

Measures of Oxidative Status Markers in Relation to Age, Sex, and Season in Sick and Healthy Captive Asian Elephants in Thailand

 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Elephants and Sample Collection
2.2. Serum Oxidative Status Markers
2.3. Fecal Glucocorticoid Metabolites
2.4. Statistical Analysis
3. Results
3.1. Oxidant Markers
3.2. Antioxidant Markers
3.3. Fecal Glucocorticoid Metabolites
3.4. Complete Blood Counts and Serum Chemistry
3.5. Correlation Matrix
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaitae, A.; Gordon, I.J.; Addison, J.; Marsh, H. Protection of Elephants and Sustainable Use of Ivory in Thailand. Oryx 2021, 56, 1–8. [Google Scholar] [CrossRef]
- Williams, C.; Tiwari, S.K.; Goswami, V.R.; De Silva, S.; Kumar, A.; Baskaran, N.; Yoganand, K.; Menon, V. Elephas maximus: The IUCN Red List of Threatened Species 2020: E.T7140A45818198 2020. Available online: https://www.iucnredlist.org/species/7140/45818198 (accessed on 30 January 2023). [CrossRef]
- Menon, V.; Tiwari, S.K. Population Status of Asian Elephants (Elephas maximus) and Key Threats. Int. Zoo Yearb. 2019, 53, 17–30. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Brown, J.L.; Thitaram, C. Welfare Assessment and Activities of Captive Elephants in Thailand. Animals 2020, 10, 919. [Google Scholar] [CrossRef]
- Pla-Ard, M.; Sukmasuang, R.; Uthairasmee, S. Some Population Characteristics and Viability of Captive Asian Elephant (Elephas maximus) in Thailand. Biodivers. J. 2022, 23, 1072–1081. [Google Scholar] [CrossRef]
- Angkawanish, T.; Boonprasert, K.; Homkong, P.; Sombutputorn, P.; Mahasawangkul, S.; Keratimanochaya, T.; Clausen, B. Elephant Health Status in Thailand: The Role of Mobile Elephant Clinic and Elephant Hospital. Gajah 2009, 31, 6. [Google Scholar]
- Long, S.Y.; Latimer, E.M.; Hayward, G.S. Review of Elephant Endotheliotropic Herpesviruses and Acute Hemorrhagic Disease. ILAR J. 2016, 56, 283–296. [Google Scholar] [CrossRef]
- Sripiboon, S.; Ditcham, W.; Vaughan-Higgins, R.; Jackson, B.; Robertson, I.; Thitaram, C.; Angkawanish, T.; Phatthanakunanan, S.; Lertwatcharasarakul, P.; Warren, K. Subclinical Infection of Captive Asian Elephants (Elephas maximus) in Thailand with Elephant Endotheliotropic Herpesvirus. Arch. Virol. 2020, 165, 397–401. [Google Scholar] [CrossRef]
- Janyamethakul, T.; Sripiboon, S.; Somgird, C.; Pongsopawijit, P.; Panyapornwithaya, V.; Klinhom, S.; Loythong, J.; Thitaram, C. Hematologic and Biochemical Reference Intervals for Captive Asian Elephants (Elephas maximus) in Thailand. Kafkas. Univ. Vet. Fak. Derg. 2017, 23, 665–669. [Google Scholar] [CrossRef]
- Kophamel, S.; Illing, B.; Ariel, E.; Difalco, M.; Skerratt, L.F.; Hamann, M.; Ward, L.C.; Méndez, D.; Munns, S.L. Importance of Health Assessments for Conservation in Noncaptive Wildlife. Conserv. Biol. 2022, 36, e13724. [Google Scholar] [CrossRef]
- Sebastiano, M.; Chastel, O.; de Thoisy, B.; Eens, M.; Costantini, D. Oxidative Stress Favours Herpes Virus Infection in Vertebrates: A Meta-Analysis. Curr. Zool. 2016, 62, 325–332. [Google Scholar] [CrossRef]
- Costantini, D.; Seeber, P.A.; Soilemetzidou, S.-E.; Azab, W.; Bohner, J.; Buuveibaatar, B.; Czirják, G.Á.; East, M.L.; Greunz, E.M.; Kaczensky, P.; et al. Physiological Costs of Infection: Herpesvirus Replication Is Linked to Blood Oxidative Stress in Equids. Sci. Rep. 2018, 8, 10347. [Google Scholar] [CrossRef]
