


New Evidence for the Bronze Age Zooarchaeology in the Inland Area of the Iberian Peninsula through the Analysis of Pista de Motos (Villaverde Bajo, Madrid)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Pista de Motos and a Regional Overview

2.1. Materials
2.2. Methods: Zooarchaeology and Taphonomy
3. Results: Zooarchaeological and Taphonomic
3.1. Taxonomical Representation and Mortality Profiles
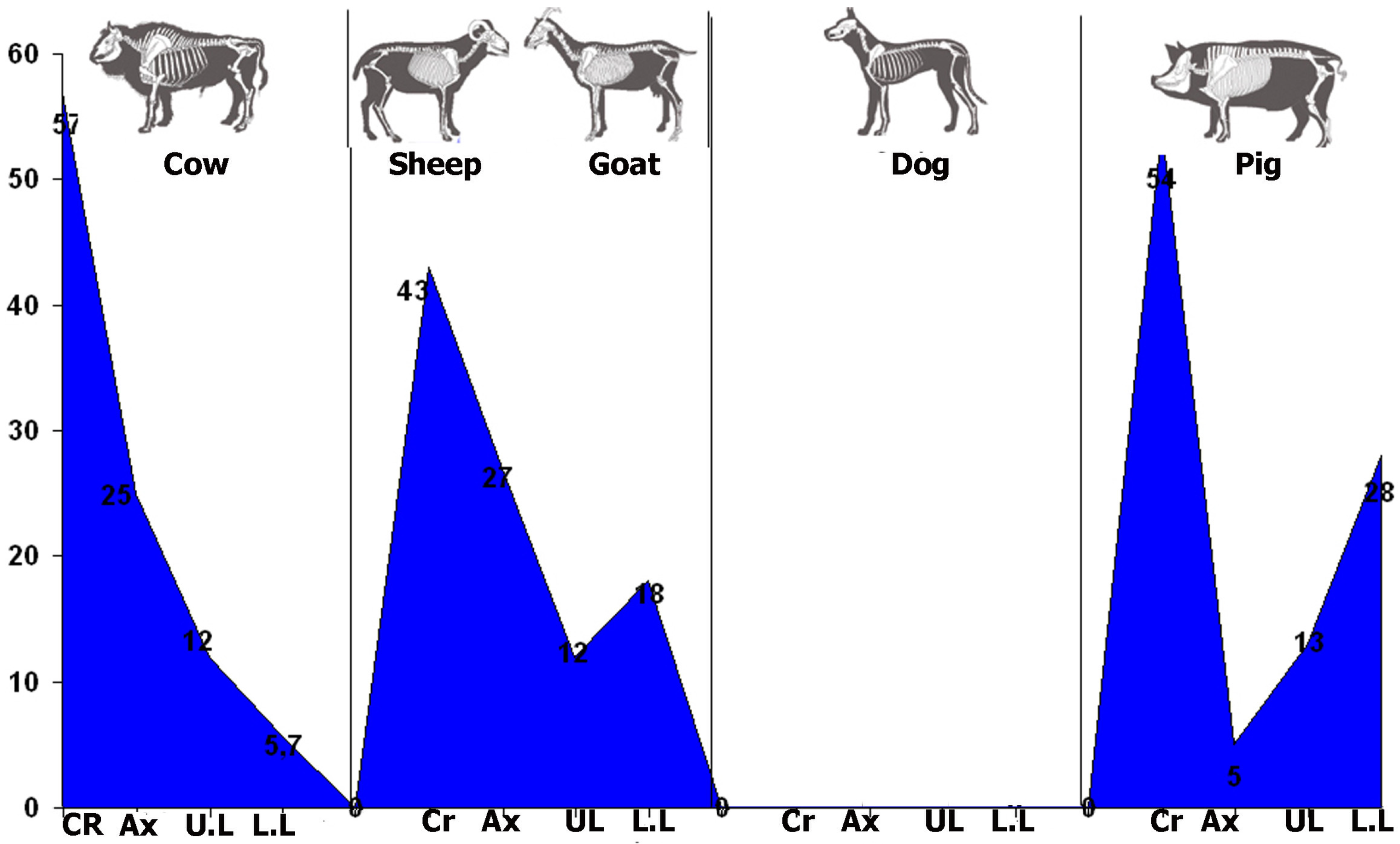
3.2. Analysis of the Skeletal Profiles
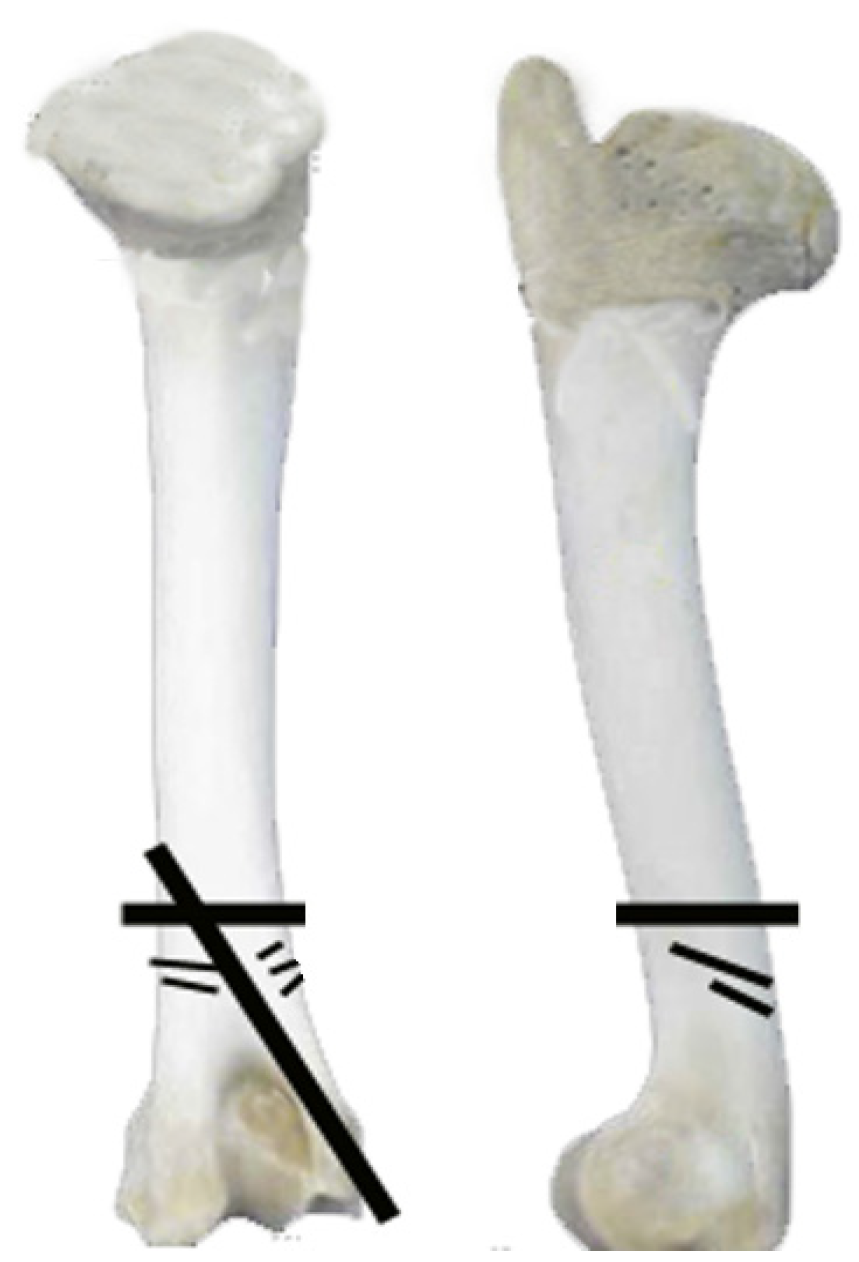
3.3. Taphonomical Analysis of Bone Surfaces
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panera, J.; Rubio-Jara, S. Bifaces y Elefantes. Zona Arqueológica 1; Museo Arqueológico Regional Madrid: Madrid, Spain, 2002; 510p. [Google Scholar]
- Altuna, J.; Mariezkurrena, K. Tipo de cabañas ganaderas durante el Neolítico del País Vasco y zonas próximas. Archeofauna 2009, 18, 137–157. [Google Scholar]
- Liesau, C.; Morales, A. Las transformaciones económicas del neolítico en la Península Ibérica: La ganadería. In El Neolítico en la Península Bérica y su Contexto Europeo; Rojo-Guerra, M., Garrido, R., García Martínez, I., Eds.; Cátedra: Madrid, Spain, 2012; pp. 107–128. [Google Scholar]
- Morales, A.; Liessau, C. Arqueozoología del Calcolítico en Madrid. Ensayo crítico de síntesis. In El Horizonte Campaniforme de la región de Madrid en el Centenario de Ciempozuelos; Blasco, C., Ed.; Departamento de Prehistoria y Arqueología, Universidad Autonoma de Madrid: Madrid, Spain, 1994; pp. 227–247. [Google Scholar]
- Díaz del Río, P. La Formación del Paisaje Agrario: Madrid en el III y II Milenios BC; Series de la Consejería de la Comunidad de Madrid; Arqueología, Paleontología y Etnografía, 9; Comunidad de Madrid: Madrid, Spain, 2001. [Google Scholar]
- Yravedra, J.; Estaca, V. Implicaciones de la Zooarqueología en la Edad del Hierro. Zona Arqueol. 2015, 17, 361–377. [Google Scholar]
- Estaca-Gómez, V. La zooarqueología Durante la Edad del Hierro en el Valle Medio del Tajo; AUDEMA: Madrid, Spain, 2017. [Google Scholar]
- Estaca-Gómez, V. Prácticas socioeconómicas de la fauna doméstica en la Edad del Hierro en el Valle Medio del Tajo. Complutum 2018, 29, 387–406. [Google Scholar] [CrossRef]
- Estaca-Gómez, V.; Linares-Matás, G.J. Husbandry practices among Iron Age communities in the centre of the Iberian Peninsula. Archaeol. Anthropol. Sci. 2019, 11, 5009–5022. [Google Scholar] [CrossRef]
- Blasco, C.; Delibes, G.; Baena, J.; Liesau, C.; Ríos, P. El poblado calcolítico de Camino de las Yeseras un escenario favorable para el estudio de la incidencia campaniforme en el Interior Peninsular. Trab. Prehist. 2007, 64, 151–163. [Google Scholar]
- Blasco, M.C.; Liesau, C.; Ríos, P. Yacimientos Calcolíticos con Campaniforme de la Región de Madrid: Nuevos Estudios; Universidad Autónoma de Madrid: Madrid, Spain, 2011. [Google Scholar]
- Liesau, C.; Blasco, C.; Ríos, P.; Vega, J.; Menduiña, R.; Blanco, J.F.; Baena, J.; Herrera, T.; Petri, A.; Gómez, J.L. Un espacio compartido por vivos y muertos: El poblado calcolítico de fosos de Camino de las Yeseras (San Fernando de Henares, Madrid). Complutum 2008, 18, 97–120. [Google Scholar]
- Liesau, C.; Daza, A.; Llorente, L.; Morales, A. More questions than answers: The singular animal deposits from Camino de Las Yeseras (Chalcolithic, Madrid, Spain). Anthropozoologica 2013, 48, 277–286. [Google Scholar] [CrossRef]
- Liesau, C. Los mamíferos de estructuras de carácter doméstico y funerario. In Yacimientos Calcolíticos con Campaniforme de la Región de Madrid. Nuevos Estudios; Blasco, C., Liesau, C., Ríos, P., Eds.; Patrimonio Arqueológico de Madrid 6 (Madrid 2011); Universidad Autónoma de Madrid: Madrid, Spain, 2011; pp. 171–198. [Google Scholar]
- Liesau, C. Depósitos con ofrendas de animales en yacimientos Cogotas I: Antecedentes y características. In Cogotas I, Una Cultura de la Edad del Bronce en la Península Ibérica; Rodríguez Marcos, J.A., Fernández Manzano, J., Eds.; Homenajes a M. Dolores Fernández-Posse; Universidad de Valladolid: Valladolid, Spain, 2012; pp. 219–257. [Google Scholar]
- Liesau, C. Fauna in Living and Funerary Contexts of the 3rd Millennium BC in Central Iberia. In Key Resources and Sociocultural Developments in the Iberian Chalcolithic; Bartelheim, M., Bueno Ramírez, P., Kunst, M., Eds.; Tübingen Publishing: Tübingen, Germany, 2017; pp. 107–128. [Google Scholar]
- Chorro, M.A. Estudio de la Fauna Calcolítica del Área Central de Camino de las Yeseras (San Fernando de Henares, Madrid). Master’s Thesis, Universidad Autónoma de Madrid, Madrid, Spain, 2013. [Google Scholar]
