

Effect of Dietary Organic Selenium on Growth Performance, Gut Health, and Coccidiosis Response in Broiler Chickens


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chickens and Animal Care
2.2. Experimental Design, Diets, and Treatments
2.3. Growth Performance Evaluation
2.4. Sample Collection
2.5. Gut Lesion Scoring
2.6. Se Analysis
2.7. Oocyst Counts
2.8. Real-Time PCR Analysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Fecal Oocyst Shedding
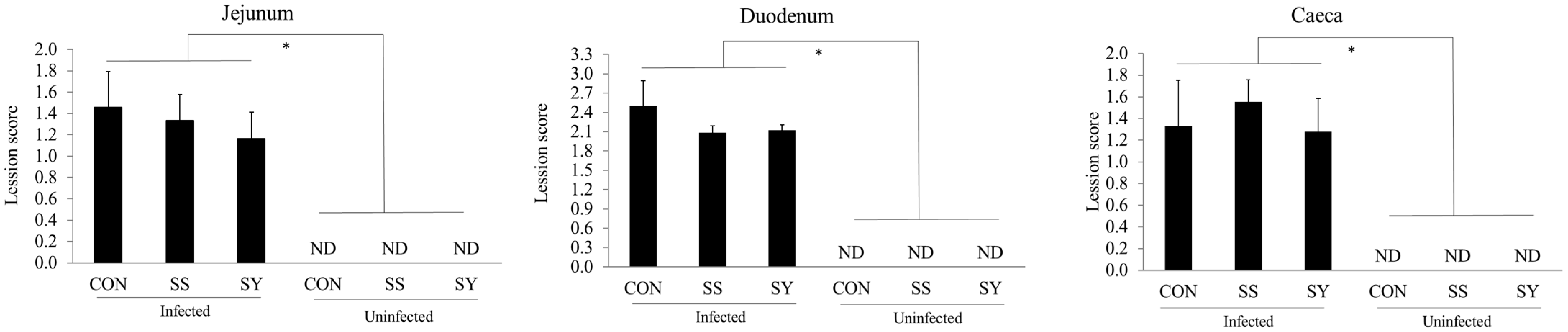
3.3. Intestinal Lesion Scores
3.4. Antioxidant Gene Expression
3.5. Tight Junction and Mucin Gene Expression
3.6. Tissue Se Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, J.H.; Kil, D.Y. Comparison of toxic effects of dietary organic or inorganic selenium and prediction of selenium intake and tissue selenium concentrations in broiler chickens using feather selenium concentrations. Poult. Sci. 2020, 99, 6462–6473. [Google Scholar] [CrossRef]
- Pardechi, A.; Tabeidian, S.A.; Habibian, M. Comparative assessment of sodium selenite, selenised yeast and nanosized elemental selenium on performance response, immunity and antioxidative function of broiler chickens. Ital. J. Anim. Sci. 2020, 19, 1108–1121. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, L.; Xu, Z.-J.; De Marco, M.; Briens, M.; Yan, X.-H.; Sun, L.-H. Hydroxy-Selenomethionine Improves the Selenium Status and Helps to Maintain Broiler Performances under a High Stocking Density and Heat Stress Conditions through a Better Redox and Immune Response. Antioxidants 2021, 10, 1542. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.L.; Rose, S.P.; Whiting, I.M.; Yovchev, D.G.; Ionescu, C.; Blanchard, A.; Pirgozliev, V. The effect of selenium source on the oxidative status and performance of broilers reared at standard and high ambient temperatures. Br. Poult. Sci. 2021, 62, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.O.; Alaqil, A.A.; Mehaisen, G.M.K.; El Sabry, M.I. Effect of Organic Selenium-Enriched Yeast on Relieving the Deterioration of Layer Performance, Immune Function, and Physiological Indicators Induced by Heat Stress. Front. Vet. Sci. 2022, 9, 880790. [Google Scholar] [CrossRef]
- Mahmoud, K.Z.; Edens, F. Influence of organic selenium on hsp70 response of heat-stressed and enteropathogenic Escherichia coli-challenged broiler chickens (Gallus gallus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 141, 69–75. [Google Scholar] [CrossRef]
- Shojadoost, B.; Kulkarni, R.R.; Yitbarek, A.; Laursen, A.; Taha-Abdelaziz, K.; Alkie, T.N.; Barjesteh, N.; Quinteiro-Filho, W.M.; Smith, T.K.; Sharif, S. Dietary selenium supplementation enhances antiviral immunity in chickens challenged with low pathogenic avian influenza virus subtype H9N2. Veter Immunol. Immunopathol. 2019, 207, 62–68. [Google Scholar] [CrossRef]