- Hayashi, I.; Morishita, Y.; Imai, K.; Nakamura, M.; Nakachi, K.; Hayashi, T. High-Throughput Spectrophotometric Assay of Reactive Oxygen Species in Serum. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2007, 631, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Shinde, A.; Ganu, J.; Naik, P. Effect of Free Radicals and Antioxidants on Oxidative Stress: A Review. J. Dent. Allied Sci. 2012, 1, 63. [Google Scholar] [CrossRef]
- Camini, F.C.; da Silva Caetano, C.C.; Almeida, L.T.; de Brito Magalhães, C.L. Implications of Oxidative Stress on Viral Pathogenesis. Arch. Virol. 2017, 162, 907–917. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Ind. J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef]
- Puppel, K.; Kapusta, A.; Kuczyńska, B. The Etiology of Oxidative Stress in the Various Species of Animals, a Review: Etiology of Oxidative Stress in Animals. J. Sci. Food Agric. 2015, 95, 2179–2184. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Majer, A.D.; Fasanello, V.J.; Tindle, K.; Frenz, B.J.; Ziur, A.D.; Fischer, C.P.; Fletcher, K.L.; Seecof, O.M.; Gronsky, S.; Vassallo, B.G.; et al. Is There an Oxidative Cost of Acute Stress? Characterization, Implication of Glucocorticoids and Modulation by Prior Stress Experience. Proc. R. Soc. B 2019, 286, 20191698. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. Oxidative Stress Index (OSi) as a New Tool to Assess Redox Status in Dairy Cattle during the Transition Period. Animal 2013, 7, 1374–1378. [Google Scholar] [CrossRef]
- Bażanów, B.A.; Chełmecka, E.; Romuk, E.; Stygar, D.M. Basic Studies on the Oxidative Stress Markers in Two Types of Horse Breed: Semi-Isolated Population of Huculs Is Different from Commercially Used Arabian Horses. BioMed Res. Int. 2020, 2020, 7542384. [Google Scholar] [CrossRef]
- Cherubini, A.; Ruggiero, C.; Polidori, M.C.; Mecocci, P. Potential Markers of Oxidative Stress in Stroke. Free Radic. Biol. Med. 2005, 39, 841–852. [Google Scholar] [CrossRef]
- Chainy, G.B.N.; Paital, B.; Dandapat, J. An Overview of Seasonal Changes in Oxidative Stress and Antioxidant Defence Parameters in Some Invertebrate and Vertebrate Species. Scientifica 2016, 2016, 6126570. [Google Scholar] [CrossRef]
- Ho, E.; Karimi Galougahi, K.; Liu, C.-C.; Bhindi, R.; Figtree, G.A. Biological Markers of Oxidative Stress: Applications to Cardiovascular Research and Practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Jakovljevic, A.; Andric, M.; Knezevic, A.; Miletic, M.; Beljic-Ivanovic, K.; Milasin, J.; Sabeti, M. Herpesviral Infection in Periapical Periodontitis. Curr. Oral Health Rep. 2018, 5, 255–263. [Google Scholar] [CrossRef]
- Rio, D.; Stewart, A.J.; Pellegrini, N. A Review of Recent Studies on Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Bhutia, Y.; Mohanta, P.; Ghosh, A.; Sherpa, M.; Pal, R. Serum Malondialdehyde Level: Surrogate Stress Marker in the Sikkimese Diabetics. J. Nat. Sc. Biol. Med. 2011, 2, 107. [Google Scholar] [CrossRef]
- Bellanti, F.; Romano, A.D.; Buglio, A.L.; Castriotta, V.; Guglielmi, G.; Greco, A.; Serviddio, G.; Vendemiale, G. Oxidative Stress Is Increased in Sarcopenia and Associated with Cardiovascular Disease Risk in Sarcopenic Obesity. Maturitas 2018, 109, 6–12. [Google Scholar] [CrossRef]
