
Molecular Characterization, Expression Pattern, DNA Methylation and Gene Disruption of Figla in Blotched Snakehead (Channa maculata)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Sampling
2.2. RNA Extraction and cDNA Synthesis
2.3. Full-Length cDNA Cloning and Sequence Analysis
2.4. Genome DNA Sequence Cloning and Structure Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. In Situ Hybridisation (ISH)
2.7. Bisulfite Sequencing PCR (BSP)
2.8. Knockout of Figla Gene in C. maculata
2.9. Statistical Analysis
3. Results
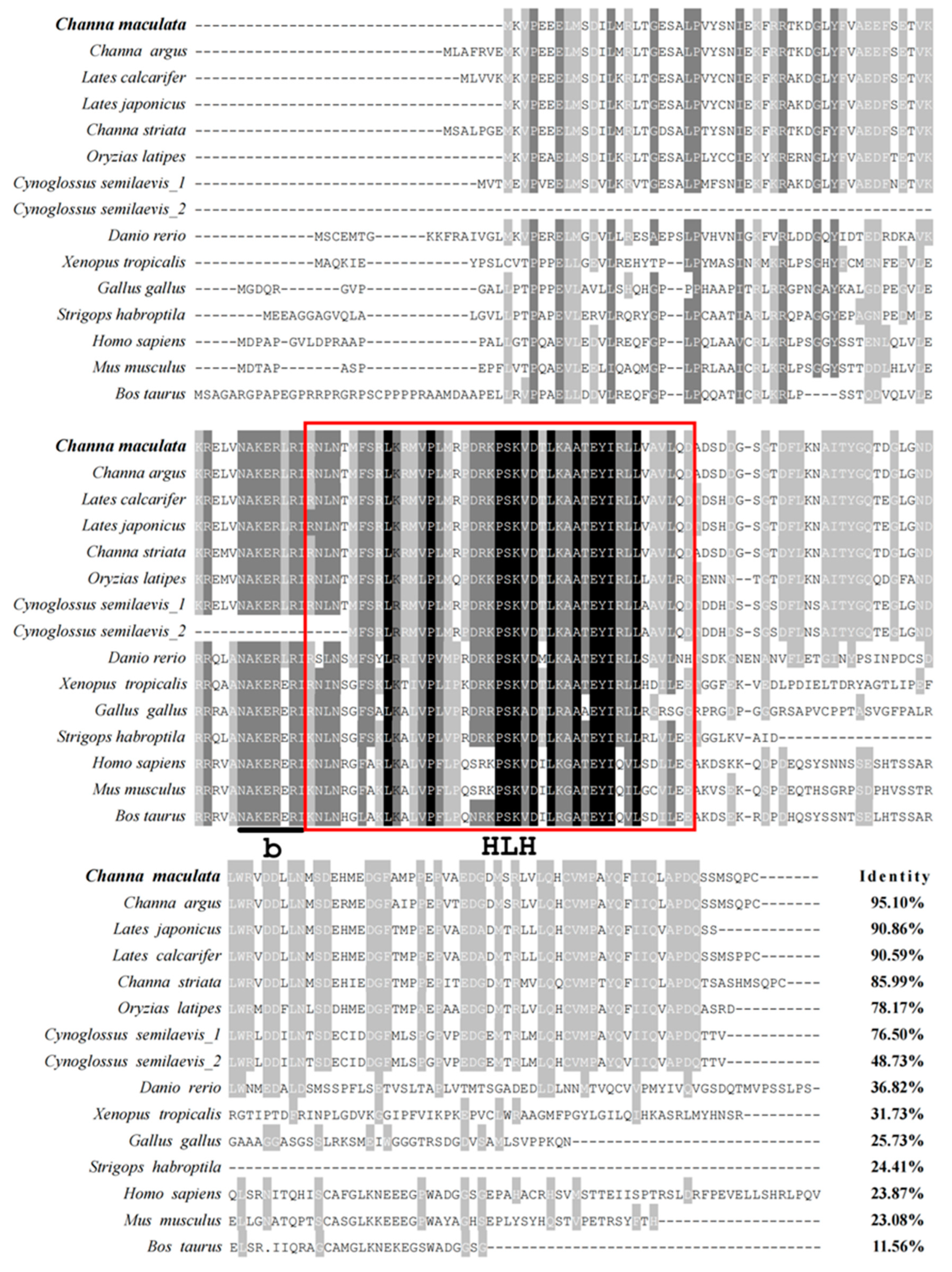
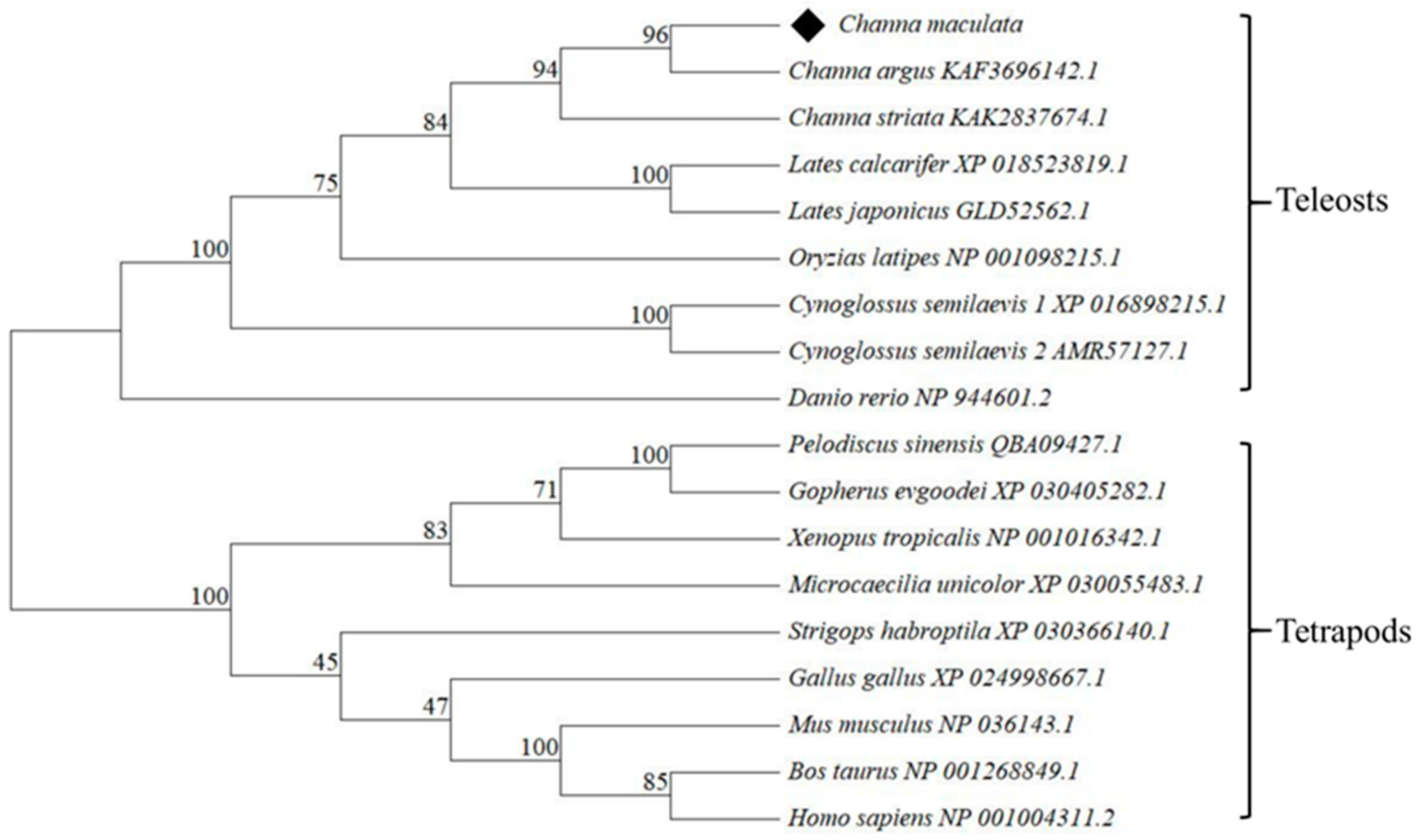
3.1. Sequence Analysis of Figla in C. maculata
3.2. Expression Patterns of CmFigla
3.3. CpG Methylation Levels of CmFigla Promoter
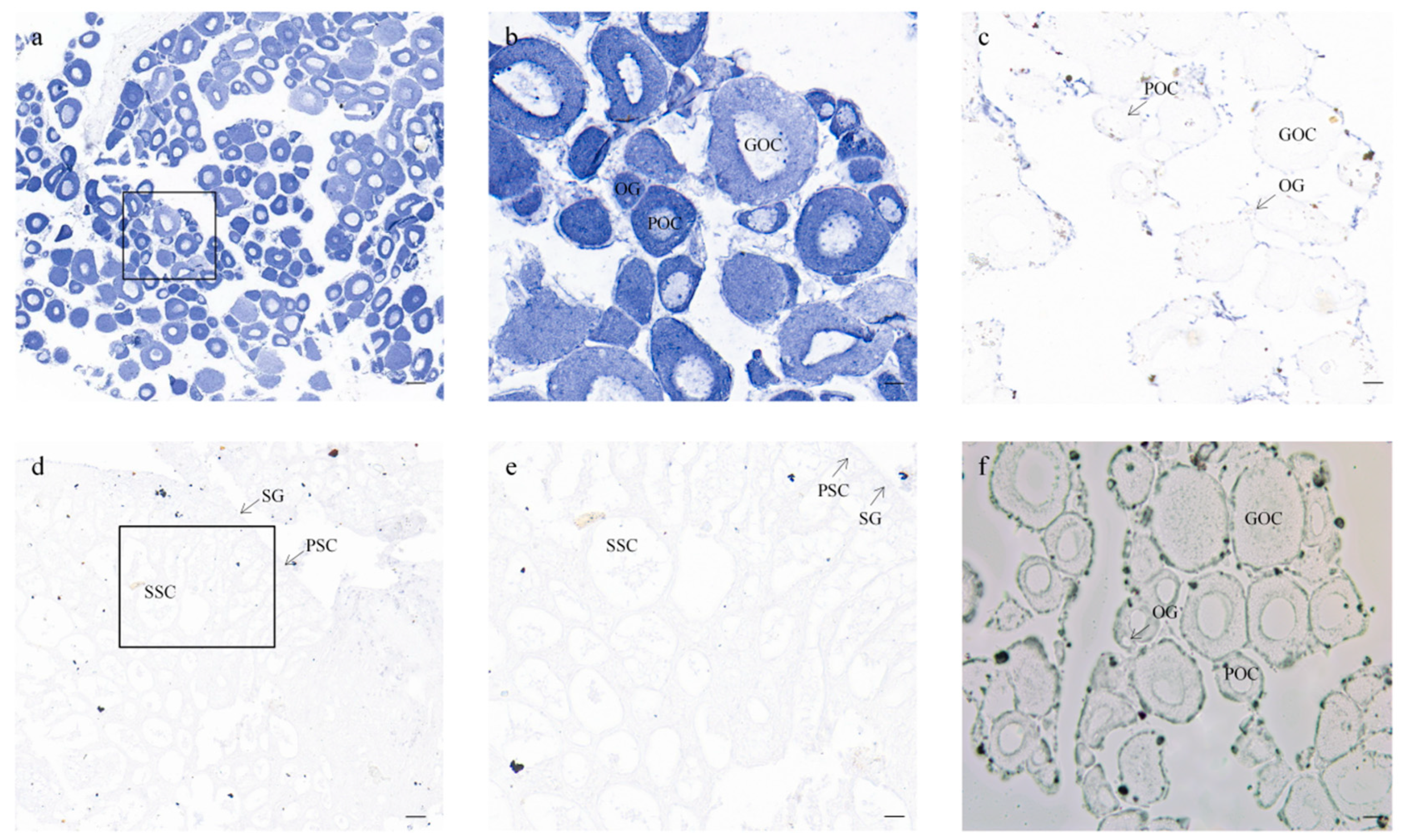
3.4. Cellular Location of CmFigla in Gonads
3.5. Generation of the CmFigla Mutantion Using the CRISPR/Cas9 System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tutukova, S.; Tarabykin, V.; Hernandez-Miranda, L.R. The role of neurod genes in brain development, function, and disease. Front. Mol. Neurosci. 2021, 14, 662774. [Google Scholar] [CrossRef] [PubMed]
- Li, A.-M.; He, W.-Z.; Wei, J.-L.; Chen, Z.-L.; Liao, F.; Qin, C.-X.; Pan, Y.-Q.; Shang, X.-K.; Lakshmanan, P.; Wang, M.; et al. Transcriptome profiling reveals genes related to sex determination and differentiation in Sugarcane borer (Chilo sacchariphagus Bojer). Insects 2022, 13, 500. [Google Scholar] [CrossRef]
- Mor, E.; Pernisová, M.; Minne, M.; Cerutti, G.; Ripper, D.; Nolf, J.; Andres, J.; Ragni, L.; Zurbriggen, M.D.; De Rybel, B.; et al. bHLH heterodimer complex variations regulate cell proliferation activity in the meristems of Arabidopsis thaliana. iScience 2022, 25, 105364. [Google Scholar] [CrossRef]