- Wickramasuriya, S.S.; Park, I.; Lee, Y.; Richer, L.M.; Przybyszewski, C.; Gay, C.G.; van Oosterwijk, J.G.; Lillehoj, H.S. Orally delivered Bacillus subtilis expressing chicken NK-2 peptide stabilizes gut microbiota and enhances intestinal health and local immunity in coccidiosis-infected broiler chickens. Poult. Sci. 2023, 102, 102590. [Google Scholar] [CrossRef]
- Wickramasuriya, S.S.; Park, I.; Lee, Y.; Kim, W.H.; Przybyszewski, C.; Gay, C.G.; van Oosterwijk, J.G.; Lillehoj, H.S. Oral Delivery of Bacillus subtilis Expressing Chicken NK-2 Peptide Protects against Eimeria acervulina Infection in Broiler Chickens. Front. Vet. Sci. 2021, 8, 684818. [Google Scholar] [CrossRef]
- Lee, Y.; Lu, M.; Lillehoj, H.S. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines 2022, 10, 215. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.; Smith, A.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef]
- Peek, H.; van der Klis, J.; Vermeulen, B.; Landman, W. Dietary protease can alleviate negative effects of a coccidiosis infection on production performance in broiler chickens. Anim. Feed. Sci. Technol. 2009, 150, 151–159. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Scapini, L.; Ribeiro, M.; Pivetta, M.; Buzim, R.; Fernandes, J. Effects of β-mannanase supplementation on the intestinal microbiota composition of broiler chickens challenged with a coccidiosis vaccine. Livest. Sci. 2019, 228, 187–194. [Google Scholar] [CrossRef]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef]
- Castro, F.L.; Teng, P.-Y.; Yadav, S.; Gould, R.L.; Craig, S.; Pazdro, R.; Kim, W.K. The effects of L-Arginine supplementation on growth performance and intestinal health of broiler chickens challenged with Eimeria spp. Poult. Sci. 2020, 99, 5844–5857. [Google Scholar] [CrossRef]
- Santos, T.; Teng, P.-Y.; Yadav, S.; Castro, F.L.D.S.; Gould, R.L.; Craig, S.W.; Chen, C.; Fuller, A.L.; Pazdro, R.; Sartori, J.R.; et al. Effects of Inorganic Zn and Cu Supplementation on Gut Health in Broiler Chickens Challenged With Eimeria spp. Front. Veter Sci. 2020, 7, 230. [Google Scholar] [CrossRef]
- El-Maddawy, Z.K.; El-Sawy, A.E.-S.F.; Ashoura, N.R.; Aboelenin, S.M.; Soliman, M.M.; Ellakany, H.F.; Elbestawy, A.R.; El-Shall, N.A. Use of Zinc Oxide Nanoparticles as Anticoccidial Agents in Broiler Chickens along with Its Impact on Growth Performance, Antioxidant Status and Hematobiochemical Profile. Life 2022, 12, 74. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Vieira, B.S.; Applegate, T.J. Influence of Dietary Zinc, Copper, and Manganese on the Intestinal Health of Broilers Under Eimeria Challenge. Front. Vet. Sci. 2020, 7, 13. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Jeong, M.; Kim, D.K.; Xu, S.; Lee, S.K.; Kim, J.B.; Park, H.J.; Kim, H.R.; et al. Immune and anti-oxidant effects of in ovo selenium proteinate on post-hatch experimental avian necrotic enteritis. Vet. Parasitol. 