1. Introduction
In the context of the circular economy, maximizing the utilization of agricultural byproducts as substitutes for traditional feed is one of the effective strategies to alleviate feed shortages, address the issue of waste disposal, and reduce the production cost of livestock breeding [
1]. The rational development and utilization of agricultural byproducts are of great significance in achieving green industrial circulation and promoting the sustainable development of animal husbandry.
Hypsizygus marmoreus is an industrial mushroom widely cultivated in East Asia and is popular among consumers for its unique umami flavor and various medicinal effects [
2]. The dietary effect is attributed to the presence of active substances, such as polysaccharides, proteins, essential amino acids, lectins, vitamins, and enzymes, thus having potential effects on improving human health [
3]. However, the byproducts after mushroom harvesting (spent
Hypsizygus marmoreus substrate, SHMS) have not been well treated, and traditional incineration, landfilling, composting, and other treatments have caused environmental pollution and resource waste [
4]. The spent mushroom substrate contains a large amount of mycelium and exhibits nutritional properties worthy of study, such as antioxidant capacity, antibacterial and antifungal activities, and anti-aging activity [
5,
6]. Therefore, it is a new type of high-quality unconventional feed material for livestock and poultry [
7]. There have been reports on the use of the waste mushroom matrix as a livestock feed [
8]. Furthermore, researchers process and store mushroom byproducts through physical, chemical, and microbial fermentation to better utilize and preserve these resources [
9,
10]. These studies indicate that adding the waste mushroom matrix to feeds has a positive impact on animals’ production performance and improves the nutritional composition of meat products [
11].
Currently, consumers have an increasing demand for lamb and its meat products while also paying more attention to the quality and flavor of the lamb. The quality of the lamb (such as the meat color and amino acid and fatty acid contents) affects consumer choices. Nutritional regulation and feeding strategies are effective means to improve meat quality [
12]. Therefore, exploring the impact of the diet on meat quality has a guiding significance for production. On the other hand, the diet affects rumen fermentation and is closely related to the production performance and meat quality of ruminants [
13].
The rumen is a unique digestive organ in ruminants, housing a large number of microorganisms. The rumen microbiome interacts with the host in a symbiotic relationship, aiding in the adaptation to high-fiber plants and providing energy for the host’s growth through the fermentation of nutrients [
14,
15]. The dietary composition is a crucial factor affecting the rumen microbiota, and changes in these nutrients can impact the structure of the rumen microbiota community [
16]. In a previous study, it was found that adding a certain proportion of the fermented spent mushroom substrate from
Pleurotus eryngii to the diet of Hu sheep increased the diversity of the rumen microbiota and altered the microbial community’s structure [
17]. However, research on the impact of the spent mushroom substrate on the rumen microbiota of ruminants is limited, and there is a lack of studies on the application of composite alkali-stored SHMS in ruminant nutrition. This study aims to investigate the effects of composite alkali-storage SHMS on goat carcass quality, rumen fermentation, and rumen microbiota in order to provide a theoretical basis for the rational development and application of SHMS.