- López, G.; Maestre, A.; Rodríguez, A.; López, M. Viña de la Huerta. In Recuperando el Pasado. La Prehistoria Reciente en la Depresión Prados-Guatén; López, G., Morín, J., Rus, I., De Aragón, E.M., Eds.; Audema Editorial: Madrid, Spain, 2011; pp. 63–99. [Google Scholar]
- López, G.; Maestre, A.; Rodríguez, A.; López, M. Viña de la Huerta II. In Recuperando el Pasado. La Prehistoria Reciente en la Depresión Prados-Guatén; López, G., Morín, J., Rus, I., De Aragón, E.M., Eds.; Audema Editorial: Madrid, Spain, 2011; pp. 99–139. [Google Scholar]
- López, G.; Maestre, A.; Morín, J.; Rodríguez, A. Las Zanjillas. In Recuperando el Pasado. La Prehistoria Reciente en la Depresión Prados-Guatén; López, G., Morín, J., Rus, I., De Aragón, E.M., Eds.; Audema Editorial: Madrid, Spain, 2011; pp. 139–189. [Google Scholar]
- Domínguez-Alonso, R.; Virseda, L. Yacimiento 718-05-H-04. Pista de Motos. In Actas de las Cuartas Jornadas de Patimonio Arqueológico en la Comunidad de Madrid; Dirección General del Patrimonio Histórico, Comunidad de Madrid: Madrid, Spain, 2009; pp. 327–332. [Google Scholar]
- Blasco, C.; Liessau, C.; Ríos, P.; López, P.; Flores, R. Un enterramiento múltiple del yacimiento calcolítico de Humanejos (Parla, Madrid) desde una perspectiva tafonómica: Agrupando y reagrupando la familia. Cuad. Prehist. Y Arqueol. Univ. Autónoma Madr. 2014, 40, 11–29. [Google Scholar]
- Estaca-Gómez, V.; Yravedra, J. Informe Arqueozoológico del Yacimiento de Humanejos (Parla, Madrid); Manuscrito Inédito; Museo Arqueológico Regional, Comunidad de Madrid: Madrid, Spain, 2015. [Google Scholar]
- Liessau Von Lettow-Vorbeck, C.; García García, J.; Carrión Santafé, E.; Blasco Bosqued, C. El depósito ritual del fondo 76-78 de la fábrica de ladrillos (Getafe, Madrid). Cuad. Prehist. Y Arqueol. Univ. Autónoma Madr. 2015, 30, 47–56. [Google Scholar] [CrossRef]
- Yravedra, J. Arqueozoología y Tafonomía del Yacimiento Calcolítico del Barranco del Herrero (San Martín de la Vega, Madrid). In Estudios de Prehistoria y Arqueología Madrileñas 14-15-16; Estudios de Prehistoria y Arqueología Madrileñas: Madrid, Spain, 2007; pp. 427–440. [Google Scholar]
- Galindo, L.; San José, V.; Sánchez, M.; Sánchez, M.; Lorente, M. Soto del Henares. Aproximación a un poblado de recintos. In Actas de las Cuartas Jornadas de Patrimonio Arqueológico de la Comunidad de Madrid; Benet, N., Benito Lopez, J.E., Eds.; Museo Arqueológico Regional: Henares, Spain, 2009; pp. 263–271. [Google Scholar]
- Moreno-García, M.; Cantalapiedra, V. Sobre el aprovechamiento de recursos de origen animal en la región de Madrid durante el III milenio cal. AC: La fauna de los contextos calcolíticos del Sector 3 de Las Cabeceras (Pozuelo de Alarcón, Madrid). BSAA Arqueol. 2020, 85–86, 177–218. [Google Scholar] [CrossRef]
- García Somoza, P. Zooarqueología de los sectores 0 y Vía Pecuaria del yacimiento Ampliación Aguas Vivas. In El yacimiento Arqueológico de Aguas Vivas. Prehistoria Reciente en el Valle del Río Henares (Guadalajara); Cantalapiedra-Jiménez, V., Ismodes-Ezcurra, A., Eds.; La Ergástula Ediciones: Madrid, Spain, 2010; pp. 133–161. [Google Scholar]
- Yravedra, J. Zooarqueología de los sectores 1 y 2 del Yacimiento Ampliación Aguas Vivas. In El yacimiento Arqueológico de Aguas Vivas. Prehistoria Reciente en el Valle del Río Henares (Guadalajara); Cantalapiedra-Jiménez, V., Ismodes-Ezcurra, A., Eds.; La Ergastula Ediciones: Madrid, Spain, 2010; pp. 121–132. [Google Scholar]
- Estaca-Gómez, V.; Yravedra, J. Informe Arqueozoológico del yacimiento de Entreviñas I (Toledo); Museo Arqueológico Regional, Comunidad de Madrid: Madrid, Spain, 2011; Manuscript unpublished. [Google Scholar]
- Estaca, V.; López, G.; Morín, J.; Yravedra, J. Estudio Zooarqueológico y Tafonómico del Yacimiento Calcolítico Las Zanjillas (Torrejón de Velasco, Madrid); Museo Arqueológico Regional, Comunidad de Madrid: Madrid, Spain, 2023; Munibe Submited. [Google Scholar]
- Estaca-Gómez, V.; de la Torre, A.; Señorán, J.M.; Martínez AB Major, M.; Yravedra, J. Aprovechamiento de recursos animales en el yacimiento Calcolítico precampaniforme del área Aldovea (Torrejón de Ardoz, Madrid, España). Complutum 2023, 34, 1. [Google Scholar] [CrossRef]
- Priego, C.M. El yacimiento de Angosto de los Mancebos. Nueva contribución al conocimiento de la edad del bronce madrileño. Estud. Prehist. Y Arqueol. Madrileños 1994, 9, 91–97. [Google Scholar]
- Martínez Navarrete, M.I. El yacimiento de La Esgaravita (Alcalá de Henares, Madrid) y la cuestión de los llamados fondos de cabaña del valle del Manzanares. Trab. Prehist. 1979, 36, 83–118. [Google Scholar]