- Hamazaki, N.; Kyogoku, H.; Araki, H.; Miura, F.; Horikawa, C.; Hamada, N.; Shimamoto, S.; Hikabe, O.; Nakashima, K.; Kitajima, T.S.; et al. Reconstitution of the oocyte transcriptional network with transcription factors. Nature 2021, 589, 264–269. [Google Scholar] [CrossRef]
- Soyal, S.M.; Amleh, A.; Dean, J. FIGα, a germ cell-specific transcription factor required for ovarian follicle formation. Development 2000, 127, 4645–4654. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, C.-Y.; Zhao, Y.; Dean, J. FIGLA, LHX8 and SOHLH1 transcription factor networks regulate mouse oocyte growth and differentiation. Nucleic Acids Res. 2020, 48, 3525–3541. [Google Scholar] [CrossRef]
- Chen, M.; Jiang, H.; Zhang, C. Selected genetic factors associated with primary ovarian insufficiency. Int. J. Mol. Sci. 2023, 24, 4423. [Google Scholar] [CrossRef]
- Qiu, Y.; Sun, S.; Charkraborty, T.; Wu, L.; Sun, L.; Wei, J.; Nagahama, Y.; Wang, D.; Zhou, L. Figla favors ovarian differentiation by antagonizing spermatogenesis in a teleosts, Nile tilapia (Oreochromis niloticus). PLoS ONE 2015, 10, e0123900. [Google Scholar] [CrossRef]
- Qu, J.; Li, R.; Liu, Y.; Sun, M.; Yan, W.; Liu, J.; Wang, X.; Zhang, Q. Molecular characterization, expression pattern and transcriptional regulation of Figla during gonad development in Japanese founder (Paralichthys olivaceus). J. Ocean Univ. China 2022, 21, 1037–1050. [Google Scholar] [CrossRef]
- Miyake, Y.; Sakai, Y.; Kuniyoshi, H. Molecular cloning and expression profile of sex-specific genes, Figla and Dmrt1, in the protogynous hermaphroditic fish, Halichoeres poecilopterus. Zool. Sci. 2012, 29, 690. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-C.; Chang, C.-F. Oocytes survive in the testis by altering the soma fate from male to female in the protandrous black porgy, Acanthopagrus schlegeli. Biol. Reprod. 2013, 88, 19. [Google Scholar] [CrossRef]