2014, 206, 115–122. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Jeong, M.S.; Xu, S.Z.; Kim, J.B.; Park, H.J.; Kim, H.R.; Lillehoj, E.P.; Bravo, D.M. Effects of in ovo injection with selenium on immune and antioxidant responses during experimental necrotic enteritis in broiler chickens. Poult. Sci. 2014, 93, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Lee, S.-H.; Lillehoj, H.S.; Hong, Y.H.; Bravo, D. Effects of dietary selenium on host response to necrotic enteritis in young broilers. Res. Vet. Sci. 2015, 98, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Colnago, G.; Jensen, L.; Long, P. Effect of Selenium and Vitamin E on the Development of Immunity to Coccidiosis in Chickens. Poult. Sci. 1984, 63, 1136–1143. [Google Scholar] [CrossRef]
- Johnson, J.; Reid, W. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Wahlen, R.; Evans, L.; Turner, J.; Hearn, R. The use of collision/reaction cell ICP-MS for the determination of elements in blood and serum samples. Spectroscopy 2005, 20, 84–89. [Google Scholar]
- Lee, Y.; Lee, S.; Gadde, U.; Oh, S.; Lillehoj, H. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Velichko, O.A. Selenium in poultry nutrition: From sodium selenite to organic selenium sources. J. Poult. Sci. 2017, 55, 79–93. [Google Scholar] [CrossRef]
- Park, I.; Oh, S.; Goo, D.; Celi, P.; Lillehoj, H.S. Effect of dietary sophorolipids on growth performance and gastrointestinal functionality of broiler chickens infected with Eimeria maxima. Poult. Sci. 2022, 99, 725–733. [Google Scholar] [CrossRef]
- Wang, C.-L.; Xing, G.-Z.; Wang, L.-S.; Li, S.-F.; Zhang, L.-Y.; Lu, L.; Luo, X.-G.; Liao, X.-D. Effects of selenium source and level on growth performance, antioxidative ability and meat quality of broilers. J. Integr. Agric. 2021, 20, 227–235. [Google Scholar] [CrossRef]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Jahromi, M.F.; Samsudin, A.A. The effect of dietary bacterial organic selenium on growth performance, antioxidant capacity, and Selenoproteins gene expression in broiler chickens. BMC Vet. Res. 2017, 13, 254. [Google Scholar] [CrossRef]
- Yoon, I.; Werner, T.M.; Butler, J.M. Effect of Source and Concentration of Selenium on Growth Performance and Selenium Retention in Broiler Chickens. Poult. Sci. 2007, 86, 727–730. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Xu, B.-H. Effect of different selenium source (sodium selenite and selenium yeast) on broiler chickens. Anim. Feed. Sci. Technol. 2008, 144, 306–314. [Google Scholar] [CrossRef]