- Molero Gutiérrez, G.; Brea López, P.; Bustos Pretel, V. Estudio faunístico de la cueva de Juan Barbero (Tielmes, Madrid). Trab. Prehist. 1984, 41, 105–112. [Google Scholar]
- Liesau, C. Análisis faunístico de los yacimientos de “Huerta de los Cabreros”, “Cantera de la Flamenca” y “Puente Largo del Jarama” (Aranjuez, Madrid). In El Poblamiento desde el Neolítico Final a la Primera Edad del Hierro en la Cuenca Media del río Tajo; Muñoz, K., Ed.; UCM: Madrid, Spain, 1998; pp. 1418–1444. [Google Scholar]
- Cerdeño, M.E.; Herráez, E. Estudio de la fauna del Yacimiento del Espinillo (Villaverde, Madrid). En Baquedano; et al. (Ed.) El Espinillo: Un yacimiento calcolítico y de la edad del bronce en las terrazas del Manzanares. Arqueol. Paleontol. Y Etnogr. 2000, 8, 141–149. [Google Scholar]
- Priego, C.M.; Quero, S. El Ventorro, un poblado prehistórico de los albores de la metalurgia. Estud. Prehist. Arqueol. Madrileñas 1992, 8, 1–131. [Google Scholar]
- Morales, A. Estudio faunístico de las osamentas de animales del yacimiento prehistórico del Negralejo. En Un nuevo yacimiento del Bronce madrileño, El Negralejo (Rivas, Vaciamadrid). Not. Arqueol. Hispánico 1983, 17, 166–177. [Google Scholar]
- Morales, A.; Villegas, C. La fauna de mamíferos del yacimiento de “El Ventorro”. Pecado-tesis osteológica de la campaña de 1981. Estud. Prehist. Arqueol. Madrileñas 1994, 9, 35–56. [Google Scholar]
- Díaz del Río, P.; Consuegra, S.; Peña Chocarro, L.; Márquez, B.; Sanpedro, C.; Moreno, R.; Albertini, D.; Pino, B. Paisajes agrarios prehistóricos en la Meseta Peninsular: El caso de las Matillas (Alcalá de Henares, Madrid). Trab. Prehist. 1997, 54, 93–111. [Google Scholar] [CrossRef]
- González, J.M. Interpretación arqueológica de un “Campo de Hoyos” en Forfoleda (Salamanca). Zephyrus 2009, 46, 309–313. [Google Scholar]
- Alameda, M.C.; Carmona, E.; Pascual SMartínez, G.; Díez, C. El “campo de hoyos” calcolítico de Fuente Celada (Burgos): Datos preliminares y perspectivas. Complutum 2011, 22, 47–69. [Google Scholar]
- Bronk Ramsey, C. Dealing with Outliers and Offsets in Radiocarbon Dating. Radiocarbon 2009, 51, 1023–1045. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. Bayesian Analysis of Radiocarbon Dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef]
- Reimer, P.; Austin, W.; Bard, E.; Bayliss, A.; Blackwell, P.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Schmid, E. Atlas of Animal Bones for Prehistorians, Archaeologist and Quaternary Geologist; Elsevier Publishing Company: Amsterdam, The Netherlands; London, UK; New York, NY, USA, 1972. [Google Scholar]
- Hilson, S. Mammal Bones and Teeth: An Introductory Guide to Methods of Identification; London Institute of Archaeology: London, UK, 1992. [Google Scholar]
- Boesseneck, J. Osteological diferences between sheep (Ovis aries Linné) and goats (Capra hircus linné). In Science in Archaeology; Brothwell, D., Higgs, D., Eds.; Thames & Hudson: Galway, Ireland, 1969; pp. 331–358. [Google Scholar]
- Payne, S. Morphological distinction between the mandibular teeth of young sheep, Ovis and goats, Capra. J. Archaeol. Sci. 1985, 12, 139–147. [Google Scholar] [CrossRef]
- Prummel, W.; Frisch, H.J. A guide for the distinction of species, sex and body size in bones of sheep and goat. J. Archaeol. Sci. 1986, 13, 567–577. [Google Scholar] [CrossRef]
- Fernández, H. Osteologie Comparee des Petties Ruminants Eurasiatiques Sauvages et Domestiques (Genres Rupicapra, Ovis, Capra et Capreolus): Diagnose Differentialle du Squelette Apendiculaire; Universite de Geneva, Facultat de Ciencies: Geneva, Switzerland, 2001. [Google Scholar]
- Payne, S.; Bull, G. Components of variation in measurements of pig bones and teeth, and the use of measurements to distinguish wild from domestic pig remains. Archaeozoologia 1988, 2, 27–66. [Google Scholar]
- Yravedra, J. Tafonomía Aplicada a Zooarqueología; Aula Abierta; UNED: Madrid, Spain, 2006. [Google Scholar]
- Brain, C.K. The contribution of Namib desert Hottentot to understanding of Australopithecus bone accumulations. Sci. Pap. Namib. Deser Res. Stn. 1969, 32, 1–11. [Google Scholar]
- Pérez Ripoll, M. Estudio de la secuencia del desgaste de los molares de Capra pyrenaica de los yacimientos prehistóricos. Arch. Prehist. Levantina 1988, 18, 83–128. [Google Scholar]
- Payne, S. Kill-off pattern in sheep and goats: The mandibles of Açvan Kale. Anatol. Stud. 1973, 23, 281–303. [Google Scholar] [CrossRef]