- Taverna, M.; Marie, A.-L.; Mira, J.-P.; Guidet, B. Specific Antioxidant Properties of Human Serum Albumin. Ann. Intensive Care 2013, 3, 4. [Google Scholar] [CrossRef]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding Oxidants and Antioxidants: Classical Team with New Players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef]
- Dalvi, S.; Patil, V.; Ramraje, N. The Roles of Glutathione, Glutathione Peroxidase, Glutathione Reductase and the Carbonyl Protein in Pulmonary and Extra Pulmonary Tuberculosis. J. Clin. Diagn. Res. 2012, 6, 1462–1465. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Strycharz-Dudziak, M.; Kiełczykowska, M.; Drop, B.; Świątek, Ł.; Kliszczewska, E.; Musik, I.; Polz-Dacewicz, M. Total Antioxidant Status (TAS), Superoxide Dismutase (SOD), and Glutathione Peroxidase (GPx) in Oropharyngeal Cancer Associated with EBV Infection. Oxidative Med. Cell Longev. 2019, 2019, 5832410. [Google Scholar] [CrossRef] [PubMed]
- Langhardt, J.; Flehmig, G.; Klöting, N.; Lehmann, S.; Ebert, T.; Kern, M.; Schön, M.R.; Gärtner, D.; Lohmann, T.; Dressler, M.; et al. Effects of Weight Loss on Glutathione Peroxidase 3 Serum Concentrations and Adipose Tissue Expression in Human Obesity. Obes. Facts 2018, 11, 475–490. [Google Scholar] [CrossRef]
- Dworzański, J.; Strycharz-Dudziak, M.; Kliszczewska, E.; Kiełczykowska, M.; Dworzańska, A.; Drop, B.; Polz-Dacewicz, M. Glutathione Peroxidase (GPx) and Superoxide Dismutase (SOD) Activity in Patients with Diabetes Mellitus Type 2 Infected with Epstein-Barr Virus. PLoS ONE 2020, 15, e0230374. [Google Scholar] [CrossRef] [PubMed]
- Tápanes, R.; Vázquez, S.; González, D.; Gil, L.; Guzmán, M.G.; Bernardo, L.; Castro, O.; Kourí, G.; Martínez, G. Oxidative Stress in Adult Dengue Patients. Am. J. Trop. Med. 2004, 71, 652–657. [Google Scholar] [CrossRef]
- Araújo, A.M.; Noronha-Dutra, A.; Reis, E.A.G.; Couto, N.S.; Reis, M.G.; Ko, A.I.; Araujo, G.C.; Ribeiro, G.S.; Damião, A.O.; Athanazio, D.A.; et al. Oxidative Stress Markers Correlate with Renal Dysfunction and Thrombocytopenia in Severe Leptospirosis. Am. J. Trop. Med. Hyg. 2014, 90, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Rasool, M.; Malik, A.; Khan, K.M.; Qureshi, M.S.; Shabbir, B.; Zahid, S.; Asif, M.; Manan, A.; Rashid, S.; Khan, S.R.; et al. Assessment of Biochemical and Antioxidative Status in Patients Suffering from Dengue Fever. J. Huazhong Univ. Sci. Technol. [Med. Sci.] 2015, 35, 411–418. [Google Scholar] [CrossRef]
- Pillai, A.B.; Muthuraman, K.R.; Mariappan, V.; Belur, S.S.; Lokesh, S.; Rajendiran, S. Oxidative Stress Response in the Pathogenesis of Dengue Virus Virulence, Disease Prognosis and Therapeutics: An Update. Arch. Virol. 2019, 164, 2895–2908. [Google Scholar] [CrossRef]
- Vasquez, M.; Zuniga, M.; Rodriguez, A. Oxidative Stress and Pathogenesis in Malaria. Front. Cell Infect. Microbiol. 2021, 11, 768182. [Google Scholar] [CrossRef]
- Unalli, O.; Ozarda, Y. Stability of Hematological Analytes during 48 Hours Storage at Three Temperatures Using Cell-Dyn Hematology Analyzer. J. Med. Biochem. 2021, 40, 252–260. [Google Scholar] [CrossRef]
- Satitmanwiwat, S.; Promthep, K.; Buranaamnuay, K.; Mahasawangkul, S.; Saikhun, K. Lipid and Protein Oxidation Levels in Spermatozoa and Seminal Plasma of Asian Elephants (Elephas maximus) and Their Relationship with Semen Parameters. Reprod. Dom. Anim. 2017, 52, 283–288. [Google Scholar] [CrossRef]
