
Microsatellite-Based Genetic Diversity and Population Structure of Huacaya Alpacas (Vicugna pacos) in Southern Peru
 , ,
, ,  , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
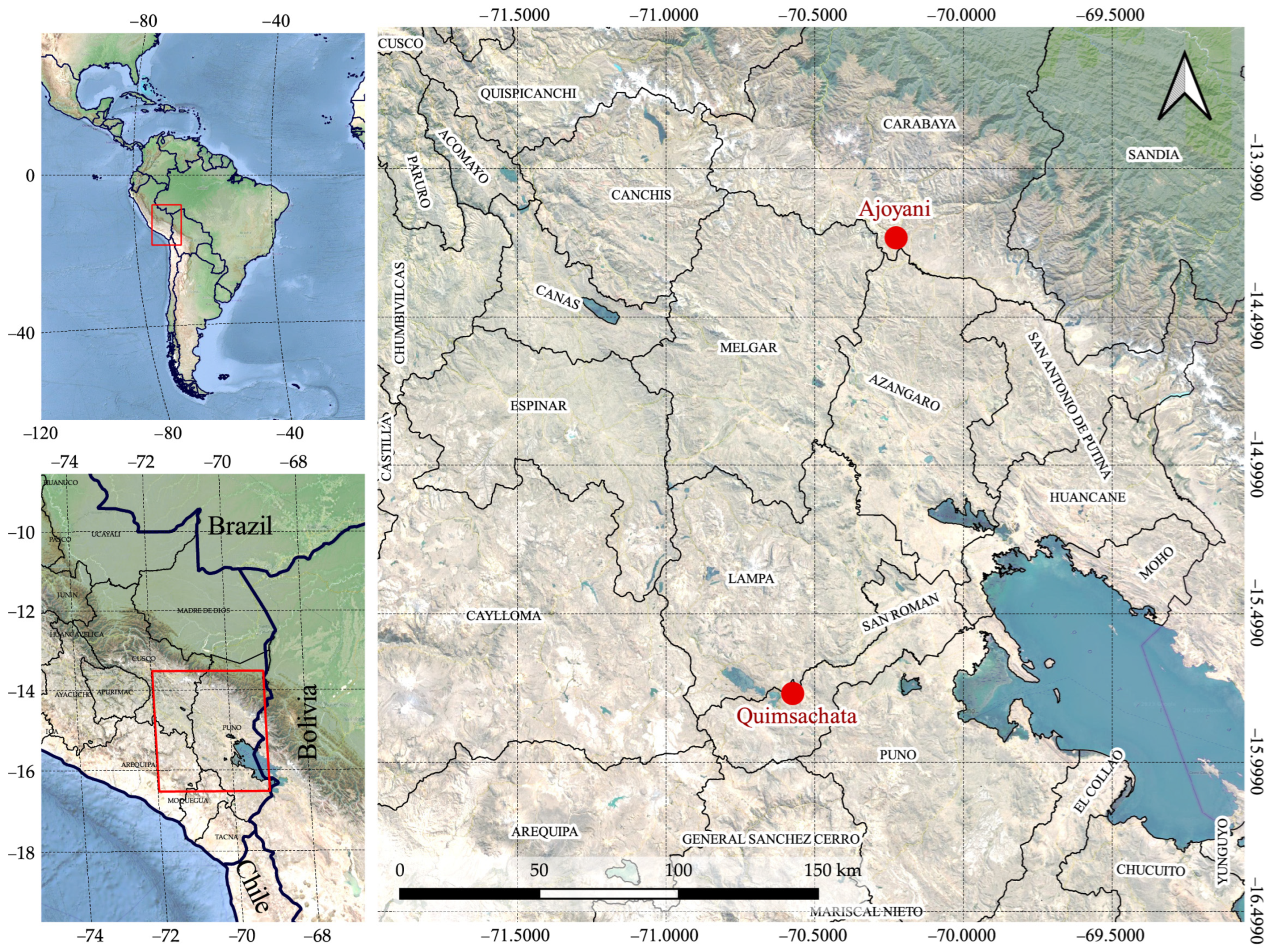
2.1. Sample Collection
2.2. Genomic DNA Extraction, and Microsatellite Genotyping
2.3. Genetic Diversity
2.4. Population Structure
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Instituto Nacional de Estadística e Informática IV Censo Nacional Agropecuario. 2012. Available online: http://censos.inei.gob.pe/Cenagro/redatam/# (accessed on 10 December 2022).