- Qin, M.; Zhang, Z.; Song, W.; Wong, Q.W.-L.; Chen, W.; Shirgaonkar, N.; Ge, W. Roles of Figla/figla in juvenile ovary development and follicle formation during zebrafish gonadogenesis. Endocrinology 2018, 159, 3699–3722. [Google Scholar] [CrossRef]
- Shao, C.; Li, Q.; Chen, S.; Zhang, P.; Lian, J.; Hu, Q.; Sun, B.; Jin, L.; Liu, S.; Wang, Z.; et al. Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 2014, 24, 604–615. [Google Scholar] [CrossRef]
- Wang, W.; Tan, S.; Yang, Y.; Zhou, T.; Xing, D.; Su, B.; Wang, J.; Li, S.; Shang, M.; Gao, D.; et al. Feminization of channel catfish with 17β-oestradiol involves methylation and expression of a specific set of genes independent of the sex determination region. Epigenetics 2022, 17, 1820–1837. [Google Scholar] [CrossRef]
- Zhao, J.; Ou, M.; Wang, Y.; Liu, H.; Luo, Q.; Zhu, X.; Chen, B.; Chen, K. Breeding of YY super-male of blotched snakehead (Channa maculata) and production of all-male hybrid (Channa argus ♀ × C. maculata ♂). Aquaculture 2021, 538, 736450. [Google Scholar] [CrossRef]
- Ou, M.; Chen, K.; Gao, D.; Wu, Y.; Chen, Z.; Luo, Q.; Liu, H.; Zhao, J. Comparative transcriptome analysis on four types of gonadal tissues of blotched snakehead (Channa maculata). Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 35, 100708. [Google Scholar] [CrossRef]
- Huang, S.; Wu, Y.; Chen, K.; Zhang, X.; Zhao, J.; Luo, Q.; Liu, H.; Wang, F.; Li, K.; Fei, S.; et al. Gene expression and epigenetic modification of aromatase during sex reversal and gonadal development in blotched snakehead (Channa maculata). Fishes 2023, 8, 129. [Google Scholar] [CrossRef]
- Feng, Z.-F.; Zhang, Z.-F.; Shao, M.-Y.; Zhu, W. Developmental expression pattern of the Fc-vasa-like gene, gonadogenesis and development of germ cell in Chinese shrimp, Fenneropenaeus chinensis. Aquaculture 2011, 314, 202–209. [Google Scholar] [CrossRef]