- Ahmed, A.Y.; Aowda, S.A.; Hadwan, M.H. A Validated Method to Assess Glutathione Peroxidase Enzyme Activity. Chem. Pap. 2021, 75, 6625–6637. [Google Scholar] [CrossRef]
- Hadwan, M.H.; Ali, S.K. New Spectrophotometric Assay for Assessments of Catalase Activity in Biological Samples. Anal. Biochem. 2018, 542, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Kosaruk, W.; Brown, J.L.; Plangsangmas, T.; Towiboon, P.; Punyapornwithaya, V.; Silva-Fletcher, A.; Thitaram, C.; Khonmee, J.; Edwards, K.L.; Somgird, C. Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand. Animals 2020, 10, 1928. [Google Scholar] [CrossRef] [PubMed]
- Yeotikar, P.V.; Nayyar, S.; Singh, C.; Mukhopadhyay, C.S.; Kakkar, S.S.; Jindal, R. Seasonal Variation in Oxidative Stress Markers of Murrah Buffaloes in Heavy Metal Exposed Areas of Ludhiana. Indian J. Anim. Res. 2019, 53, 1310–1315. [Google Scholar] [CrossRef]
- Junqueira, V.B.C.; Barros, S.B.M.; Chan, S.S.; Rodrigues, L.; Giavarotti, L.; Abud, R.L.; Deucher, G.P. Aging and Oxidative Stress. Mol. Asp. Med. 2004, 25, 5–16. [Google Scholar] [CrossRef]
- Luo, J.; Mills, K.; le Cessie, S.; Noordam, R.; van Heemst, D. Ageing, Age-Related Diseases and Oxidative Stress: What to Do Next? Ageing Res. Rev. 2020, 57, 100982. [Google Scholar] [CrossRef]
- Phillips, M.; Cataneo, R.N.; Greenberg, J.; Gunawardena, R.; Rahbari-Oskoui, F. Increased Oxidative Stress in Younger as Well as in Older Humans. Clin. Chim. Acta 2003, 328, 83–86. [Google Scholar] [CrossRef]
- Herborn, K.A.; Daunt, F.; Heidinger, B.J.; Granroth-Wilding, H.M.V.; Burthe, S.J.; Newell, M.A.; Monaghan, P. Age, Oxidative Stress Exposure and Fitness in a Long-lived Seabird. Funct. Ecol. 2016, 30, 913–921. [Google Scholar] [CrossRef]
- Anik, M.I.; Mahmud, N.; Masud, A.A.; Khan, M.I.; Islam, M.N.; Uddin, S.; Hossain, M.K. Role of Reactive Oxygen Species in Aging and Age-Related Diseases: A Review. ACS Appl. Bio Mater. 2022, 5, 4028–4054. [Google Scholar] [CrossRef]
- Matarrese, P.; Colasanti, T.; Ascione, B.; Margutti, P.; Franconi, F.; Alessandri, C.; Conti, F.; Riccieri, V.; Rosano, G.; Ortona, E.; et al. Gender Disparity in Susceptibility to Oxidative Stress and Apoptosis Induced by Autoantibodies Specific to RLIP76 in Vascular Cells. Antioxid. Redox Signal. 2011, 15, 2825–2836. [Google Scholar] [CrossRef]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender Difference in Oxidative Stress: A New Look at the Mechanisms for Cardiovascular Diseases. J. Cell Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef]
- Sebastiano, M.; Eens, M.; Elgawad, H.A.; de Thoisy, B.; Lacoste, V.; Pineau, K.; Asard, H.; Chastel, O.; Costantini, D. Oxidative Stress Biomarkers Are Associated with Visible Clinical Signs of a Disease in Frigatebird Nestlings. Sci. Rep. 2017, 7, 1599. [Google Scholar] [CrossRef]
- Costantini, D.; Czirják, G.Á.; Melzheimer, J.; Menges, V.; Wachter, B. Sex and Species Differences of Stress Markers in Sympatric Cheetahs and Leopards in Namibia. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 227, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Molinari, L.; Basini, G.; Ramoni, R.; Bussolati, S.; Aldigeri, R.; Grolli, S.; Bertini, S.; Quintavalla, F. Evaluation of Oxidative Stress Parameters in Healthy Saddle Horses in Relation to Housing Conditions, Presence of Stereotypies, Age, Sex and Breed. Processes 2020, 8, 1670. [Google Scholar] [CrossRef]