- Chen, G.; Wu, J.; Li, C. Effect of different selenium sources on production performance and biochemical parameters of broilers. J. Anim. Physiol. Anim. Nutr. 2014, 98, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Upton, J.R.; Edens, F.W.; Ferket, P.R. Selenium Yeast Effect on Broiler Performance. Int. J. Poult. Sci. 2008, 7, 798–805. [Google Scholar] [CrossRef]
- Sundu, B.; Adjis, A.; Hatta, U. Effect of Addition of Selenium from Different Feedstuffs on Feed Digestibility, Growth Performance, Carcass Percentage and Meat Selenium of Broiler Chickens. Int. J. Poult. Sci. 2019, 18, 208–213. [Google Scholar] [CrossRef]
- Arnaut, P.R.; Viana, G.D.S.; da Fonseca, L.; Alves, W.J.; Muniz, J.C.L.; Pettigrew, J.E.; e Silva, F.F.; Rostagno, H.S.; Hannas, M.I. Selenium source and level on performance, selenium retention and biochemical responses of young broiler chicks. BMC Vet. Res. 2021, 17, 151. [Google Scholar] [CrossRef] [PubMed]
- Choct, M.; Naylor, A.; Reinke, N. Selenium supplementation affects broiler growth performance, meat yield and feather coverage. Br. Poult. Sci. 2004, 45, 677–683. [Google Scholar] [CrossRef]
- Park, I.; Goo, D.; Nam, H.; Wickramasuriya, S.S.; Lee, K.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.G.; Lillehoj, H.S. Effects of Dietary Maltol on Innate Immunity, Gut Health, and Growth Performance of Broiler Chickens Challenged with Eimeria maxima. Front. Vet. Sci. 2021, 8, 667425. [Google Scholar] [CrossRef]
- Mengistu, B.M.; Bitsue, H.K.; Huang, K. The Effects of Selenium-Enriched Probiotics on Growth Performance, Oocysts Shedding, Intestinal Cecal Lesion Scores, Antioxidant Capacity, and mRNA Gene Expression in Chickens Infected with Eimeria tenella. Biol. Trace Element Res. 2021, 199, 278–291. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Abdel-Baki, A.A.S.; Wunderlich, F.; Sies, H.; Al-Quraishy, S. Dietary selenium affects intestinal development of Eimeria papillata in mice. Parasitol. Res. 2014, 113, 267–274. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Park, I.; Wickramasuriya, S.S.; Ben Arous, J.; Koziol, M.-E.; Lillehoj, H.S. Co-administration of chicken IL-7 or NK-lysin peptide 2 enhances the efficacy of Eimeria elongation factor-1α vaccination against Eimeria maxima infection in broiler chickens. Poult. Sci. 2022, 101, 102013. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry. Vaccines 2022, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Nooreh, Z.; Taherpour, K.; Ghasemi, H.A.; Gharaei, M.A.; Shirzadi, H. Protective and immunostimulatory effects of in-feed preparations of an anticoccidial, a probiotic, a vitamin-selenium complex, and Ferulago angulata extract in broiler chickens infected with Eimeria species. BMC Vet. Res. 2021, 17, 307. [Google Scholar] [CrossRef]
- Georgieva, N.; Gabrashanska, M.; Koinarski, V.; Ermidou-Pollet, S. Antioxidant status in Eimeria acervulina infected chickens after dietary selenium treatment. Trace Elem. Electrolytes 2011, 28, 42–48. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I. Nutritional modulation of the antioxidant capacities in poultry: The case of selenium. Poult. Sci. 2019, 98, 4231–4239. [Google Scholar] [CrossRef]