- Gao, J.; Wang, J.; Jiang, J.; Fan, L.; Wang, W.; Liu, J.; Zhang, Q.; Wang, X. Identification and characterization of a nanog homolog in Japanese flounder (Paralichthys olivaceus). Gene 2013, 531, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.; Wang, F.; Li, K.; Wu, Y.; Huang, S.; Luo, Q.; Liu, H.; Zhang, X.; Fei, S.; Chen, K.; et al. Generation of myostatin gene-edited blotched snakehead (Channa maculata) using CRISPR/Cas9 system. Aquaculture 2023, 563, 738988. [Google Scholar] [CrossRef]
- Wu, K.; Zhai, Y.; Qin, M.; Zhao, C.; Ai, N.; He, J.; Ge, W. Genetic evidence for differential functions of Figla and Nobox in zebrafish ovarian differentiation and folliculogenesis. Commun. Biol. 2023, 6, 1185. [Google Scholar] [CrossRef]
- Liang, S.; Wang, W.; Wang, L.; Wu, Z.; Zou, Y.; Tan, X.; Liu, Y.; Peng, Z.; You, F. Figla gene roles in the proliferation of oocytes in the olive flounder Paralichthys olivaceus. Aquaculture 2020, 528, 735493. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhu, Y.; Chen, Z.; Wang, L.; Li, W.; Lu, S.; Chen, S. Identification and expression analysis of Figla gene in the turbot, Scophthalmus maximus. Aquac. Res. 2018, 49, 3483–3490. [Google Scholar] [CrossRef]
- Li, H.; Xu, W.; Zhang, N.; Shao, C.; Zhu, Y.; Dong, Z.; Wang, N.; Jia, X.; Xu, H.; Chen, S. Two Figla homologues have disparate functions during sex differentiation in half-smooth tongue sole (Cynoglossus semilaevis). Sci. Rep. 2016, 6, 28219. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-J.; Maugars, G.; Lafont, A.-G.; Jeng, S.-R.; Wu, G.-C.; Dufour, S.; Chang, C.-F. Basal teleosts provide new insights into the evolutionary history of teleost-duplicated aromatase. Gen. Comp. Endocrinol. 2020, 291, 113395. [Google Scholar] [CrossRef] [PubMed]