- MINAGRI. Situación de La Alpaca En El Perú; Ministerio de Agricultura y Riego-MINAGRI: Lima, Perú, 2018.
- Gutierrez, G.; Corredor, F.A.; Robles, R.; Mendoza, J.; Hidalgo, V.; Wurzinger, M. Alfalfa Hay Supplementation to Improve Llama Meat Production for Smallholders in Pasco Region. In Advances in Fibre Production Science in South American Camelids and other Fibre Animals; Universitätsverlag Göttingen: Göttingen, Germany, 2019. [Google Scholar]
- Germaná, C.; Chaquilla, O.; Santos, G.; Ferrari, M.; Krusich, C.; Kindgard, F. Estudio Socioeconómico de Los Pastores Andinos de Perú, Ecuador, Bolivia y Argentina; El Alva SRL: Apurímac, Perú, 2016. [Google Scholar]
- Gutierrez, G.; Candio, J.; Ruiz, J.; Mamani, G.; Corredor, F.-A.; Flores, E. Performance of Alpacas from a Dispersed Nucleus in Pasco Region, Peru. In Proceedings of the 64th EAAP Annual Meeting, Symposium on South American Camelids and Other Fibre Animals, Nantes, France, 25–30 August 2013. [Google Scholar]
- Machaca Machaca, V.; Bustinza Choque, A.V.; Corredor Arizapana, F.A.; Paucara Ocsa, V.; Quispe Peña, E.E.; Machaca Machaca, R. Fiber Characteristics of Huacaya Alpaca at Cotaruse, Apurímac, Perú. Rev. Investig. Vet. Perú 2017, 28, 843–851. [Google Scholar] [CrossRef]
- MINAGRI. Potencial Productivo y Comercial de La Alpaca; Ministerio de Agricultura y Riego-MINAGRI: Lima, Perú, 2019.
- Cecilio Barrantes, C.; Enrique Flores, M.; José Ruiz, C. Characterization of Genetic Nuclei of the Alpaca Production Systems of the Central Highlands of Peru. Rev. Investig. Vet. Peru 2018, 29, 1335–1348. [Google Scholar] [CrossRef]
- Vallejo, A.R.; Yalta, C.; Veli, E.; Cerna, D. Diversidad y Estructuración Genética de Alpacas de Color de La Región Puno (Perú). In Proceedings of the Congreso Mundial de Camelidos Sudamericanos, Arica, Chile, 21–23 November 2012; p. 140. [Google Scholar]
- Chambers, G.K.; MacAvoy, E.S. Microsatellites: Consensus and Controversy. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 126, 455–476. [Google Scholar] [CrossRef] [PubMed]
- Barreta, J.; Iñiguez, V.; Saavedra, V.; Romero, F.; Callisaya, A.M.; Echalar, J.; Gutiérrez-Gil, B.; Arranz, J.J. Genetic Diversity and Population Structure of Bolivian Alpacas. Small Rumin. Res. 2012, 105, 97–104. [Google Scholar] [CrossRef]
- Paredes, G.F.; Yalta-Macedo, C.E.; Gutierrez, G.A.; Veli-Rivera, E.A. Genetic Diversity and Population Structure of Llamas (Lama glama) from the Camelid Germplasm Bank—Quimsachata. Genes 2020, 11, 541. [Google Scholar] [CrossRef] [PubMed]
- Echalar, J.; Barreta, J.; Iniguez, V.; Romero, F.; Callisaya, A.M.; Saavedra, V. Intraspecific Genetic Analysis of Bolivian Alpacas and Interspecific Relationship with Llamas and Vicunas. Small Rumin. Res. 2020, 189, 106137. [Google Scholar] [CrossRef]