- Oh, S.; Gadde, U.D.; Bravo, D.; Lillehoj, E.P.; Lillehoj, H.S. Growth-Promoting and Antioxidant Effects of Magnolia Bark Extract in Chickens Uninfected or Co-Infected with Clostridium perfringens and Eimeria maxima as an Experimental Model of Necrotic Enteritis. Curr. Dev. Nutr. 2018, 2, nzy009. [Google Scholar] [CrossRef]
- Briens, M.; Mercier, Y.; Rouffineau, F.; Vacchina, V.; Geraert, P.-A. Comparative study of a new organic selenium source v. seleno-yeast and mineral selenium sources on muscle selenium enrichment and selenium digestibility in broiler chickens. Br. J. Nutr. 2013, 110, 617–624. [Google Scholar] [CrossRef]
- Deng, S.; Hu, S.; Xue, J.; Yang, K.; Zhuo, R.; Xiao, Y.; Fang, R. Productive Performance, Serum Antioxidant Status, Tissue Selenium Deposition, and Gut Health Analysis of Broiler Chickens Supplemented with Selenium and Probiotics—A Pilot Study. Animals 2022, 12, 1086. [Google Scholar] [CrossRef]
- Yang, S.; Li, L.; Yu, L.; Sun, L.; Li, K.; Tong, C.; Xu, W.; Cui, G.; Long, M.; Li, P. Selenium-enriched yeast reduces caecal pathological injuries and intervenes changes of the diversity of caecal microbiota caused by Ochratoxin-A in broilers. Food Chem. Toxicol. 2020, 137, 111139. [Google Scholar] [CrossRef] [PubMed]
- Marković, R.; Ćirić, J.; Starčević, M.; Šefer, D.; Baltić, M. Effects of selenium source and level in diet on glutathione peroxidase activity, tissue selenium distribution, and growth performance in poultry. Anim. Health Res. Rev. 2018, 19, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Bień, D.; Michalczuk, M.; Szkopek, D.; Kinsner, M.; Konieczka, P. Changes in lipids metabolism indices as a result of different form of selenium supplementation in chickens. Sci. Rep. 2022, 12, 13817. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.E.; Alzawqari, M.H.; Eid, Y.Z.; Zommara, M.; Hassan, A.M.; Dawood, M.A.O. Comparing the Influences of Selenium Nanospheres, Sodium Selenite, and Biological Selenium on the Growth Performance, Blood Biochemistry, and Antioxidative Capacity of Growing Turkey Pullets. Biol. Trace Element Res. 2022, 200, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the Environment, Metabolism and Involvement in Body Functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | CON | SS | SY |
|---|---|---|---|
| Corn | 63.36 | 63.45 | 63.38 |
| Soybean Meal | 31.69 | 31.68 | 31.68 |
| Soybean Oil | 1.51 | 1.48 | 1.51 |
| Defluorinated Phosphate | 1.12 | 1.12 | 1.12 |
| Limestone | 0.51 | 0.46 | 0.50 |
| DL-Methionine | 0.38 | 0.38 | 0.38 |
| L-Lysine, HCl | 0.35 | 0.35 | 0.35 |
| Salt | 0.30 | 0.30 | 0.30 |