- Kleppe, L.; Edvardsen, R.B.; Furmanek, T.; Andersson, E.; Juanchich, A.; Wargelius, A. Bmp15l, Figla, Smc1bl, and Larp6l are preferentially expressed in germ cells in Atlantic salmon (Salmo salar L.): Germ cell factors in salmon. Mol. Reprod. Dev. 2017, 84, 76–87. [Google Scholar] [CrossRef] [PubMed]
- He, F.-X.; Jiang, D.-N.; Huang, Y.-Q.; Mustapha, U.F.; Yang, W.; Cui, X.-F.; Tian, C.-X.; Chen, H.-P.; Shi, H.-J.; Deng, S.-P.; et al. Comparative transcriptome analysis of male and female gonads reveals sex-biased genes in spotted scat (Scatophagus argus). Fish Physiol. Biochem. 2019, 45, 1963–1980. [Google Scholar] [CrossRef] [PubMed]
- Palma, P.; Nocillado, J.; Superio, J.; Ayson, E.G.D.J.; Ayson, F.; Bar, I.; Elizur, A. Gonadal response of juvenile protogynous grouper (Epinephelus fuscoguttatus) to long-term recombinant follicle-stimulating hormone administration. Biol. Reprod. 2019, 100, 798–809. [Google Scholar] [CrossRef]
- Jeng, S.-R.; Wu, G.-C.; Yueh, W.-S.; Kuo, S.-F.; Dufour, S.; Chang, C.-F. Gonadal development and expression of sex-specific genes during sex differentiation in the Japanese eel. Gen. Comp. Endocrinol. 2018, 257, 74–85. [Google Scholar] [CrossRef]
- Hayman, E.S.; Fairgrieve, W.T.; Luckenbach, J.A. Molecular and morphological sex differentiation in sablefish (Anoplopoma fimbria), a marine teleost with XX/XY sex determination. Gene 2021, 764, 145093. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Hannigan, B.; Crews, D. Temperature shift alters DNA methylation and histone modification patterns in gonadal aromatase (Cyp19a1) gene in species with temperature-dependent sex determination. PLoS ONE 2016, 11, e0167362. [Google Scholar] [CrossRef]
- Pan, B.; Chao, H.; Chen, B.; Zhang, L.; Li, L.; Sun, X.; Shen, W. DNA methylation of germ-cell-specific basic helix-loop-helix (HLH) transcription factors, Sohlh2 and Figlα during gametogenesis. Mol. Hum. Reprod. 