- Bhat, S.; Rao, G.; Murthy, K.D.; Bhat, P.G. Seasonal Variations in Markers of Stress and Oxidative Stress in Rats. Indian J. Clin. Biochem. 2008, 23, 191–194. [Google Scholar] [CrossRef]
- Valeanu, S.; Johannisson, A.; Lundeheim, N.; Morrell, J.M. Seasonal Variation in Sperm Quality Parameters in Swedish Red Dairy Bulls Used for Artificial Insemination. Livest. Sci. 2015, 173, 111–118. [Google Scholar] [CrossRef]
- Maibam, U.; Hooda, O.K.; Sharma, P.S.; Upadhyay, R.C.; Mohanty, A.K. Differential Level of Oxidative Stress Markers in Skin Tissue of Zebu and Crossbreed Cattle during Thermal Stress. Livest. Sci. 2018, 207, 45–50. [Google Scholar] [CrossRef]
- Kowalska, K.; Milnerowicz, H. The Influence of Age and Gender on the pro/Antioxidant Status in Young Healthy People. Ann. Clin. Lab. Sci. 2016, 46, 9. [Google Scholar]
- González, N.T.; Otali, E.; Machanda, Z.; Muller, M.N.; Wrangham, R.; Thompson, M.E. Urinary Markers of Oxidative Stress Respond to Infection and Late-Life in Wild Chimpanzees. PLoS ONE 2020, 15, e0238066. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Pizzimenti, S.; Daga, M.; Dianzani, C.; Arcaro, A.; Cetrangolo, G.P.; Giordano, G.; Cucci, M.A.; Graf, M.; Gentile, F. Lipid Peroxidation-Derived Aldehydes, 4-Hydroxynonenal and Malondialdehyde in Aging-Related Disorders. Antioxidants 2018, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid Peroxidation as Measured by Chromatographic Determination of Malondialdehyde. Human Plasma Reference Values in Health and Disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef]
- Brown, J.L.; Paris, S.; Prado-Oviedo, N.A.; Meehan, C.L.; Hogan, J.N.; Morfeld, K.A.; Carlstead, K. Reproductive Health Assessment of Female Elephants in North American Zoos and Association of Husbandry Practices with Reproductive Dysfunction in African Elephants (Loxodonta africana). PLoS ONE 2016, 11, e0145673. [Google Scholar] [CrossRef]
- Li, M.; Hassan, F.; Guo, Y.; Tang, Z.; Liang, X.; Xie, F.; Peng, L.; Yang, C. Seasonal Dynamics of Physiological, Oxidative and Metabolic Responses in Non-Lactating Nili-Ravi Buffaloes under Hot and Humid Climate. Front. Vet. Sci. 2020, 7, 622. [Google Scholar] [CrossRef]
- Yehia, S.G.; Ramadan, E.S.; Megahed, E.A.; Salem, N.Y. Influence of Season and Lactation Stage on Oxidative, Haematological, and Metabolic Profile Parameters in Holstein Dairy Cows. Adv. Anim. Vet. Sci. 2021, 9, 2095–2102. [Google Scholar] [CrossRef]
- Weaving, G.; Batstone, G.F.; Jones, R.G. Age and Sex Variation in Serum Albumin Concentration: An Observational Study. Ann. Clin. Biochem. 2016, 53, 106–111. [Google Scholar] [CrossRef]
- Reijnierse, E.M.; Trappenburg, M.C.; Leter, M.J.; Sipilä, S.; Stenroth, L.; Narici, M.V.; Hogrel, J.Y.; Butler-Browne, G.; McPhee, J.S.; Pääsuke, M.; et al. Serum Albumin and Muscle Measures in a Cohort of Healthy Young and Old Participants. Age 2015, 37, 88. [Google Scholar] [CrossRef] [PubMed]
- Atata, J.A.; Esievo, K.A.N.; Adamu, S.; Abdulsalam, H.; Avazi, D.O.; Ajadi, A.A. Haemato-Biochemical Studies of Dogs with Haemorrhage-Induced Dehydration. Comp. Clin. Pathol. 2019, 28, 129–135. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Brown, J.L.; Thitaram, C.; Punyapornwithaya, V.; Somgird, C.; Edwards, K.L.; Nganvongpanit, K. Changing Trends in Elephant Camp Management in Northern Thailand and Implications for Welfare. PeerJ 2018, 6, e5996. [Google Scholar] [CrossRef]