4. Discussion
Sensory indicators of meat quality (marble score, meat color, fat color, etc.) and intrinsic indicators (cooking loss, pH, and shear force) are commonly used for evaluating meat quality [
30]. Marble score can be used to measure the juiciness and tenderness of muscles, while meat color is the main sensory indicator that influences consumers’ purchasing decisions [
31]. Although the correlation between muscle color and meat flavor is weak, it strongly affects consumer preferences [
32]. The change in flesh color is mainly determined by the proportion of ferrimyoglobin (bright red), myoglobin (dark red), and metmyoglobin (gray, brown) in muscles [
33]. The color of fat can reflect lipid deposition in muscles [
34]. Previous studies have found that the supplementation of the spent mushroom substrates improves animal carcass quality [
35]. This study reached the same conclusion that feeding SHMS to goats can improve meat quality and increase marbling, meat color score, and fat color score. Among them, the abundance of marble score in Chuanzhong black goats is relatively low. After adding SHMS to the diet, the marble scores of MG and HG were significantly higher than those of CG. It may be because
Hypsizygus marmoreus is rich in natural antioxidants and characteristic bioactive compounds [
36], and there are still residues in SHMS after mushroom harvest. On the other hand, lipid oxidation can alter the chemical properties of heme and cause myoglobin oxidation, resulting in the loss of brown color [
37]. Antioxidants can inhibit the conversion of myoglobin in meat to metmyoglobin, protecting mutton from discoloration [
38], thus affecting the grading of meat and fat color. This indicates that the sensory index of SHMS goat muscle is superior to that of the control group. As for intrinsic indicators, cooking loss is utilized to evaluate muscle water retention during cooking [
39]. Different dietary supplements may produce variations in the tissue structure of mutton, leading to different changes in the cooking loss of longissimus dorsi [
16], a trend similar to the results of this experiment. The pH value is a physical indicator of muscle acidity and alkalinity and is related to the degradation of glycogen and the release of lactic acid before and after slaughter [
40]. It plays an important role in the biochemical processing after slaughter [
41]. Shear force is used to describe the tenderness of meat, and it is directly influenced by muscle fibers and intramuscular fat [
42]. In this study, there were no significant differences in the pH and shear force between the experimental groups, indicating that the intrinsic quality of muscles is less affected by the amount of SHMS added. Additionally, the hygiene standards for the experimental goat carcasses complied with national regulations, suggesting the feasibility of adding SHMS to the diet.
The composition of amino acids and fatty acids is closely related to the flavor of meat [
43]. Therefore, this experiment evaluated the effect of adding SHMS to goat diets on the amino acid and fatty acid composition of the meat. Previous studies have shown that the addition of fertilized spent mushroom substrate from
Pleurotus eryngii to Hu sheep can increase the amino acid content of lamb meat [
17]. In line with the results of this experiment, the addition of SHMS significantly increased the content of histidine and tyrosine in goats. Moreover, with the increase in the added amount, the content of EAAs, N-EAAs, and FAAs showed an upward trend, indicating that SHMS can help to improve the nutritional value of lamb. The fishy taste of goat meat is one of the main factors affecting consumer consumption, primarily caused by volatile compounds produced from the oxidation of saturated fatty acids (SFAs) [
44]. In this study, there was no significant difference in SFA levels among the groups, indicating that substituting 20–40% SHMS for roughage (silage corn and oat hay) in the fattening goat diet had little effect on the fishy smell of lamb meat. Furthermore, although the MG group exhibited relatively low levels of monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs), there was no significant difference in the fatty acid profile among the four groups. It is possible that certain bioactive components can influence the fatty acid composition of lamb, which needs to be further investigated.
On the other hand, the improved carcass quality of goats fed in this study may be attributed to the higher content of total volatile fatty acids (TVFAs) in the rumen. Rumen pH and volatile fatty acid (VFA) content are important indicators for evaluating ruminant fermentation [
45]. VFA accounts for 80% of the total energy required by ruminants [
46]. In this research, the addition of 30% SHMS (MG) significantly reduced the rumen pH, which may be related to changes in rumen VFAs. These VFAs are mainly produced by the fermentation of feed carbohydrates by rumen microorganisms and include short-chain fatty acids such as acetic acid, propionic acid, and butyric acid. Propionic acid in the rumen is a precursor of glucose synthesis, produced by gluconeogenesis in the liver, providing energy for the body [
47]. Acetic acid and butyric acid are precursors for fat synthesis. The former is a product of fiber degradation and the primary carbon resource for the synthesis of milk fat and body fat [
48]. The latter is converted into β-hydroxybutyric acid and participates in the citric acid cycle [
49]. This study shows that the VFA content of MG (70.26 mmol/L) is significantly higher than CG (48.77 mmol/L). This may be attributed to the use of alkaline storage methods (5% CaO and 3% urea) in this experiment. The NH
3-N produced by urea hydrolysis in the rumen stimulates the absorption of propionic acid by the rumen epithelium while providing a nitrogen source for cellulose-degrading bacteria. This process promotes cellulose degradation, leading to increased production of acetic acid and higher VFA content in the rumen [
50]. On the other hand, a high content of propionic acid and the ratio of acetic acid to propionic acid indicate a higher energy utilization rate [
51]. In this experiment, with the increase in SHMS addition, the acetic acid/propionic acid ratio gradually increased, and HG was significantly higher than CG. Additionally, NH
3-N is an important product of rumen-fermented feed protein, endogenous protein, and non-protein nitrogen, providing a nitrogen source for rumen microorganisms to synthesize bacterial protein, with an effective concentration of 5–30 mg/100 mL [
52]. In this experiment, the concentration of NH
3-N ranged from 16.30 to 25.02 mg/100 mL, falling within the normal range and proving beneficial for microbial growth, and the NH3-N content of MG is the highest, exhibiting a trend of initially increasing and then decreasing. This trend may be attributed to the limitation of carbohydrate fermentation speed, which affects the animal’s utilization of urea [
53,
54]. Therefore, this experiment suggested that 30% SHMS was a suitable addition amount; at this time, the state of TVFA, pH, and NH
3-N in the rumen was the best. In summary, based on the current research results, the supplementation of SHMS in the diet has a positive effect on promoting rumen fermentation and improving the carcass quality of goats. These results indicate that SHMS has nutritional value as ruminant feed.