- Payne, S. Reference codes for wear states in the mandibular cheek teeth of sheep and goats. J. Archaeol. Sci. 1987, 14, 609–614. [Google Scholar] [CrossRef]
- Ducos, P. L’Origine des animaux domestiques en Palestine. In Publications de l’Institut de Préhistoire de l’Université de Bordeau /Mémoire; Delmas: Bordeaux, France, 1968. [Google Scholar]
- Grant, A. The use of tooth wear as a guide to the age of domestic ungulates. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; BAR International Series 109; BAR Publishing: Oxford, UK, 1982; pp. 91–108. [Google Scholar]
- O’Connor, T.P. Husbandry Decisions: Age at Death. In The Analysis of Urban Animal Bones Assemblages: A Hand Book for Archaeologists; Council for British Archaeology: York, UK, 2003; pp. 157–165. [Google Scholar]
- Barone, R. Anatomie Comparée des Mammifères Domestiques, 2nd ed.; Tome I y II: Revue et Mise à a Jour; Vigot Frèges: Paris, France, 1999. [Google Scholar]
- Blumenschine, R.J. Carcass consumption sequences and the archaeological distinction of scavenging and hunting. J. Hum. Evol. 1986, 15, 639–659. [Google Scholar] [CrossRef]
- Blumenschine, R.J. 1995. Percussion marks, tooth marks, and experimental determination of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 1995, 29, 21–51. [Google Scholar] [CrossRef]
- Behrensmeyer, A.K. Taphonomic and ecological information from bone weathering. Paleobiology 1978, 4, 150–162. [Google Scholar] [CrossRef]
- Binford, L.R. Bones: Ancient Men and Modern Myths; Academic Press Inc.: New York, NY, USA, 1981. [Google Scholar]
- Blumenschine, R.J.; Salvaggio, M.M. Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 1988, 333, 763–765. [Google Scholar] [CrossRef]
- Fisher, D.C. Bone surface modifications in zooarchaeology. J. Archaeol. Method Theory 1995, 2, 7–65. [Google Scholar] [CrossRef]
- Fernández Jalvo, Y.; Andrews, P. Atlas of Taphonomy Identification; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Yravedra, J.; Uzquiano, P. Burnt bone assemblages from El Esquilleu cave (Cantabria, Northern Spain): Deliberate use for fuel or systematic disposal of organic waste? Quat. Sci. Rev. 2012, 68, 175–190. [Google Scholar] [CrossRef]
- Villa, P.; Mahieu, E. Breakage patterns of human long bones. J. Hum. Evol. 1991, 20, 1–22. [Google Scholar] [CrossRef]
- Schwartz, M. A History of Dog’s in the Early Americas; Yale University Press: New Haven, UK, 1997. [Google Scholar]
- Howald, G.; Donlan, C.J.; Galvan, J.P.; Russell, J.C.; Parkes, J.; Samaniego, A.; Wang, Y.W.; Veitch, D.; Genovesi, P.; Pascal, M.; et al. Invasive rodent eradicationon islands. Conserv. Biol. 2007, 21, 1258–1268. [Google Scholar] [CrossRef]
- Butler, J.R.A.; Toit, J.T.; Bingham, J. Free-ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: Threats of competition and disease to large wild carnivores. Biol. Conserv. 2004, 115, 369–378. [Google Scholar] [CrossRef]
- Sillero-Zubiri, C.; Gottelli, D. Diet and feeding behavior of Ethiopian wolves (Canis simensis). J. Ofmammal. 1995, 76, 531–541. [Google Scholar] [CrossRef]
- Russell, J.C.; Beaven, B.M.; MacKay, J.W.B.; Towns, D.R.; Clout, M.N. Testing island biosecurity systems for invasive rats. Wildl. Res. 2008, 35, 215–221. [Google Scholar] [CrossRef]
- Escosteguy, P.; Salerno, V. La caza del coipo. Su importancia económica y social desde momentos prehispánicos hasta la actualidad. An. Arqueol. Y Etnol. 2008, 63–64, 277–303. [Google Scholar]
- Gsell, A.; Innes, J.; de Monchy, P.; Brunton, D. The success of using trained dogs to locate sparse rodents in pest-free sanctuaries. Wildl. Res. 2010, 37, 39–46. [Google Scholar] [CrossRef]
- Shapira, I.; Buchanan, F.; Brunton, D. Detection of caged and free-ranging Norway rats Rattus norvegicus by a rodent sniffing dog on Browns Island, Auckland, New Zealand. Conserv. Evid. 2011, 8, 38–42. [Google Scholar]
- Deleidi, F. Parson jack russell terrier. In El Gran Libro de los Terriers; Editorial de Vecchi: Barcelona, Spain, 1991; pp. 327–331. [Google Scholar]
- Cárcel Rubio, M.J. El perro ratonero valenciano, Gos rater valencià. In Boletín Informativo el Perro en España; Real Sociedad Canina de España. Número 24 (II/2008); Real Sociedad Canina de España: Valencia, Spain, 2009; pp. 59–64. [Google Scholar]
- Jaén Téllez, J.A. 2012. El Perro Ratonero-Bodeguero Andaluz. Diferencias con Otros Terriers y Ratoneros Españoles. REDVET Rev. electrón. vet. Volumen 13 Nº 11. Available online: https://www.researchgate.net/publication/368190001_El_perro_ratonero-bodeguero_andaluz_Diferencias_con_otros_terriers_y_ratoneros_espanoles (accessed on 15 December 2023).