- Smuts, M.P.; de Bruyn, S.; Thompson, P.N.; Holm, D.E. Serum Albumin Concentration of Donor Cows as an Indicator of Developmental Competence of Oocytes. Theriogenology 2019, 125, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Høstmark, A.T.; Tomten, S.E.; Berg, J.E. Serum Albumin and Blood Pressure: A Population-Based, Cross-Sectional Study. J. Hypertens. 2005, 23, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Usvyat, L.A.; Carter, M.; Thijssen, S.; Kooman, J.P.; van der Sande, F.M.; Zabetakis, P.; Balter, P.; Levin, N.W.; Kotanko, P. Seasonal Variations in Mortality, Clinical, and Laboratory Parameters in Hemodialysis Patients: A 5-Year Cohort Study. Clin. J. Am. Soc. Nephrol. 2012, 7, 108–115. [Google Scholar] [CrossRef]
- Gocho, Y.; Tanaka, M.; Sugawara, H.; Furuhashi, M.; Moniwa, N.; Yamashita, T.; Takizawa, H.; Mukai, H.; Ohno, K.; Maeda, T.; et al. Seasonal Variation of Serum 25-Hydroxyvitamin D Level in Hemodialysis Patients in the Northernmost Island of Japan. Clin. Exp. Nephrol. 2021, 25, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- Shrikhande, G.B.; Rode, A.M.; Pradhan, M.S.; Satpute, A.K. Seasonal Effect on the Composition of Blood in Cattle. Vet. World 2008, 1, 341–342. [Google Scholar]
- Hurst, E.A.; Homer, N.Z.; Gow, A.G.; Clements, D.N.; Evans, H.; Gaylor, D.; Campbell, S.; Handel, I.; Mellanby, R.J. Vitamin D Status Is Seasonally Stable in Northern European Dogs. Vet. Clin. Pathol. 2020, 49, 279–291. [Google Scholar] [CrossRef]
- AL-Eissa, M.S.; Saad, A.; Al Farraj, S.A.; Saud, A.A.; Al Dahmash, B.; Hamad, A.Y. Seasonal Variation Effects on the Composition of Blood in Nubian Ibex (Capra nubiana) in Saudi Arabia. Afr. J. Biotechnol. 2012, 11, 1283–1286. [Google Scholar] [CrossRef]
- Yanai, M.; Satomura, A.; Uehara, Y.; Murakawa, M.; Takeuchi, M.; Kumasaka, K. Circannual Rhythm of Laboratory Test Parameters among Chronic Haemodialysis Patients. Blood Purif. 2008, 26, 196–203. [Google Scholar] [CrossRef]
- Guinsburg, A.M.; Usvyat, L.A.; Etter, M.; Xu, X.; Thijssen, S.; Marcelli, D.; Canaud, B.; Marelli, C.; Barth, C.; Wang, Y.; et al. Seasonal Variations in Mortality and Clinical Indicators in International Hemodialysis Populations from the MONDO Registry. BMC Nephrol. 2015, 16, 139. [Google Scholar] [CrossRef]
- Pérez-Severiano, F.; Santamaría, A.; Pedraza-Chaverri, J.; Medina-Campos, O.N.; Ríos, C.; Segovia, J. Increased Formation of Reactive Oxygen Species, but No Changes in Glutathione Peroxidase Activity, in Striata of Mice Transgenic for the Huntington’s Disease Mutation. Neurochem. Res. 2004, 29, 729–733. [Google Scholar] [CrossRef]
- Jimenez, A.G.; O’Connor, E.S.; Elliott, K.H. Muscle Myonuclear Domain, but Not Oxidative Stress, Decreases with Age in a Long-Lived Seabird with High Activity Costs. J. Exp. Biol. 2019, 222, jeb211185. [Google Scholar] [CrossRef]
- Darbaz, İ.; Salar, S.; Sayiner, S.; Baştan, İ.; Ergene, O.; Baştan, A. Evaluation of Milk Glutathione Peroxidase and Superoxide Dismutase Levels in Subclinical Mastitis in Damascus Goats. Turk. J. Vet. Anim. Sci. 2019, 43, 259–263. [Google Scholar] [CrossRef]