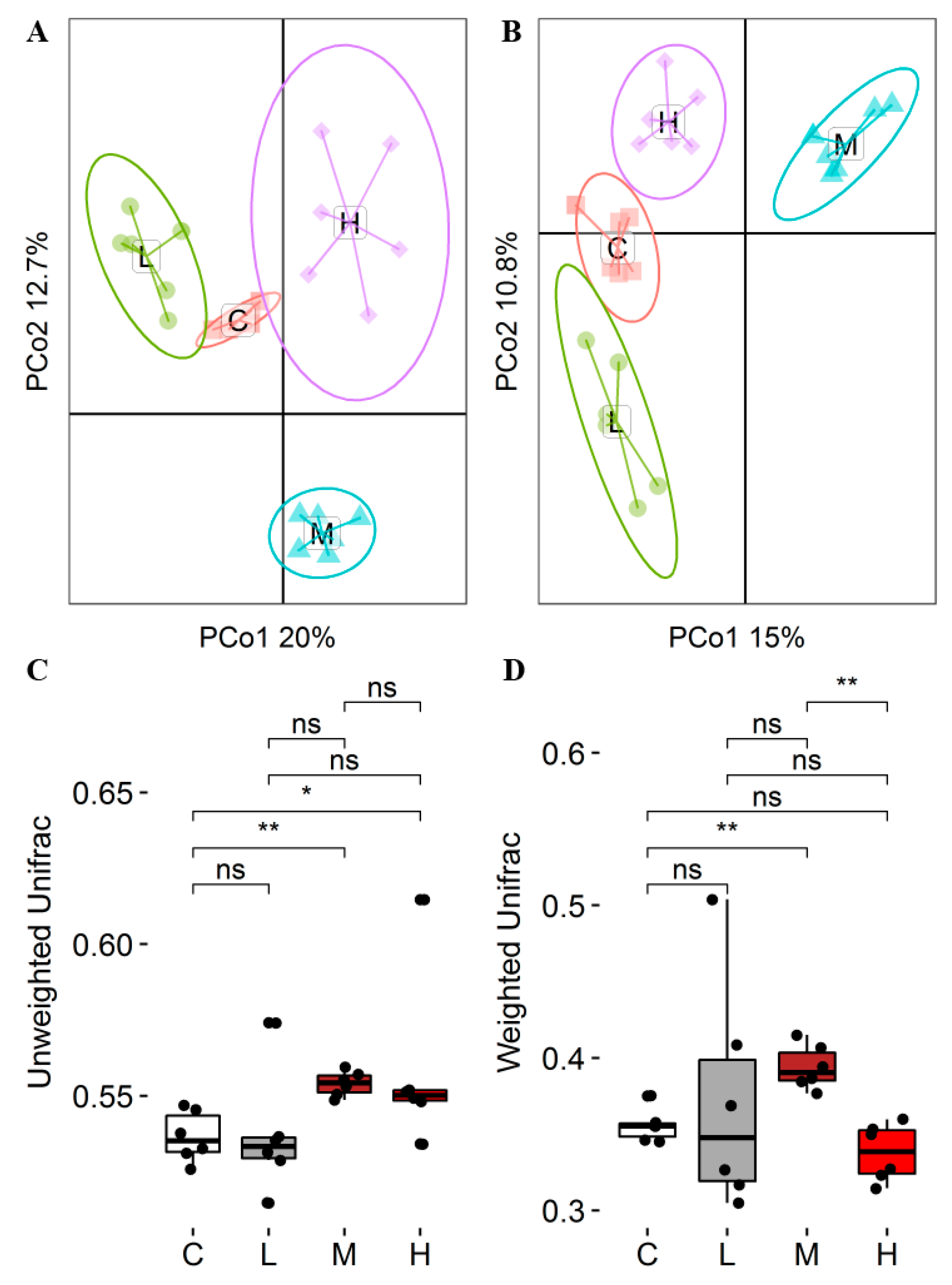
Rumen microorganisms play a crucial role in the digestion, absorption, and metabolism of nutrients in ruminants [
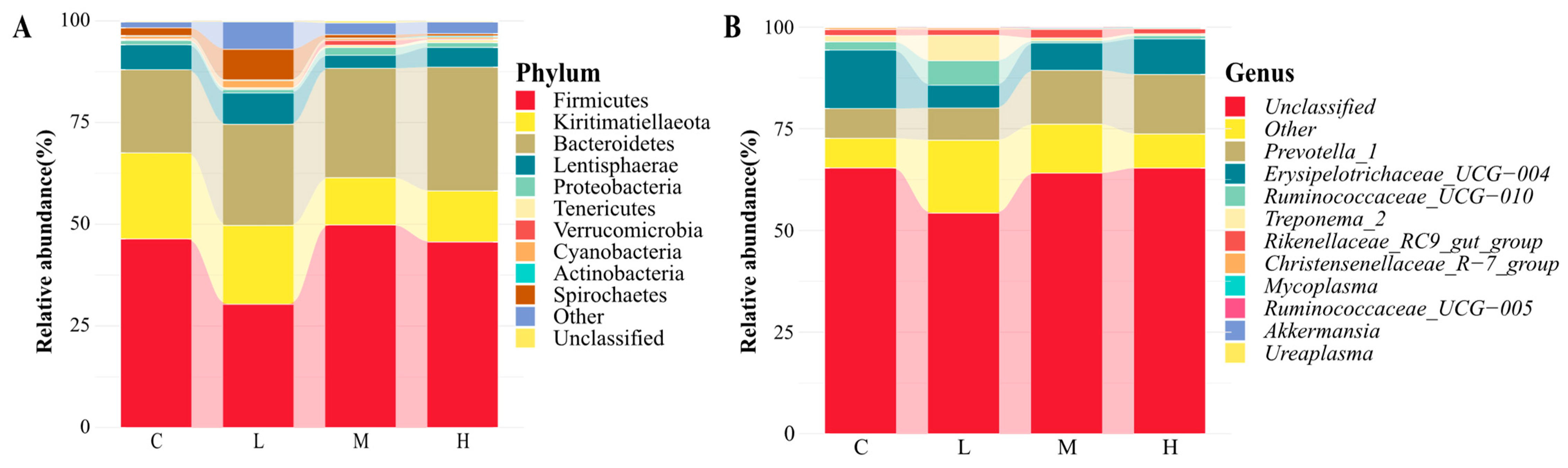
55]. This research demonstrated that the addition of SHMS resulted in changes in rumen α diversity, observed species, and Shannon index. The values of MG were significantly lower in comparison to CG. Additionally, the beta diversity analysis revealed that different levels of SHMS addition altered the rumen microbial community of goats. However, different diets did not alter the fact that Firmicutes and Bacteroidetes are the most abundant bacteria in the rumen of goats. Consistent with previous research results [
56], this experiment identified Firmicutes and Bacteroidetes as the dominant phyla. Firmicutes play a key role as cellulose decomposers. Bacteroidetes is the primary decomposer of non-fiber plant polysaccharides and proteins in the rumen, and its relative abundance is correlated with the dietary NDF level [
57,
58]. This research revealed that there was no statistically significant difference in the relative abundance of Firmicutes among the four groups. However, the content of MG Bacteroidetes was significantly higher compared to CG and LG. At the genus level, these genera were not affected by the addition of SHMS to the diet.