- Yravedra, J. Estudio arqueozoológico del Caserío de Perales. In Nuevos Datos para el Estudio de la Edad del Bronce en la Comunidad de Madrid; López, G., Morín, J., Eds.; Los yacimientos de Perales II y la Peineta. MArq Audema. Serie Prehistoria 5; Audema Editorial: Madrid, Spain, 2007. [Google Scholar]
- Yravedra Sainz de los Terreros, J. Anejo V–III. Estudio Arqueozoológico. Los Yacimientos de Merinas y Velilla (Mocejón, Toledo). Un Modelo de Estudio para la Edad del Bronce en la Provincia de Toledo; Morín de Pablos, J., Ed.; MArq AUDEMA Serie Época Romana/Antigüedad Tardía 6; Audema Editorial: Madrid, Spain, 2007; pp. 166–174. ISBN 978-84-611-7542-0. [Google Scholar]
- Soto, E. Análisis de los restos faunísticos del yacimiento de «fondos de cabaña» de Getafe. Estud. Prehist. Arqueol. Madrileños 1983, 2, 279–284. [Google Scholar]
- Molero, G. Anexo I estudio de los restos óseos hallados en la zona A/norte de la Loma del Lomo (Cogolludo, Guadalajara). Camapañas de 1985–1994)- La Loma del Lomo III, Cogolludo Guadalajara. In Cogolludo Guadalajara; Valiente, J., del Lomo, L.L., III, Eds.; Patrimonio histórico Arqueología Castilla la Mancha: Guadalajar, Spain, 2001. [Google Scholar]
- Aguilar, A.L.; Maicas, R.; Morales, A.; Moreno, R. Análisis faunístico del yacimiento arqueológico de Perales del Río (Madrid). Arqueol. Paleontol. Etnogr. 1991, 1, 149–181. [Google Scholar]
- Díaz del Río, P. Campesinado y gestión pluriactiva del ecosistema: Un marco teórico para el análisis del III y el II milenios a.C. en la Meseta peninsular. Trab. Prehist. 1995, 52, 99–109. [Google Scholar] [CrossRef]
- Yravedra Sainz de los Terreros, J. Estudio Arqueozoológico. In Recuperando el Pasado. La Prehistoria Reciente en la Depresión Prados-Guatén (Comunidad de Madrid); López, G., Rus, I., Aragón, E., Eds.; Comunidad de Madrid: Madrid, Spain, 2010; pp. 201–220. ISBN 978-84-612-9360-5. [Google Scholar]
- Liesau, C.; Blasco, C. Depósitos de fauna en yacimientos del Bronce Medio en la Cuenca del Tajo. In Actas do IV Congreso de Arqueología Peninsular (Faro, 14–19 de Sept. de 2004); Universidade do Algarve: Faro, Portugal, 2006; pp. 81–92. [Google Scholar]
- Quero, S. El poblado del Bronce Medio de Tejar del Sastre (Madrid). Estud. Prehist. Arqueol. Madrileñas 1982, 1, 183–248. [Google Scholar]
- Macarro, J.A. La Alcalá prehistórica. El poblado de la Edad del Bronce de “La Dehesa”. In Alcalá de Henares, Ayuntamiento de Alcalá de Henares; Comunidad de Madrid: Madrid, Spain, 2000; 330p. [Google Scholar]
- Blasco, M.C.; Barrio, J. Excavaciones de dos nuevos asentamientos prehistóricos de Getafe (Madrid). Not. Arqueol. Hispánico 1986, 27, 75142. [Google Scholar]
- Márquez-Romero, J.E. Sobre los depósitos estructurados de animales en yacimientos de fosos del Sur de la Península Ibérica. In Animais na Pré-Hisória e Arqueología da Península Ibérica; Actas do IV Congresso de Arqueología Peninsular: Braga, Portugal, 2006; pp. 15–25. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phases/Samples | Uncalibrated Dates BP [21] | Modelled Date (95.4%) cal BP |
|---|---|---|
| End Middle Bronze Age | 3384–2933 | |
| SU1660 | 3090 ± 24 | 3381–3241 |
| SU2581 | 3161 ± 27 | 3450–3343 |
| SU962 | 3269 ± 21 | 3558–3402 |
| Start Middle Bronze Age | 3901–3407 | |
| End Bell Beaker | 4219–3691 | |
| SU2417 | 3764 ± 25 | 4232–4001 |
| SU1251 | 3768 ± 31 | 4234–4001 |
| Start Bell Beaker | 4546–4011 |
| NISP | Neolithic | Bell Beaker Bronze Age | % | Middle Bronze Age | % | Iron Age | Visigoths | Islamic | Prehistoric | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| Bos taurus | 3 | 263 | 27.4 | 1293 | 18.9 | 2 | 15 | 204 | 1779 | |
| Ovis aries | 17 | 1.8 | 53 | 0.8 | 7 | 3 | 19 | 99 | ||
| Capra hircus | 1 | 8 | 0.8 | 333 | 4.9 | 1 | 6 | 523 | ||
| Ovis/Capra | 6 | 103 | 10.7 | 969 | 14.2 | 5 | 15 | 1 | 107 | 1206 |
| All Ovis and Capra | 7 | 128 | 13.3 | 1529 | 22.4 | 12 | 19 | 1 | 132 | 1828 |
| Canis famil. | 1 | 0.1 | 476 | 7.0 | 64 | 541 | ||||
| Sus domest. | 39 | 4.1 | 604 | 8.8 | 12 | 21 | 654 | |||
| Sus scrofa | 2 | 0.2 | 9 | 0.1 | 11 | |||||