| Sodium Carbonate | 0.20 | 0.20 | 0.20 |
| L-Threonine | 0.17 | 0.17 | 0.17 |
| L-Valine | 0.07 | 0.07 | 0.07 |
| Choline Chloride | 0.05 | 0.05 | 0.05 |
| Mineral Mix 2 | 0.16 | 0.16 | 0.16 |
| Vitamin Premix 3 | 0.06 | 0.06 | 0.06 |
| Phytase, 5000 FTU/g | 0.02 | 0.02 | 0.02 |
| Sand | 0.05 | 0.00 | 0.00 |
| Sodium Selenite | 0.00 | 0.05 | 0.00 |
| Seleno Yeast | 0.00 | 0.00 | 0.05 |
| Calculated Nutrients | |||
| AME, kcal/kg | 3045 | 3045 | 3045 |
| Crude Protein, % | 20.61 | 20.61 | 20.61 |
| Dig. Lysine, % | 1.20 | 1.20 | 1.20 |
| Dig. Threonine, % | 0.82 | 0.82 | 0.82 |
| Dig. Methionine, % | 0.66 | 0.66 | 0.66 |
| Dig. Cysteine, % | 0.26 | 0.26 | 0.26 |
| Calcium, % | 0.90 | 0.90 | 0.90 |
| Total Phosphorus, % | 0.55 | 0.55 | 0.55 |
| Available Phosphorus, % | 0.45 | 0.45 | 0.45 |
| Analyzed Value | |||
| Selenium, mg/kg | 0.07 | 0.36 | 0.35 |
| Target Gene | Primer Sequence | Accession No. |
|---|---|---|
| GAPDH | F: 5′-GGTGGTGCTAAGCGTGTTAT-3′ | K01458 |
| R: 5′-ACCTCTGTCATCTCTCCACA-3′ | ||
| JAM-2 | F: 5′-AGCCTCAAATGGGATTGGATT-3′ | NM0,010,06257.1 |
| R: 5′-CATCAACTTGCATTCGCTTCA-3′ | ||
| OCLN | F: 5′-GAGCCCAGACTACCAAAGCAA-3′ | NM205,128.1 |
| R: 5′-GCTTGATGTGGAAGAGCTTGTTG-3′ | ||
| ZO-1 | F: 5′-CCGCAGTCGTTCACGATCT-3′ | XM01,527,8981.1 |
| R: 5′-GGAGAATGTCTGGAATGGTCTGA-3′ | ||
| MUC-2 | F: 5′-GCCTGCCCAGGAAATCAAG-3′ | NM0,013,18434.1 |
| R: 5′-CGACAAGTTTGCTGGCACAT-3′ | ||
| HMOX-1 | F: 5′-CTGGAGAAGGGTTGGCTTTCT-3′ | NM205344 |
| R: 5′-GAAGCTCTGCCTTTGGCTGTA-3′ | ||
| SOD-1 | F: 5′-ATTACCGGCTTGTCTGATGG-3′ | NM205064.1 |
| R: 5′-CCTCCCTTTGCAGTCACATT-3′ | ||
| CAT | F: 5′-ACTGCAAGGCGAAAGTGTTT-3′ | NM001031215.1 |
| R: 5′-GGCTATGGATGAAGGATGGA-3′ |
| Item | Diet 2 | SEM | Infection 3 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | SS | SY | Un-Infected | Infected | Diet | Infection | D × I | |||
| Body Weight, g | ||||||||||
| Day 16 | 689.44 a | 695.44 a | 717.80 b | 7.672 | 700.16 | 701.62 | 6.265 | 0.034 | 0.871 | 0.348 |
| Day 22 (6 dpi) | 1112.91 a | 1128.89 ab | 1152.82 b | 12.670 | 1160.67 | 1102.41 | 10.345 | 0.092 | 0.001 | 0.834 |
| Day 24 (8 dpi) | 1322.43 | 1303.94 | 1323.28 | 16.272 | 1349.92 | 1283.19 | 13.904 | 0.645 | 0.001 | 0.206 |
| Average Daily Gain, g/bird/day | ||||||||||
| Day 1–16 | 38.53 a | 38.88 a | 40.22 b | 0.436 | 39.21 | 39.21 | 0.356 | 0.025 | 0.987 | 0.369 |
| Day 16–22 | 67.42 a | 75.01 b | 72.90 b | 1.655 | 75.83 | 67.72 | 1.351 | 0.008 | 0.001 | 0.335 |
| Day 16–24 | 75.76 | 80.64 | 79.15 | 2.229 | 82.02 | 75.02 | 1.905 | 0.335 | 0.013 | 0.120 |
| Average Daily Feed Intake, g/bird/day | ||||||||||
| Day 1–16 | 33.15 | 33.13 | 34.74 | 5.978 | 34.11 | 33.23 | 4.881 | 0.976 | 0.899 | 0.995 |
| Day 16–22 | 115.61 b | 109.45 a | 112.74 a,b | 1.645 | 115.22 | 109.98 | 1.343 | 0.038 | 0.008 | 0.233 |