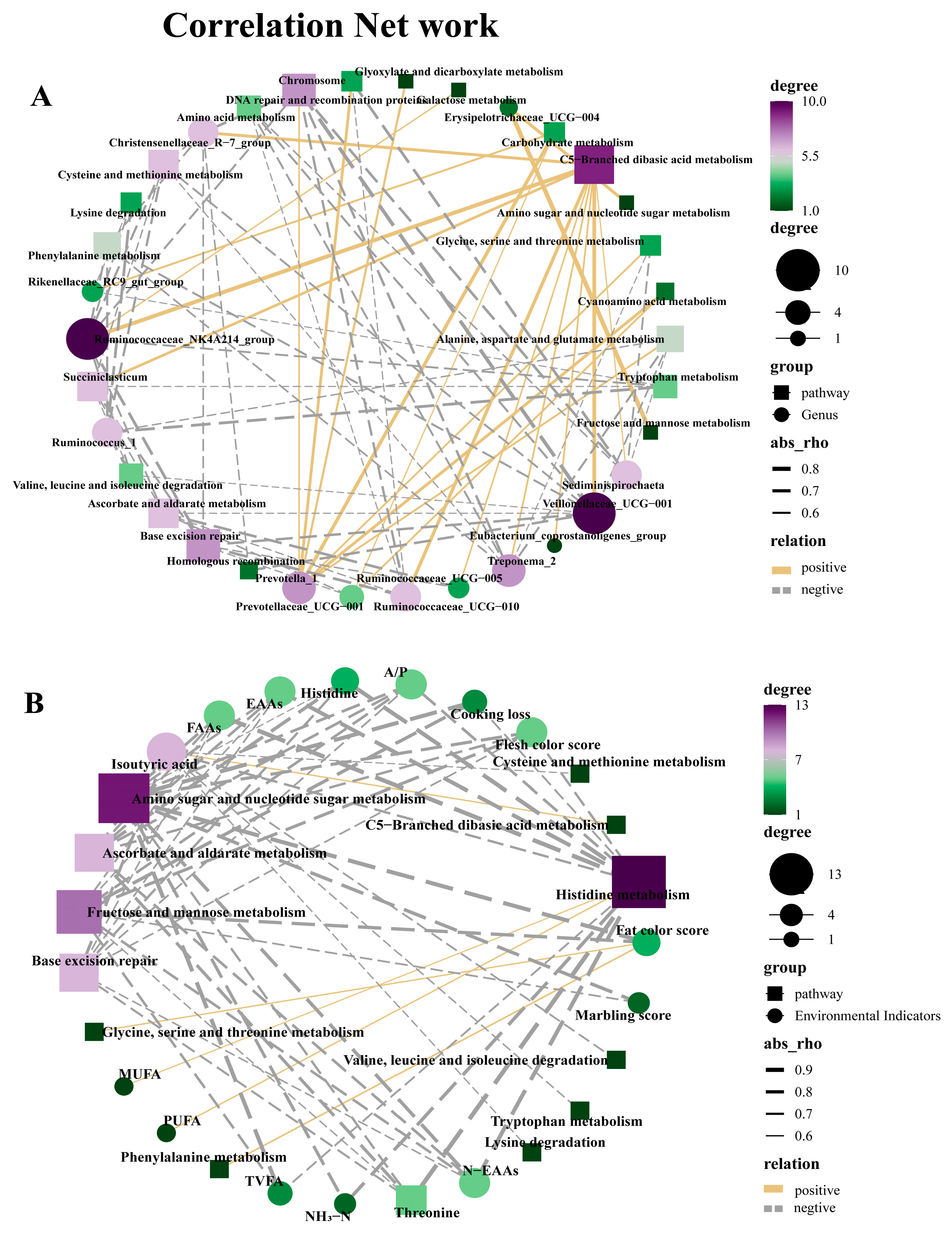
Prevotella 1 rumen is the main dominant genus in the rumen, abundant in high-fiber diets, and plays a crucial role in the degradation of high molecular weight substances such as starch and protein [
59]. The predominant bacteria at the genus level in the four groups of rumen content samples in this study was
Prevotella 1, consistent with previous studies [
60]. Among them, Prevotella exhibited the highest abundance in the MG group, which may be due to the fact that the MG after composite treatment contains more non-fibrous plant polysaccharides and non-protein nitrogen, and the C/N ratio is more suitable for the growth of
Prevotella 1. Further research is needed to determine the specific reasons.