| Equus ferus | 21 | 2.2 | 13 | 0.2 | 6 | 40 | ||||
| Capra pyrenaica | 174 | 2.5 | ||||||||
| Cervus elaphus | 1 | 0.1 | 9 | 0.1 | 10 | |||||
| Oryct. cuniculus | 32 | 3.3 | 41 | 0.6 | 1 | 1 | 8 | 83 | ||
| Lepus sp. | 0.0 | 5 | 0.1 | 1 | 6 | |||||
| Corvus sp. | 0.0 | 45 | 0.7 | 45 | ||||||
| Tyto alba | 0.0 | 28 | 0.4 | 28 | ||||||
| Alectoris rufa | 4 | 0.4 | 4 | |||||||
| Birds indet | 1 | 0.0 | 1 | |||||||
| Bufo bufo | 18 | 18 | ||||||||
| Large A. Size | 8 | 302 | 31.5 | 1335 | 19.5 | 6 | 5 | 1 | 169 | 1824 |
| Small A.Size | 18 | 147 | 15.3 | 991 | 14.5 | 10 | 32 | 4 | 173 | 1372 |
| indet. | 19 | 2.0 | 469 | 6.9 | 5 | 30 | 522 | |||
| Total | 36 | 959 | 6849 | 49 | 89 | 6 | 809 | 8766 |
| MNI | Neolithic | Bell Beaker Bronze Age | Middle Bronze Age | Iron Age | Visigoths | Islamic | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| S-A-J-I | S-A-J-I | MNI | % | S-A-J-I | MNI | % | S-A-J-I | S-A-J-I | S-A-J-I | |
| Bos taurus | 0/1/0/0 | 0/3/2/1 | 6 | 16.7 | 2/11/2/1 | 16 | 15.7 | 0/1/0/0 | 1/1/0/0 | |
| Ovis aries | 1/2/0/0 | 3 | 8.3 | 1/4/0/0 | 5 | 4.9 | 0/1/0/0 | 0/1/0/0 | ||
| Capra hircus | 0/1/0/0 | 1-1-0-0 | 2 | 5.6 | 2/6/1/3 | 12 | 11.8 | 0/1/0/0 | ||
| Ovis/Capra | 0/1/0/0 | 1-2-1-1 | 5 | 13.9 | 1/12/1/1 | 15 | 14.7 | 0/1/0/0 | 1/1/0/0 | 0/1/0/0 |
| All Ovis and Capra | 1/3/1/1 | 6 | 16.7 | 3/12/1/3 | 19 | 18.6 | 0/1/0/0 | 1/1/0/0 | 0/1/0/0 | |
| Canis familiaris | 0/1/0/0 | 1 | 2.8 | 0/10/0/0 | 10 | 9.8 | ||||
| Sus domesticus | 0/2/1/2 | 5 | 13.9 | 1/4/5/3 | 13 | 12.7 | 0/1/0/0 | |||
| Sus scrofa | 0/1/0/0 | 1 | 2.8 | 0/2/0/0 | 2 | 2.0 | ||||
| Equus ferus | 0/2/0/0 | 2 | 5.6 | 0/1/0/0 | 1 | 1.0 | ||||
| Cervus elaphus | 0/1/0/0 | 1 | 1.0 | |||||||
| Capra pyrenaica | 0/0/1/0 | 1 | 1.0 | |||||||
| Oryct. cuniculus | 0/4/0/0 | 4 | 11.1 | 0/3/0/1 | 4 | 3.9 | 0/1/0/0 | 0/1/0/0 | ||
| Lepus sp. | 0/1/0/0 | 1 | 1.0 | |||||||
| Corvus sp. | 0/1/0/0 | 1 | 1.0 | |||||||
| Tyto alba | 0/1/0/0 | 1 | 1.0 | |||||||
| Alectoris rufa | 0/1/0/0 | 1 | 2.8 | |||||||
| SU with Animals as a Deposit. Complete (C) or Partial (P) Named “Deposit” | SU with Bones Sample > 100 NISP | SU with Fauna and Human Inhumations |
|---|---|---|
| 1018 (Dog. MNI = 1 ad) (C) Pre 1666 (Raven, MNI = 1 ad) (C) MBA 1361 (Cow, MNI = 3ad) (1C-1P-1P) MBA 1361 (Barn Owl, MNI = 1ad) (C) MBA 1661 (Dog, MNI = 1ad) (P) MBA 1662 (Pig, MNI = 1 Inf) (C) MBA 1663 (Dog, MNI = 1ad) (C) MBA 1664 (Goat, MNI = 1 Inf) (C) MBA 2692 (Goat, MNI = 1 ad) (C) MBA 2693 (Goat, MNI = 1 Inf) (P) MBA 3171 (Cow, MNI = 2 juv) (C-P) MBA 857 (iber.Ibex–1juv) (C)—and goat –ad-(P) MBA 573 (Cow, MNI= 2 juv and ad) (C-P) BSB 981 (Cow, MNI = 1ad) (P) MBA 2371 (Pig MNI = 1 inf (P) MBA | 1251 Ovis/Capra 1331 Pig and Ovis/Capra 1336 Ovis/Capra 120 Pig 2201 Ovis/Capra 2371 Ovis/Capra) 2391 Cow 2417 Ovis/Capra 3001 Pig, Cow and Ovis/Capra 3021 Cow and Ovis/Capra 3036 Ovis/Capra 481 Cow 511 Cow 856 Cow 571 Cow 573 Cow | 1401 MBA 2227 Visig. 2565 Islam. 963 y 962 MBA |
| MNI | Bos | Equus | Ovis | Capra | Ovis/Capra | Canis | Sus d. | Sus sc | Oryctol. | Lepus |
|---|---|---|---|---|---|---|---|---|---|---|
| Neolithic | 2 | 1 | 2 | |||||||
| Bell Beaker Bronze Age | 17 | 5 | 5 | 5 | 9 | 2 | 6 | 1 | 5 | |
| Middle Bronze Age | 144 | 6 | 22 | 51 | 126 | 40 | 61 | 2 | 25 | |
| Iron Age II | 2 | 2 | 2 | 1 | 1 | |||||
| Visigoths | 3 | 1 | 1 | 3 | 1 | 1 | ||||
| Islamic Age | 1 | |||||||||
| Prehistoric | 27 | 3 | 9 | 1 | 20 | 1 | 5 | 3 | 1 |
| Bos taurus | % | Ovis aries | Capra hircus | Ovis/Capra | Ovis and Capra. Total | % | Equus | Sus | % | |
|---|---|---|---|---|---|---|---|---|---|---|
| Horn | 41 | 15.6 | 0.0 | 0.0 | ||||||
| Craneal | 34 | 12.9 | 5 | 5 | 3.9 | 0.0 | ||||
| Maxillar | 1 | 0.4 | 0.0 | 4 | 10.3 | |||||
| Mandible | 13 | 4.9 | 2 | 1 | 7 | 10 | 7.8 | 2 | 5.1 | |
| Teeth | 62 | 23.6 | 13 | 9 | 18 | 40 | 31.3 | 15 | 38.5 | |
| Vertebrae | 33 | 12.5 | 8 | 8 | 6.3 | 3 | 0.0 | |||
| Rib | 21 | 8.0 | 20 | 20 | 15.6 | 1 | 0.0 | |||
| Scapule | 5 | 1.9 | 1 | 6 | 7 | 5.5 | 1 | 0.0 | ||
| Humerus | 6 | 2.3 | 2 | 2 | 1.6 | 2 | 0.0 | |||
| Radio | 6 | 2.3 | 6 | 6 | 4.7 | 1 | 0.0 | |||