- Morón, J.A.; Veli, E.A.; Membrillo, A.; Paredes, M.M.; Gutiérrez, G.A. Genetic Diversity and Validation of a Microsatellite Panel for Parentage Testing for Alpacas (Vicugna pacos) on Three Peruvian Farms. Small Rumin. Res. 2020, 193. [Google Scholar] [CrossRef]
- Mesas, A.; Baldi, R.; González, B.A.; Burgi, V.; Chávez, A.; Johnson, W.E.; Marín, J.C. Past and Recent Effects of Livestock Activity on the Genetic Diversity and Population Structure of Native Guanaco Populations of Arid Patagonia. Animals 2021, 11, 1218. [Google Scholar] [CrossRef]
- Mesas, A.; Cuéllar-Soto, E.; Romero, K.; Zegers, T.; Varas, V.; González, B.A.; Johnson, W.E.; Marín, J.C. Assessing Patterns of Genetic Diversity and Connectivity among Guanacos (Lama guanicoe) in the Bolivian Chaco: Implications for Designing Management Strategies. Stud. Neotrop. Fauna Environ. 2021. [Google Scholar] [CrossRef]
- Barreta, J.; Gutiérrez-Gil, B.; Iñiguez, V.; Romero, F.; Saavedra, V.; Chiri, R.; Rodríguez, T.; Arranz, J.J. Analysis of Genetic Diversity in Bolivian Llama Populations Using Microsatellites. J. Anim. Breed. Genet. 2013, 130, 321–331. [Google Scholar] [CrossRef]
- Sharma, R.; Ahlawat, S.; Sharma, H.; Prakash, V.; Shilpa; Khatak, S.; Sawal, R.K.; Tantia, M.S. Identification of a New Indian Camel Germplasm by Microsatellite Markers Based Genetic Diversity and Population Structure of Three Camel Populations. Saudi J. Biol. Sci. 2020, 27, 1699–1709. [Google Scholar] [CrossRef] [PubMed]
- Bitaraf Sani, M.; Zare Harofte, J.; Bitaraf, A.; Esmaeilkhanian, S.; Banabazi, M.H.; Salim, N.; Teimoori, A.; Shafei Naderi, A.; Faghihi, M.A.; Burger, P.A.; et al. Genome-Wide Diversity, Population Structure and Demographic History of Dromedaries in the Central Desert of Iran. Genes 2020, 11, 599. [Google Scholar] [CrossRef]
- AlAskar, H.; Alhajeri, B.H.; Almathen, F.; Alhaddad, H. Genetic Diversity and Population Structure of Dromedary Camel-Types. J. Hered. 2020, 111, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.; Siren, D.; Hasi, S.; Jambl, T.; Ji, R. Review of Genetic Diversity in Bactrian Camel (Camelus bactrianus). Anim. Front. 2022, 12, 20–29. [Google Scholar] [CrossRef]
- Yalta, C.; Sotil, G.; Veli, E. Genetic Variability of Peruvian Alpacas (Vicugna pacos) by Using Microsatellite Markers. Salud tecnol. vet 2014, 2, 134–145. [Google Scholar] [CrossRef]
- Oliveira, D.A.; Pietrafesa, J.P.; Barbalho, M.G.d.S. Manutenção Da Biodiversidade e o Hotspots Cerrado. Caminhos Geogr. 2008, 9, 101–114. [Google Scholar] [CrossRef]
- Agapito, J.; Rodríguez, J.; Herrera-Velit, P.; Timoteo, O.; Rojas, P.; Boettcher, P.J.; García, F.; Espinoza, J.R. Parentage Testing in Alpacas (Vicugna pacos) Using Semi-Automated Fluorescent Multiplex PCRs with 10 Microsatellite Markers. Anim. Genet. 2008, 39, 201–203. [Google Scholar] [CrossRef]