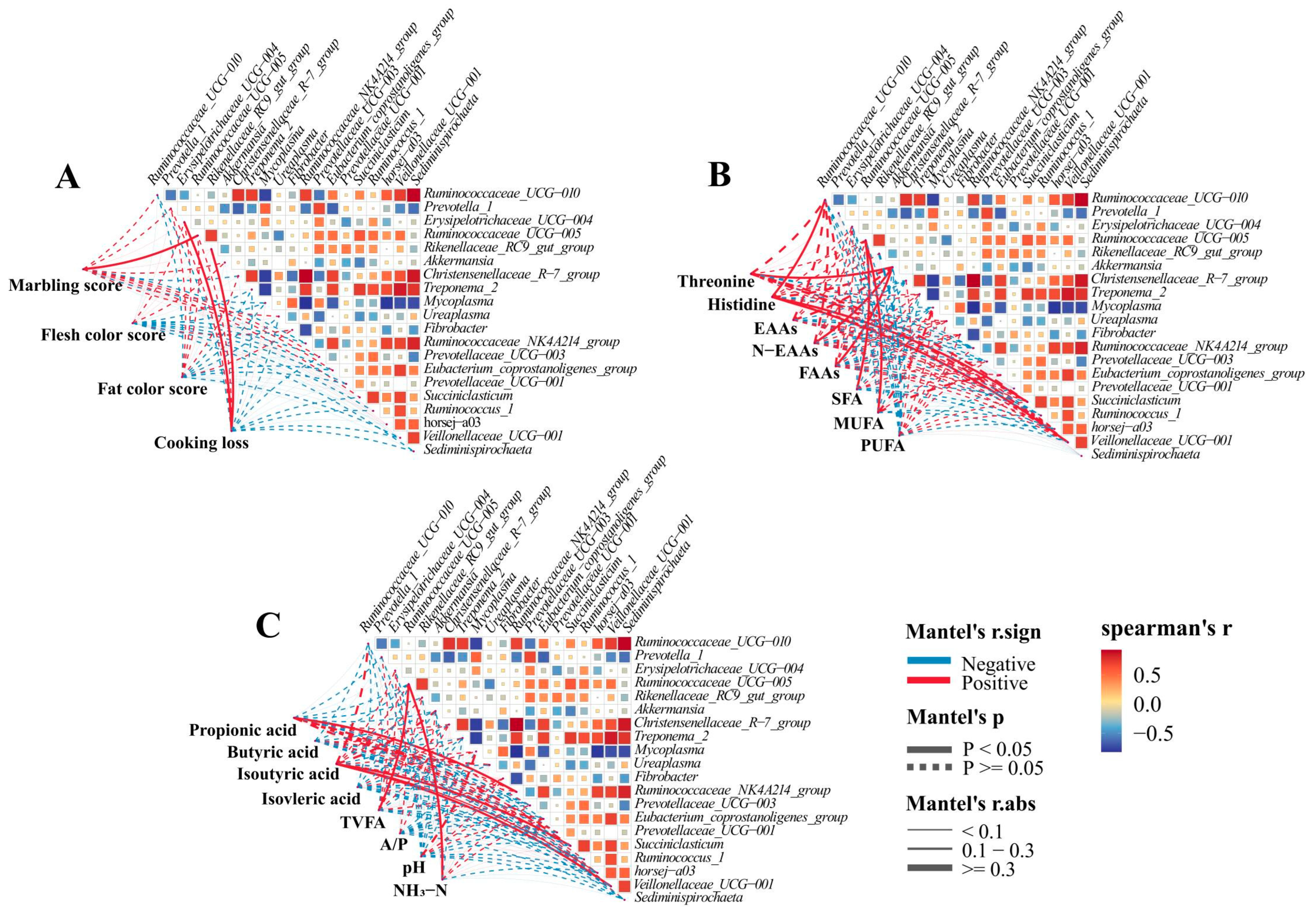
Rumen bacteria are closely related to animal carcass quality and rumen fermentation [
61]. Therefore, we evaluated whether there is a correlation between bacterial genera, carcass quality, and rumen fermentation. The
Rikenellaceae_RC9_gut_group and
Ruminococcaceae_UCG-005 play a crucial role in carbohydrate degradation, which is important for fermenting cellulose and other complex carbohydrates [
62,
63].