| Ulna | 6 | 2.3 | 0.0 | 1 | 2 | 5.1 | ||||
| Metacarpal | 4 | 1.5 | 3 | 3 | 2.3 | 0.0 | ||||
| Carpal | 0.0 | 1 | 1 | 0.8 | 0.0 | |||||
| Pelvis | 7 | 2.7 | 0.0 | 2 | 2 | 5.1 | ||||
| Femur | 6 | 2.3 | 1 | 1 | 0.8 | 2 | 3 | 7.7 | ||
| Tibiae | 7 | 2.7 | 6 | 6 | 4.7 | 3 | 0.0 | |||
| Metatarsal | 3 | 1.1 | 1 | 4 | 5 | 3.9 | 1 | 0.0 | ||
| Metapodial | 3 | 1.1 | 1 | 1 | 0.8 | 6 | 15.4 | |||
| Talus | 2 | 0.8 | 1 | 8 | 9 | 7.0 | 0.0 | |||
| Calcaneus | 0.0 | 1 | 1 | 2 | 1.6 | 1 | 0.0 | |||
| Phalange | 3 | 1.1 | 2 | 2 | 1.6 | 3 | 5 | 12.8 | ||
| Total | 263 | 100 | 17 | 12 | 99 | 128 | 100.2 | 21 | 39 | 100 |
| Sus scro | Cervus | Lepus | Bos | % | Ovis | Capra | Ov/Cap | All Ovis and Capra. | % | Equus | Canis | Sus | % | Oryc. | Indet Large Size | Indet Lower Size | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Horn | 40 | 3.1 | 8 | 8 | 0.5 | ||||||||||||
| Cranial | 222 | 17.2 | 1 | 67 | 68 | 4.4 | 6 | 60 | 10.4 | 59 | 11 | ||||||
| Maxillar | 1 | 25 | 1.9 | 8 | 9 | 17 | 1.1 | 16 | 12 | 2.1 | |||||||
| Mandíble | 52 | 4.0 | 3 | 15 | 34 | 52 | 3.4 | 1 | 21 | 19 | 3.3 | 4 | 13 | 2 | |||
| Teeth | 5 | 227 | 17.6 | 23 | 140 | 98 | 261 | 17.0 | 3 | 211 | 180 | 31.3 | 5 | ||||
| Vertebrae | 125 | 9.7 | 1 | 182 | 58 | 241 | 15.7 | 49 | 64 | 11.1 | 1 | 21 | 12 | ||||
| Rib | 189 | 14.6 | 0 | 103 | 280 | 383 | 25.0 | 3 | 51 | 85 | 14.8 | 45 | 7 | ||||
| Scapule | 1 | 18 | 1.4 | 2 | 8 | 21 | 31 | 2.0 | 2 | 4 | 0.7 | 1 | 3 | 1 | |||
| Humerus | 37 | 2.9 | 4 | 12 | 35 | 51 | 3.4 | 4 | 4 | 10 | 1.7 | 4 | 22 | 5 | |||
| Radio | 1 | 1 | 2 | 55 | 4.3 | 6 | 26 | 13 | 45 | 2.9 | 8 | 10 | 1.7 | 6 | |||
| Ulna | 1 | 22 | 1.7 | 10 | 10 | 20 | 1.3 | 8 | 9 | 1.6 | 2 | ||||||
| Metacarpal | 1 | 22 | 1.7 | 5 | 5 | 3 | 13 | 0.8 | 10 | 8 | 1.4 | ||||||
| Carpal | 49 | 3.8 | 30 | 30 | 2.0 | 20 | 10 | 1.7 | 2 | ||||||||
| Pelvis | 1 | 8 | 0.6 | 0 | 9 | 17 | 26 | 1.7 | 4 | 7 | 1.2 | 5 | 5 | ||||
| Femur | 2 | 16 | 1.2 | 1 | 7 | 14 | 22 | 1.4 | 4 | 10 | 1.7 | 2 | 1 | 1 | |||
| Tibia | 1 | 1 | 2 | 43 | 3.3 | 2 | 6 | 47 | 55 | 3.6 | 1 | 8 | 7 | 1.2 | 8 | 7 | 7 |
| Fibula | 0.0 | 0.0 | 2 | 0 | 0.0 | ||||||||||||
| Patella | 3 | 0.2 | 1 | 1 | 0.1 | 4 | 2 | 0.3 | |||||||||
| Metatarsal | 2 | 20 | 1.5 | 3 | 8 | 25 | 36 | 2.3 | 10 | 0.0 | |||||||
| Metapodial | 1 | 9 | 0.7 | 5 | 5 | 0.3 | 1 | 5 | 14 | 2.4 | 7 | 2 | 4 | ||||
| Talus | 2 | 15 | 1.2 | 2 | 8 | 10 | 0.7 | 4 | 0.0 | ||||||||
| Calcaneus | 1 | 13 | 1.0 | 1 | 6 | 2 | 15 | 1.0 | 5 | 2 | 0.3 | 1 | |||||
| Tarsal | 1 | 0.1 | 18 | 18 | 1.2 | 12 | 6 | 1.0 | |||||||||
| Sesamoid | 10 | 0.8 | 1 | 1 | 0.1 | 0.0 | |||||||||||
| Phalange | 71 | 5.5 | 72 | 7 | 79 | 5.2 | 72 | 57 | 9.9 | ||||||||
| Indet Shafts | 41 | 41 | 2.7 | 1140 | 838 | ||||||||||||
| Total | 11 | 10 | 5 | 1292 | 100 | 53 | 694 | 788 | 1532 | 13 | 536 | 576 | 41 | 1325 | 988 |
| Anatomical Section | Bos | % | Equus | Capra | Ov/Ca | Ovis | Ovis/Capra | % | Canis | Sus | Large Size | Small Size |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cranial | 1 | 1.3 | 1 | 6 | 2 | |||||||
| Axial | 21 | 26.6 | 1 | 2 | 11 | 1 | 14 | 19.7 | 2 | 4 | 10 | 1 |
| Ep-Dist | 6 | 7.6 | 1 | 1 | 1 | 2 | 2.8 | 2 | 2 | |||
| Ep-Prox | 13 | 16.5 | 3 | 2 | 5 | 7.0 | ||||||
| Epiphysis | 19 | 24.1 | 1 | 1 | 4 | 2 | 7 | 9.9 | 2 | 2 | ||
| Compact Bones | 7 | 8.9 | 1 | |||||||||
| Shafts | 31 | 39.2 | 1 | 3 | 35 | 2 | 40 | 56.3 | 4 | 8 | 26 | 42 |
| Total | 79 | 3 | 7 | 56 | 5 | 71 | 6 | 15 | 40 | 43 |
| NISP | Cut Mark | Tooth Mark | ||
|---|---|---|---|---|
| Bell Beaker Bronze Age | Middle Bronze Age | Bell Beaker Bronze Age | Middle Bronze Age | |
| Bos taurus | 10 | 108 | 7 | 70 |
| Equus ferus | 0 | 0 | 2 | 1 |
| Capra hircus | 0 | 9 | 0 | 7 |
| Ovis aries | 0 | 6 | 0 | 4 |
| Ovis/capra | 8 | 53 | 5 | 45 |
| Sus domestic | 0 | 16 | 12 | |
| Oryct. cuniculus | 1 | 1 | 0 | 2 |
| Canis familiaris | 4 | 1 | 5 | |
| Cervus elaphus | 1 | |||
| Sus scrofa | 1 | 1 | ||
| Indet small animal size | 5 | 27 | 1 | 36 |
| Indet large animal size | 9 | 51 | 5 | 28 |
| Total | 33 | 276 | 22 | 214 |
| Axial | Cranial | Shafts | Ep-Dist | Epiph. | Ep-Prox | Compact Bones | Total | |
