first period of 2 weeks only. -Higher proportions of acetate and propionate with AR supplementation. | 4 |
| Hart, 2019 | Holstein–Friesian cows | 0.05 g/kg DM | In vivo | -Yields of milk fat, protein, lactose, and solids were higher for AR-fed cows. -CH4 output was reduced by 27 g/day with AR compared to control treatment. | 73 |
| Castro Montoya, 2015 | Lactating Holstein cow | 0.05 g/kg DM | In vivo | -Milk production displayed a linear decrease towards the end of the study. -Addition of AR accounted for 15% (g/d) decrease in CH4 over the experimental period. | 4 |
| Carrazco, 2020 | Lactating Holstein cow | 0.04 g/kg DM | In vivo | -CH4 yield was not significantly reduced by essential oil blend. -Ruminant production parameters did not differ with AR supplementation. | 10 |
| Bach, 2023 | Lactating Holstein cow | 0.04 g/kg DM | In vivo | -AR supplementation decreased CH4 yield (L/kg DM) by 12.3%. -Feed efficiency (ECM/DMI) and diversity of microbiome was lower with AR supplemented cows. | 20 |
| Roque, 2019 | Angus x Hereford steers | 1.6 g/kg DM | In vivo | -CH4 yield decreased by 13.3% with MO supplementation. -DMI, ADG, and feed efficiency remained unchanged with essential oil blend supplementation. | 10 |
| Bitsie, 2022 | Angus x Simmental steers | 2.5 g/kg DM | In vivo | -CH4 yield (g/kg DMI) decreased by 24.6% with MO. -MO increased DM, CP, and ADF apparent digestibility. | 72 |
| 3-NOP Low-Dose Study | Ruminant System | Concentration Given 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Hristov, 2015 | Lactating Holstein cow | 60 mg/kg DM | in vivo | -Milk protein and lactose increased by 3-NOP. -Increased body weight. -CH4 emissions decreased by 30% lower than control. | 12 |
| Lopes, 2016 | Lactating Holstein cow | 60 mg/kg DM | In vivo | -Acetate to propionate ratio was lower when treated with 3-NOP. -Proportions of methanogens decreased. -Inhibition of enteric methane with increased hydrogen emissions. | 3 |
| Vyas, 2018 | Crossbred steers | 75 mg/kg DM | In vivo | -Lowered total CH4 emissions with increased 3-NOP supplementation. -No negative effects on DMI. | 10 |
| Haisan, 2017 | Lactating Holstein cow | 68 mg/kg DM | In vivo | -Molar proportions of acetate to propionate reduced in dose dependant manner. -CH4 yield (g/kg DM) decreased by 23%. | 12 |
| Melgar, 2020a | Lactating Holstein cow | 60 mg/kg DM | In vivo | -3-NOP decreased CH4 yield by 21% over a 15-week treatment period. -Hydrogen emissions were increase by 48-fold. | 48 |
| Melgar, 2020b | Lactating Holstein cow | 73 mg/kg DM | In vivo | -Hydrogen emissions increase over 7-fold with 3-NOP inclusion. -CH4 yield decreased by 16% g/kg DM. | 7 |
| Yanibada, 2020 | Lactating Holstein cow | 60 mg/kg DM | In vivo | -CH4 yield mitigated by 21.7% over 5-week treatment. -No changes in milk yield or composition. | 8 |
| Melgar, 2021 | Lactating Holstein cow | 60 mg/kg DM | In vivo | -3-NOP decreased CH4 yield by 27% compared to the control. -Hydrogen emissions were increased 6-fold with 3-NOP inclusion. -Increased milk fat yield with treatment. | 20 |
| 3-NOP High Dose Study | Ruminant System | Concentration Given 2 | In Vitro/In Vivo | Overall Effect 1 | N |
|---|---|---|---|---|---|
| Haisan 2014 | Holstein lactating | 126.9 mg/kg DM | In vivo | -CH4 production was reduced by 10.62 g/kg DMI. -Cattle fed 3-NOP gained more body weight. -Reduction in acetate to propionate ratio was observed. | 11 |
| Reynolds 2014 | Holstein-Friesian cows | 135.1 mg/kg DM | In vivo | -Decrease in acetate to propionate ratio. -Dry matter, organic matter, acid detergent fibre and energy digestibility were reduced. | 4 |
| Van Wesemael 2019 | Holstein-Friesian lactating | 100 mg/kg DM | In vivo | -CH4 production was 28% lower for basal diet treated with 3-NOP and 23% lower for concentrate diets. | 6 |
| Haisan 2017 | Holstein lactating | 132 mg/kg DM | In vivo | -CH4 yield (g/kg DM) was decreased by 36.7% with high 3-NOP. -Apparent total-tract digestibility significantly increased with 3-NOP. | 12 |
| Melgar 2020b | Holsten lactating | 137 mg/kg DM + 183 mg/kg DM | In vivo | -CH4 yield decreased by 36% and 31.8% with 137 and 183 mg/kg DM 3-NOP, respectively. -Hydrogen emission production increased 9-fold and 7-fold with 137 and 183 mg/kg DM 3-NOP, respectively. -Milk fat % increased with 3-NOP supplementation. | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hodge, I.; Quille, P.; O’Connell, S. A Review of Potential Feed Additives Intended for Carbon Footprint Reduction through Methane Abatement in Dairy Cattle. Animals 2024, 14, 568. https://doi.org/10.3390/ani14040568
Hodge I, Quille P, O’Connell S. A Review of Potential Feed Additives Intended for Carbon Footprint Reduction through Methane Abatement in Dairy Cattle. Animals. 2024; 14(4):568. https://doi.org/10.3390/ani14040568
Chicago/Turabian StyleHodge, Ian, Patrick Quille, and Shane O’Connell. 2024. "A Review of Potential Feed Additives Intended for Carbon Footprint Reduction through Methane Abatement in Dairy Cattle" Animals 14, no. 4: 568. https://doi.org/10.3390/ani14040568