Erysipelotrichaceae_UCG-004 can produce metabolites that enhance acids and reduce rumen pH [
64]. In this investigation,
Ruminococcaceae_UCG-005 was positively correlated with marble score, TVFA, NH
3-N, and MUFA;
Erysipelotrichaceae_UCG-004 and
Rikenellaceae_RC9_gut_group were positively correlated with cooking loss, indicating that these bacterial genera may affect the carcass quality and rumen fermentation of goats by regulating carbohydrate degradation processes or producing metabolites. On the other hand, research has shown that
Ruminococcaceae_UCG-010,
Ruminococcaceae_NK4A214_group,
Succinniclassicum, and
Veillonellaceae can promote the degradation of cellulose and hemicellulose in animal rumen [
65,
66]. This study indicates a positive correlation between the
Ruminococcaceae_NK4A214_group and propionic acid, as well as between
Ruminococcaceae_UCG-010 and Veillonellaceae with isobutyric acid, similar to previous research results [
67]. This indicates a close correlation between rumen microbiota and VFA production. This indicates a close correlation between rumen microbiota and VFA production. These results indicate that adding SHMS to the diet can promote microbial growth and regulate rumen fermentation in goats. In addition, other studies have shown that
Alloprevotella may play an important role in the fermentation of structural carbohydrates in the rumen of goats, thereby promoting energy absorption and affecting meat quality [
68]. In research, it was also found that
Akkermansia showed a significant positive correlation with threonine, N-EAAs, and FAAs, similar to a previous study showing a positive correlation between rumen microbiota and fatty acid production [
16]. Overall, correlation analysis provides some reference for us to understand the relationships between rumen bacterial communities and carcass and rumen fermentation.
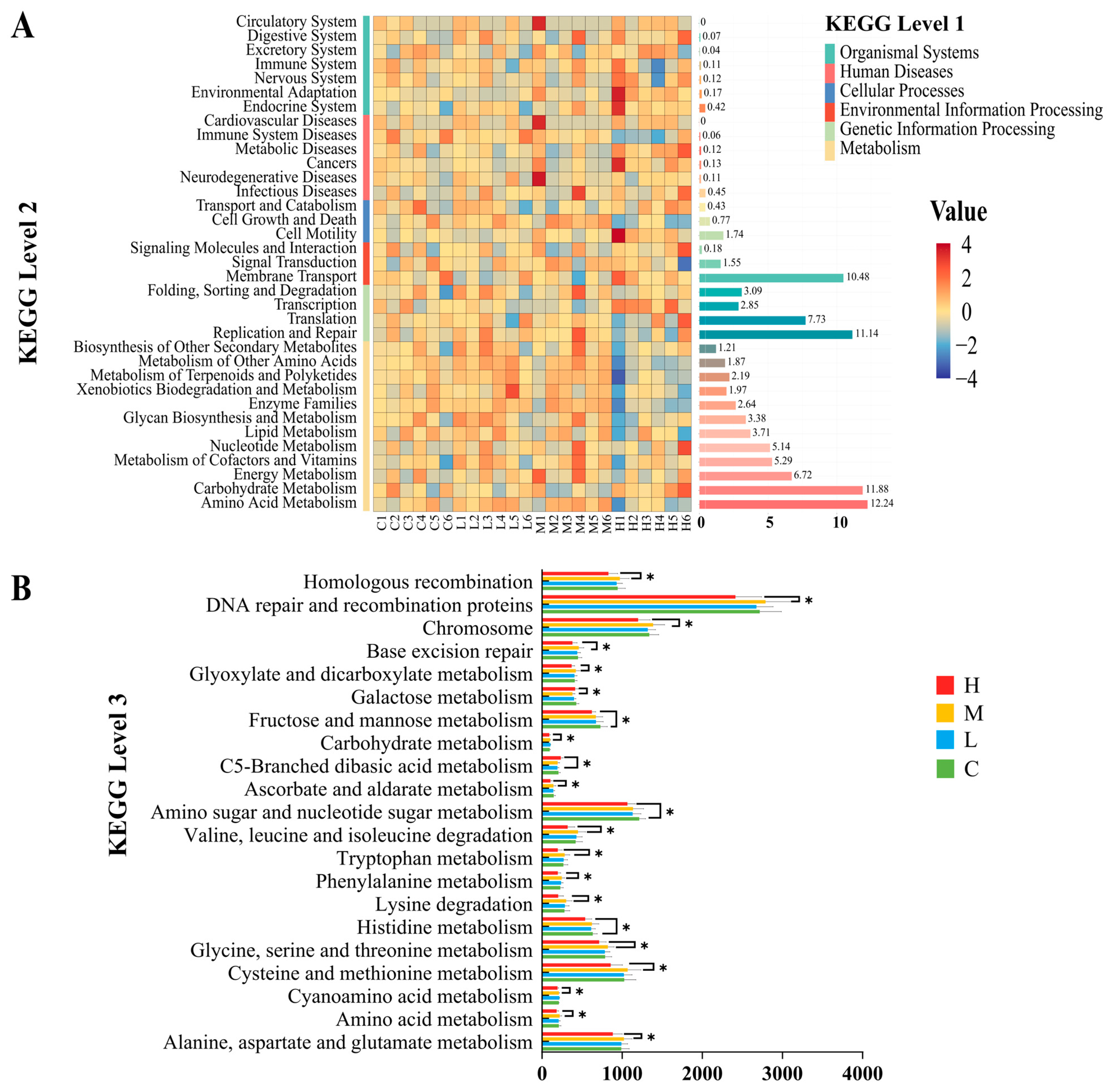
In addition, we also conducted KEGG pathway prediction and network interaction analysis between the KEGG pathway and bacterial genera, carcass quality, and rumen fermentation. It was found that at the second level, the KEGG pathways in each group were mainly concentrated in amino acid metabolism, carbohydrate metabolism, replication and repair, and membrane transport. There was no significant difference among the four groups in this experiment. However, at the third level, 21 pathways with significant differences were found, all of which showed the highest MG enrichment content and a significant decrease in HG. It is worth noting that carbohydrate metabolism and amino acid metabolism play an important role in the rumen [