- Pastori, D.; Pignatelli, P.; Farcomeni, A.; Menichelli, D.; Nocella, C.; Carnevale, R.; Violi, F. Aging-related Decline of Glutathione Peroxidase 3 and Risk of Cardiovascular Events in Patients with Atrial Fibrillation. J. Am. Heart Assoc. 2016, 5, e003682. [Google Scholar] [CrossRef]
- Kozakiewicz, M.; Kornatowski, M.; Krzywińska, O.; Kędziora-Kornatowska, K. Changes in the Blood Antioxidant Defense of Advanced Age People. Clin. Interv. Aging 2019, 14, 763–771. [Google Scholar] [CrossRef]
- Espinoza, S.E.; Guo, H.; Fedarko, N.; DeZern, A.; Fried, L.P.; Xue, Q.-L.; Leng, S.; Beamer, B.; Walston, J.D. Glutathione Peroxidase Enzyme Activity in Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Chusyd, D.E.; Ackermans, N.L.; Austad, S.N.; Hof, P.R.; Mielke, M.M.; Sherwood, C.C.; Allison, D.B. Aging: What We Can Learn from Elephants. Front. Aging 2021, 2, 726714. [Google Scholar] [CrossRef]
- Hermsdorff, H.H.M.; Barbosa, K.B.; Volp, A.C.P.; Puchau, B.; Bressan, J.; Zulet, M.Á.; Martínez, J.A. Gender-Specific Relationships between Plasma Oxidized Low-Density Lipoprotein Cholesterol, Total Antioxidant Capacity, and Central Adiposity Indicators. Eur. J. Prev. Cardiolog. 2014, 21, 884–891. [Google Scholar] [CrossRef]
- Tomsič, K.; Seliškar, A.; Lukanc, B.; Nemec Svete, A. Plasma Total Antioxidant Capacity and Activities of Blood Glutathione Peroxidase and Superoxide Dismutase Determined in Healthy Dogs by Using Commercially Available Kits. Acta Vet. 2016, 66, 534–548. [Google Scholar] [CrossRef]
- Pilarczyk, B.; Drozd, R.; Pilarczyk, R.; Tomza-Marciniak, A.; Jankowiak, D.; Hendzel, D.; Kuba, J.; Kowalska, J. Glutathione Peroxidase (GSHPx) Activity in the Liver of Red Deer in Relation to Hepatic Selenium Concentrations, Sex, Body Weight and Season of the Year. Biol. Trace. Elem. Res. 2011, 144, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, M.K.; Koiri, R.K.; Srivastava, R. Expression of Estrogen Receptor Alpha in Response to Stress and Estrogen Antagonist Tamoxifen in the Shell Gland of Gallus gallus domesticus: Involvement of Anti-Oxidant System and Estrogen. Stress 2021, 24, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Casado, A.; López-Fernández, M.E. Age-Correlated Changes of the Erythrocyte Catalase Activity in the Spanish Population. Gerontology 2003, 49, 251–254. [Google Scholar] [CrossRef]
- Ide, T.; Tsutsui, H.; Ohashi, N.; Hayashidani, S.; Suematsu, N.; Tsuchihashi, M.; Tamai, H.; Takeshita, A. Greater Oxidative Stress in Healthy Young Men Compared with Premenopausal Women. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 438–442. [Google Scholar] [CrossRef]
- Tsuber, V.; Kadamov, Y.; Tarasenko, L. Activation of Antioxidant Defenses in Whole Saliva by Psychosocial Stress Is More Manifested in Young Women than in Young Men. PLoS ONE 2014, 9, e115048. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Ukita, H.; Kawahara, M.; Mitani, T.; Furukawa, E.; Yanagawa, Y.; Yabuuchi, N.; Kim, H.; Takahashi, M. Effect of Summer Heat Stress on Gene Expression in Bovine Uterine Endometrial Tissues. Anim. Sci. J. 2020, 91, e13474. [Google Scholar] [CrossRef] [PubMed]
- Kodydková, J.; Vávrová, L.; Kocík, M.; Žák, A. Human Catalase, Its Polymorphisms, Regulation and Changes of Its Activity in Different Diseases. Folia Biol. 2014, 60, 15. [Google Scholar]