2011, 17, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Ribas, L.; Vanezis, K.; Imués, M.A.; Piferrer, F. Treatment with a DNA methyltransferase inhibitor feminizes zebrafish and induces long-term expression changes in the gonads. Epigenet. Chromatin 2017, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Yamada, K.; Fujimori, C.; Kikuchi, M.; Kawasaki, T.; Siegfried, K.R.; Sakai, N.; Tanaka, M. Germ cells in the teleost fish medaka have an inherent feminizing effect. PLoS Genet. 2018, 14, e1007259. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Kinoshita, M.; Kanamori, A. Medaka factor in the germline alpha (Figα) is indispensable for oogenesis but not for gonadal sex differentiation. Aquac. Fish. 2023; in press. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | Application |
|---|---|---|
| Figla-F1 | CAGTCTGTCAGGTTGGGTAT | Partial sequence obtaining |
| Figla-R1 | TTGGTACGCAGGCATCACA | |
| Figla-5′Rout | TACAAGCCGTCCTTAGTCCGTCTGAA | 5′-Race PCR amplification |
| Figla-5′Rin | CCGTTAAACGCATCAAAATGTCACTC | |
| Figla-3′Fout | AGACTGATGGCTTGGGCAATGACCT | 3′-Race PCR amplification |
| Figla-3′F in | CGGAGGATGGAGATATGAGCAGGC | |
| Figla-F2 | GAGTGGAAAACACACTGC | ORF qualifying |
| Figla-R2 | AGAGCAGATGGCTTCTAC | |
| Figla-BSP-F | TTTCGGTTTATTTCGTGAGTTT | DNA methylation analysis |
| Figla-BSP-R | ACCCTCTACCCACGACTACCT | |
| Figla-qF | CAACGCCAAGGAACGACTG | Quantitative real-time PCR |
| Figla-qR | CTCTCCATAAGTCATTGCCCA | |
| β-actin-qF | GCAAGCAGGAGTATGATGAG | |
| β-actin-qR | TTGGGATTGTTTCAGTCAGT | |
| EF1α-qF | GGGACACCCACAATAACATCC | |
| EF1α-qR | CCAGGCATACTTGAAGGAGC | |
| Figla-JC-F | ACCGCAAACCCAGTAAAGTC | Knockout detection |
| Figla-JC-R | TACATTCATAAAGAGTATTTCCACA | |
| Figla-ISH-F | CGGACTAAGGACGGCTTGT | In situ hybridization |
| Figla-ISH-R | AATTGGTACGCAGGCATCAC | |
| Figla-ISH-T7-zy-F | TAATACGACTCACTATAGGGCGGACTAAGGACGGCTTGT | |
| Figla-ISH-T7-zy-R | AATTGGTACGCAGGCATCAC | |