69]. Carbohydrates are one of the carbon sources for rumen bacteria, especially Bacteroidetes and Firmicutes, which can decompose complex carbohydrates with the help of digestive enzymes [
70]. Previous studies have demonstrated that amino acids serve as one of the primary nitrogen sources for these bacteria, and the influence of amino acid metabolism pathways on bacterial protein synthesis and utilization is crucial [
71]. It is speculated that rumen microorganisms may indirectly affect the deposition of metabolites through interactions with the host [
72]. Previous studies have shown a close correlation between amino acid metabolism and meat quality [
73]. This research also found a close correlation between fat color score, MUFA, PUFA, and amino acid metabolism (glycine, serine, threonine, phenylalanine, and histidine). In addition, the previous experimental results showed that with the addition of SHMS, the content of these indicators showed an increasing trend in each group, which was also demonstrated in KEGG L3 prediction. However, the difference is that at 40% of SHMS addition (HG), the enrichment of metabolic pathways is significantly reduced. Adding an appropriate proportion of SHMS to the diet can increase metabolic pathways, but excessive addition can reduce their enrichment. In the network diagram between KEGG and bacterial genera, it was observed that
Prevotella 1 showed a positive correlation with seven metabolic pathways. Previous studies have highlighted Prevotella as the predominant and early colonizer, occupying various ecological niches in the rumen [
74]. These early-arriving species play a crucial role and have a long-term impact on the development of animal microbiota [
75]. In addition, it has been demonstrated that C5-Branched chain amino acid metabolism is intrinsically linked to energy generation [
76]. C5-Branched chain amino acids are converted into other metabolites through glycolysis in the rumen, thereby providing energy and nutrients for ruminants. The network graph in this experiment revealed a positive correlation between C5-Branched chain amino acid metabolism and nine bacterial genera, emphasizing the crucial role played by the microbial community in the rumen during this process. In summary, the addition of SHMS does indeed impact the rumen microbiota. Moreover, in the subsequent research, metabolites in the rumen can be examined through metabolomics to further investigate the potential mechanisms of adding SHMS to the diet on rumen bacteria, carcass quality, and rumen fermentation in Sichuan Black Mountain goats.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}