| Day 16–24 | 125.66 | 119.24 | 122.96 | 2.094 | 125.96 | 119.28 | 1.709 | 0.104 | 0.008 | 0.216 |
| Feed Conversion Rate, g/g | ||||||||||
| Day 1–16 | 1.24 | 1.23 | 1.24 | 0.021 | 1.25 | 1.22 | 0.017 | 0.902 | 0.224 | 0.975 |
| Day 16–22 | 1.66 b | 1.48 a | 1.53 a | 0.035 | 1.49 | 1.62 | 0.029 | 0.002 | 0.006 | 0.841 |
| Day 16–24 | 1.63 | 1.50 | 1.58 | 0.049 | 1.54 | 1.60 | 0.040 | 0.169 | 0.301 | 0.929 |
| Item | Diet 2 | SEM | Infection 3 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | SS | SY | Un-Infected | Infected | Diet | Infection | D × I | |||
| Duodenum | ||||||||||
| SOD-1 | 3.02 | 1.89 | 1.72 | 0.468 | 2.43 | 1.99 | 0.382 | 0.119 | 0.420 | 0.767 |
| CAT | 0.53 | 0.38 | 0.38 | 0.077 | 0.41 | 0.45 | 0.063 | 0.289 | 0.618 | 0.316 |
| HMOX-1 | 0.06 | 0.08 | 0.07 | 0.010 | 0.08 | 0.06 | 0.009 | 0.342 | 0.030 | 0.078 |
| Jejunum | ||||||||||
| SOD-1 | 2.95 | 4.85 | 5.51 | 1.257 | 3.55 | 5.32 | 0.954 | 0.306 | 0.193 | 0.073 |
| CAT | 0.27 | 0.26 | 0.36 | 0.049 | 0.31 | 0.29 | 0.041 | 0.352 | 0.745 | 0.963 |
| HMOX-1 | 0.04 a | 0.07 b | 0.08 b | 0.009 | 0.07 | 0.06 | 0.007 | 0.040 | 0.331 | 0.778 |
| Item | Diet 2 | SEM | Infection 3 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| CON | SS | SY | Un-Infected | Infected | Diet | Infection | D × I | |||
| Duodenum | ||||||||||
| JAM-2 | 0.072 ab | 0.013 a | 0.15 b | 0.036 | 0.034 | 0.123 | 0.029 | 0.037 | 0.058 | 0.084 |
| OCLN | 0.164 | 0.097 | 0.187 | 0.046 | 0.114 | 0.185 | 0.038 | 0.367 | 0.197 | 0.398 |
| ZO-1 | 0.207 | 0.099 | 0.341 | 0.073 | 0.151 | 0.281 | 0.062 | 0.085 | 0.136 | 0.136 |
| MUC-2 | 1.396 | 0.846 | 1.392 | 0.374 | 1.04 | 1.382 | 0.322 | 0.526 | 0.446 | 0.256 |
| Jejunum | ||||||||||
| JAM-2 | 0.048 | 0.048 | 0.048 | 0.017 | 0.047 | 0.046 | 0.014 | 0.835 | 0.936 | 0.083 |
| OCLN | 0.078 a | 0.078 a | 0.129 b | 0.013 | 0.111 | 0.079 | 0.01 | 0.008 | 0.033 | 0.137 |
| ZO-1 | 0.146 | 0.116 | 0.170 | 0.031 | 0.138 | 0.151 | 0.026 | 0.464 | 0.718 | 0.795 |
| MUC-2 | 4.465 | 4.270 | 4.955 | 0.979 | 6.205 | 2.922 | 0.799 | 0.879 | 0.008 | 0.960 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wickramasuriya, S.S.; Park, I.; Lee, Y.; Lillehoj, H.S. Effect of Dietary Organic Selenium on Growth Performance, Gut Health, and Coccidiosis Response in Broiler Chickens. Animals 2023, 13, 1560. https://doi.org/10.3390/ani13091560
Wickramasuriya SS, Park I, Lee Y, Lillehoj HS. Effect of Dietary Organic Selenium on Growth Performance, Gut Health, and Coccidiosis Response in Broiler Chickens. Animals. 2023; 13(9):1560. https://doi.org/10.3390/ani13091560
Chicago/Turabian StyleWickramasuriya, Samiru S., Inkyung Park, Youngsub Lee, and Hyun S. Lillehoj. 2023. "Effect of Dietary Organic Selenium on Growth Performance, Gut Health, and Coccidiosis Response in Broiler Chickens" Animals 13, no. 9: 1560. https://doi.org/10.3390/ani13091560