- Lang, K.D.; Wang, Y.; Plante, Y. Fifteen Polymorphic Dinucleotide Microsatellites in Llamas and Alpacas. Anim. Genet. 1996, 27, 293. [Google Scholar] [CrossRef]
- Penedo, M.C.; Caetano, A.R.; Cordova, K.I. Microsatellite Markers for South American Camelids. Anim. Genet. 1998, 29, 411–412. [Google Scholar]
- Penedo, M.C.T.; Caetano, A.R.; Cordova, K. Eight Microsatellite Markers for South American Camelids. Anim. Genet. 1999, 30, 166–167. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising How the Computer Program CERVUS Accommodates Genotyping Error Increases Success in Paternity Assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A Program for Identifying Clustering Modes and Packaging Population Structure Inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.S.F. A Global Protocol for Determining Genetic Distances among Domestic Livestock Breeds. In Proceedings of the 5th World Congress of Genetic, Applied to Livestock Productions, Guelph, ON, Canada, 7–12 August 1994; pp. 501–508. [Google Scholar]
- La Manna, V.; La Terza, A.; Ghezzi, S.; Saravanaperumal, S.; Apaza, N.; Huanca, T.; Bozzi, R.; Renieri, C. Analysis of Genetic Distance between Peruvian Alpaca (Vicugna pacos) Showing Two Distinct Fleece Phenotypes, Suri and Huacaya, by Means of Microsatellite Markers. Ital. J. Anim. Sci. 2011, 10, 271–276. [Google Scholar] [CrossRef]
- Rodríguez, J.; Agapito, J.; De la Flora, I.D.; Barreto, T.; Timoteo, O.; Herrera-Velit, P.; Espinoza, J.R. Genetic Diversity in Alpaca (Vicugna pacos) Populations Using 10 Microsatellite Markers. In Proceedings of the FAO/IAEA International Symposium on Sustainable Improvement of Animal Production and Health, Vienna, Austria, 8–11 June 2009; pp. 119–120. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a Genetic Linkage Map in Man Using Restriction Fragment Length Polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Yalta, C.; Sotil, G.; Veli, E. Variabilidad Genética y Detección de Error En Filiación Utilizando Microsatélites En Dos Rebaños de Alpacas Huacaya (Vicugna pacos). Salud Tecnol. Vet. 2015, 2, 134–145. [Google Scholar] [CrossRef]
- Podbielska, A.; Piórkowska, K.; Szmatoła, T. Microsatellite-Based Genetic Structure and Hybrid Detection in Alpacas Bred in Poland. Animals 2021, 11, 2193. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Microsatellite | Ajoyani | |||
|---|---|---|---|---|
| k | Ho | He | PIC | |
| LCA5 | 8 | 0.804 | 0.756 | 0.71 |
| LCA66 | 14 | 0.673 | 0.826 | 0.802 |
| LCA94 | 7 | 0.808 | 0.785 | 0.748 |
| LCA99 | 11 | 0.769 | 0.834 | 0.804 |
| YWLL29 | 7 | 0.788 | 0.745 | 0.711 |
| YWLL40 | 6 | 0.635 | 0.626 | 0.564 |
| LCA19 | 11 | 0.788 | 0.744 | 0.708 |
| LCA8 | 11 | 0.904 | 0.845 | 0.819 |
| LCA24 | 7 | 0.75 | 0.702 | 0.654 |