| Figla-ISH-T7-fy-F | CGGACTAAGGACGGCTTGT | |
| Figla-ISH-T7-fy-R | TAATACGACTCACTATAGGGAATTGGTACGCAGGCATCAC |
| Primer Name | Sequence (5′→3′) | Length (bp) |
|---|---|---|
| Figla-gDNA-F1 | GAGTGTAAAGGGGTTGGTAC | 1969 |
| Figla-gDNA-R1 | ATTCGCTTTTGAGATTTATGTA | |
| Figla-gDNA-F2 | GTAGAAGTACCTCAAAAACCGT | 1116 |
| Figla-gDNA-R2 | CACTTACGCTTGGCTTCTT | |
| Figla-gDNA-F3 | GTCTAAGAACAGACTATGAGCTTGA | 1841 |
| Figla-gDNA-R3 | TGTACGGCAAAGCCCAAT | |
| Figla-gDNA-F4 | AGGATGTACCATGCCATAGTAG | 1879 |
| Figla-gDNA-R4 | GATAGGGGTCCATGTAACAGTAC | |
| Figla-gDNA-F5 | AACAAAGACGGCAGAGGTGAAT | 1773 |
| Figla-gDNA-R5 | AACAGTCCATCTCCCTCCTCG |
| Primer Name | Sequence (5′→3′) | Locus |
|---|---|---|
| Figla-gRNA1 | TAATACGACTCACTATAGTGTCGACTTTACTGGGTTTGGTTTTAGAGCTAGAAATAGC | Exon 2 |
| Figla-gRNA2 | TAATACGACTCACTATAgTATGGTCAGACTGATGGCTTGTTTTAGAGCTAGAAATAGC | Exon 3 |
| Common reverse | AAAAAAAGCACCGACTCGGT | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lu, Y.; Xu, F.; Zhang, X.; Wu, Y.; Zhao, J.; Luo, Q.; Liu, H.; Chen, K.; Fei, S.; et al. Molecular Characterization, Expression Pattern, DNA Methylation and Gene Disruption of Figla in Blotched Snakehead (Channa maculata). Animals 2024, 14, 491. https://doi.org/10.3390/ani14030491
Zhang Y, Lu Y, Xu F, Zhang X, Wu Y, Zhao J, Luo Q, Liu H, Chen K, Fei S, et al. Molecular Characterization, Expression Pattern, DNA Methylation and Gene Disruption of Figla in Blotched Snakehead (Channa maculata). Animals. 2024; 14(3):491. https://doi.org/10.3390/ani14030491
Chicago/Turabian StyleZhang, Yang, Yuntao Lu, Feng Xu, Xiaotian Zhang, Yuxia Wu, Jian Zhao, Qing Luo, Haiyang Liu, Kunci Chen, Shuzhan Fei, and et al. 2024. "Molecular Characterization, Expression Pattern, DNA Methylation and Gene Disruption of Figla in Blotched Snakehead (Channa maculata)" Animals 14, no. 3: 491. https://doi.org/10.3390/ani14030491