|---|---|---|---|---|---|---|---|---|
| Equus | 0 | 0 | 1 | 0 | 0 | 0 | 1 | |
| Bos | 45 | 10 | 31 | 6 | 18 | 31 | 141 | |
| Oryctolagus | 0 | 0 | 1 | 1 | 0 | 0 | 2 | |
| Ovis/Capra | 39 | 4 | 26 | 2 | 0 | 1 | 72 | |
| Ovis | 2 | 0 | 2 | 1 | 0 | 1 | 6 | |
| Capra | 3 | 1 | 4 | 0 | 1 | 0 | 9 | |
| Canis | 2 | 2 | 0 | 0 | 0 | 4 | ||
| Sus | 3 | 3 | 9 | 2 | 1 | 0 | 18 | |
| Cervus | 0 | 0 | 2 | 0 | 1 | 3 | ||
| Lepus | 0 | 0 | 1 | 1 | ||||
| Large Siez | 18 | 5 | 41 | 0 | 1 | 0 | 0 | 65 |
| Small Size | 8 | 0 | 32 | 0 | 40 | |||
| Indet. | 2 | 0 | 1 | 0 | 0 | 3 | ||
| Total | 122 | 25 | 148 | 15 | 1 | 20 | 34 | 365 |
| Functionality | Disarti-culation | Skinning | Filleting and Disarticulation | Disarti-culation | Disarti-culation | Disarti-culation | Skining and Disarticulation |
| Matillas | Espinillo Chalcol.Bronze | Loma del Lomo | Espinillo Bronze | Angosta Mancebos | Barr. Del Herrero | Fondos Cabaña Getafe | Fábrica de Ladrillos | Caserío Perales | Perales del Río | Velilla I | Arenero Soto | Merinas | Viña de la Huerta I | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| References | [41] | [37] | [83] | [37] | [33] | [25] | [85] | [24] | [86] | [87] | [84] | [88] | [84] | [89] |
| NISP | NISP | NISP | NISP | NISP | NISP | NISP | NISP | NISP | NR | NISP | NISP | NISP | NISP | |
| Equus ferus | 8 | 8 | 3 | 6 | 7 | 11 | 2 | 6 | ||||||
| Bos taurus | 78 | 70 | 190 | 20 | 5 | 118 | 697 | 23 | 30 | 67 | 118 | 7 | 79 | |
| Capra hircus | 2 | 5 | 9 | 1 | 12 | 3 | ||||||||
| Ovis aries | 3 | 5 | 12 | 9 | 3 | 6 | 27 | |||||||
| Ovis/Capra | 91 | 83 | 296 | 25 | 21 | 10 | 258 | 256 | 10 | 54 | 40 | 258 | 10 | 42 |
| Canis familiaris | 6 | 144 | 44 | 1 | 22 | 12 | 6 | 7 | 9 | 22 | 3 | |||
| Sus domesticus | 18 | 27 | 7 | 2 | 16 | 3 | 47 | 8 | 3 | 5 | ||||
| Sus scrofa | 3 | 4 | 4 | |||||||||||
| Sus sp. | 144 | 42 | 2 | 42 | ||||||||||
| Cervus elaphus | 26 | 4 | 12 | 4 | 2 | 1 | 22 | 4 | 3 | 2 | 22 | 1 | 1 | |
| Capreolus capre | 1 | |||||||||||||
| Vulpes vulpes | 1 | 1 | 1 | |||||||||||
| Oryctol. cunicul. | 16 | 15 | 2 | 4 | 62 | 13 | 4 | |||||||
| Lepus sp. | 3 | 1 | 8 | 10 | 1 | 8 | ||||||||
| Leporidae indet | 59 | 4 | 59 | |||||||||||
| Large size | 12 | 42 | 65 | 9 | 303 | |||||||||
| Intermediate size | 5 | 40 | ||||||||||||
| Small size | 20 | 59 | 8 | 161 | ||||||||||
| Carnivores | 8 | |||||||||||||
| Indet | 1824 | 98 | 27 | 84 | ||||||||||
| Total | 225 | 353 | 713 | 61 | 27 | 71 | 564 | 2949 | 158 | 220 | 266 | 565 | 62 | 708 |
| Domestic Bone Assemblages (See Data of Table 3) | Symbolic Deposits Bone Assemblages (See Data of Table 4) | |||
|---|---|---|---|---|
| %MNI | % Infant or Juvenil | %MNI | % Infant or Juvenil | |
| Bos taurus | 15.7% | 19% | 39% | 43% |
| Capra hircus | 5% | 33% | 22% | 50% |
| Sus domesticus | 12.7% | 69% | 11% | 100% |
| Canis familaris | 9.8% | 0% | 11% | 0% |
| Capra pyrenaica | 2.5% | 100% | 5.5% | 100% |
| Birds | 1.1% | 0% | 11% | 0% |
| Skeletal profiles | Skeletal profiles compensated with bones of all anatomical parts | All individual are complete and articulated | ||
| Taphonomy | Human and carnivore activity | No human or carnivore activity | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estaca-Gómez, V.; Cruz-Alcázar, R.; Tardaguila-Giacomozzi, S.; Yravedra, J. New Evidence for the Bronze Age Zooarchaeology in the Inland Area of the Iberian Peninsula through the Analysis of Pista de Motos (Villaverde Bajo, Madrid). Animals 2024, 14, 413. https://doi.org/10.3390/ani14030413
Estaca-Gómez V, Cruz-Alcázar R, Tardaguila-Giacomozzi S, Yravedra J. New Evidence for the Bronze Age Zooarchaeology in the Inland Area of the Iberian Peninsula through the Analysis of Pista de Motos (Villaverde Bajo, Madrid). Animals. 2024; 14(3):413. https://doi.org/10.3390/ani14030413
Chicago/Turabian StyleEstaca-Gómez, Verónica, Rocío Cruz-Alcázar, Silvia Tardaguila-Giacomozzi, and José Yravedra. 2024. "New Evidence for the Bronze Age Zooarchaeology in the Inland Area of the Iberian Peninsula through the Analysis of Pista de Motos (Villaverde Bajo, Madrid)" Animals 14, no. 3: 413. https://doi.org/10.3390/ani14030413