| LCA37 | 13 | 0.827 | 0.849 | 0.824 |
| LCA56 | 11 | 0.827 | 0.855 | 0.83 |
| LCA65 | 12 | 0.8 | 0.877 | 0.854 |
| LGU49 | 11 | 0.885 | 0.844 | 0.818 |
| YWLL46 | 4 | 0.442 | 0.508 | 0.453 |
| TOTAL | 133 | - | - | - |
| Microsatellite | Quimsachata | |||
|---|---|---|---|---|
| k | Ho | He | PIC | |
| LCA5 | 10 | 0.802 | 0.755 | 0.713 |
| LCA66 | 10 | 0.733 | 0.744 | 0.696 |
| LCA71 | 5 | 0.595 | 0.528 | 0.457 |
| LCA77 | 17 | 0.828 | 0.859 | 0.843 |
| LCA94 | 8 | 0.81 | 0.8 | 0.766 |
| LCA99 | 13 | 0.862 | 0.832 | 0.808 |
| YWLL36 | 13 | 0.853 | 0.868 | 0.849 |
| YWLL29 | 9 | 0.759 | 0.77 | 0.739 |
| YWLL40 | 6 | 0.664 | 0.694 | 0.636 |
| YWLL44 | 15 | 0.888 | 0.871 | 0.853 |
| LCA19 | 12 | 0.724 | 0.703 | 0.673 |
| LCA8 | 11 | 0.879 | 0.848 | 0.825 |
| TOTAL | 129 | - | - | - |
| Population | k | Ho | He | PIC |
|---|---|---|---|---|
| Ajoyani | 9.5 | 0.764 ± 0.112 | 0.771 ± 0.1 | 0.736 |
| Quimsachata | 10.75 | 0.783 ± 0.087 | 0.773 ± 0.095 | 0.738 |
| Microsatellite | Ajoyani | |
|---|---|---|
| Fis | HWD | |
| LCA5 | −0.064 | NS |
| LCA66 | 0.187 | ** |
| LCA94 | −0.029 | NS |
| LCA99 | 0.078 | NS |
| YWLL29 | −0.06 | NS |
| YWLL40 | −0.014 | NS |
| LCA19 | −0.06 | NS |
| LCA8 | −0.07 | NS |
| LCA24 | −0.07 | NS |
| LCA37 | 0.026 | NS |
| LCA56 | 0.033 | NS |
| LCA65 | 0.089 | NS |
| LGU49 | −0.048 | NS |
| YWLL46 | 0.131 | NS |
| Microsatellite | Quimsachata | |
|---|---|---|
| Fis | HWD | |
| LCA5 | −0.062 | NS |
| LCA66 | 0.015 | NS |
| LCA71 | −0.127 | * |
| LCA77 | 0.037 | NS |
| LCA94 | −0.012 | NS |
| LCA99 | −0.036 | NS |
| YWLL36 | 0.017 | NS |
| YWLL29 | 0.015 | NS |
| YWLL40 | 0.043 | NS |
| YWLL44 | −0.02 | NS |
| LCA19 | −0.031 | NS |
| LCA8 | −0.038 | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa, D.; Corredor, F.-A.; Mamani-Cato, R.H.; Gallegos-Acero, R.F.; Condori-Rojas, N.; Estrada, R.; Heredia, L.; Salazar, W.; Quilcate, C.; Arbizu, C.I. Microsatellite-Based Genetic Diversity and Population Structure of Huacaya Alpacas (Vicugna pacos) in Southern Peru. Animals 2023, 13, 1552. https://doi.org/10.3390/ani13091552
Figueroa D, Corredor F-A, Mamani-Cato RH, Gallegos-Acero RF, Condori-Rojas N, Estrada R, Heredia L, Salazar W, Quilcate C, Arbizu CI. Microsatellite-Based Genetic Diversity and Population Structure of Huacaya Alpacas (Vicugna pacos) in Southern Peru. Animals. 2023; 13(9):1552. https://doi.org/10.3390/ani13091552
Chicago/Turabian StyleFigueroa, Deyanira, Flor-Anita Corredor, Ruben H. Mamani-Cato, Roberto F. Gallegos-Acero, Nicoll Condori-Rojas, Richard Estrada, Lizeth Heredia, Wilian Salazar, Carlos Quilcate, and Carlos I. Arbizu. 2023. "Microsatellite-Based Genetic Diversity and Population Structure of Huacaya Alpacas (Vicugna pacos) in Southern Peru" Animals 13, no. 9: 1552. https://doi.org/10.3390/ani13091552