- Seltmann, M.W.; Ukonaho, S.; Reichert, S.; Dos Santos, D.; Nyein, U.K.; Htut, W.; Lummaa, V. Faecal Glucocorticoid Metabolites and H/L Ratio Are Related Markers of Stress in Semi-Captive Asian Timber Elephants. Animals 2020, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Bansiddhi, P.; Brown, J.L.; Khonmee, J.; Norkaew, T.; Nganvongpanit, K.; Punyapornwithaya, V.; Angkawanish, T.; Somgird, C.; Thitaram, C. Management Factors Affecting Adrenal Glucocorticoid Activity of Tourist Camp Elephants in Thailand and Implications for Elephant Welfare. PLoS ONE 2019, 14, e0221537. [Google Scholar] [CrossRef] [PubMed]
- Mumby, H.S.; Mar, K.U.; Thitaram, C.; Courtiol, A.; Towiboon, P.; Min-Oo, Z.; Htut-Aung, Y.; Brown, J.L.; Lummaa, V. Stress and Body Condition Are Associated with Climate and Demography in Asian Elephants. Conserv. Physiol. 2015, 3, cov030. [Google Scholar] [CrossRef] [PubMed]
- Norkaew, T.; Brown, J.L.; Bansiddhi, P.; Somgird, C.; Thitaram, C.; Punyapornwithaya, V.; Punturee, K.; Vongchan, P.; Somboon, N.; Khonmee, J. Body Condition and Adrenal Glucocorticoid Activity Affects Metabolic Marker and Lipid Profiles in Captive Female Elephants in Thailand. PLoS ONE 2018, 13, e0204965. [Google Scholar] [CrossRef]
- Boonprasert, K.; Yun, Y.; Kosaruk, W.; Towiboon, P.; Tankaew, P.; Punyapornwithaya, V.; Janyamathakul, T.; Muanghong, P.; Brown, J.L.; Thitaram, C.; et al. A Longitudinal Study of Hematology and Stress Biomarker Profiles in Young Asian Elephants (Elephas maximus) in Relation to Elephant Endotheliotropic Herpesvirus (EEHV) in Thailand. Animals 2021, 11, 2530. [Google Scholar] [CrossRef]
- Crawley, J.A.H.; Liehrmann, O.; Franco dos Santos, D.J.; Brown, J.; Nyein, U.K.; Aung, H.H.; Htut, W.; Oo, Z.M.; Seltmann, M.W.; Webb, J.L.; et al. Influence of Handler Relationships and Experience on Health Parameters, Glucocorticoid Responses and Behaviour of Semi-Captive Asian Elephants. Conserv. Physiol. 2021, 9, coaa116. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.S.; Edwards, K.L.; Paris, S.; Scarlata, C.; Lee, B.; Wielebnowski, N.; Finnell, S.; Somgird, C.; Brown, J.L. Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events. Animals 2022, 12, 1332. [Google Scholar] [CrossRef]
- Supanta, J.; Brown, J.L.; Bansiddhi, P.; Thitaram, C.; Punyapornwithaya, V.; Khonmee, J. Effect of the COVID-19 Pandemic and International Travel Ban on Elephant Tourist Camp Management in Northern Thailand. Front. Vet. Sci. 2022, 9, 1038855. [Google Scholar] [CrossRef] [PubMed]
- Valle, L.; DuqueVizcaino, M.; CalasHechavarria, V.; GravierHernandez, R.; JimenezPerez, N.; BermudezAlfonso, Y.; HernandezRequejo, D.; GonzalezRubio, D. Multivariate Discriminant Analysis of Redox and Progression Indexes in Cuban AIDS Patients with Kaposis Sarcoma. Oxid. Antioxid. Med. Sci. 2017, 6, 1–8. [Google Scholar] [CrossRef]
- Incani, A.; Marras, L.; Serreli, G.; Ingianni, A.; Pompei, R.; Deiana, M.; Angius, F. Human Herpesvirus 8 Infection May Contribute to Oxidative Stress in Diabetes Type 2 Patients. BMC Res. Notes 2020, 13, 75. [Google Scholar] [CrossRef]
- Guntawang, T.; Sittisak, T.; Kochagul, V.; Srivorakul, S.; Photichai, K.; Boonsri, K.; Janyamethakul, T.; Boonprasert, K.; Langkaphin, W.; Thitaram, C.; et al. Pathogenesis of Hemorrhagic Disease Caused by Elephant Endotheliotropic Herpesvirus (EEHV) in Asian Elephants (Elephas maximus). Sci. Rep. 2021, 11, 12998. [Google Scholar] [CrossRef]
- Yun, Y.; Sripiboon, S.; Pringproa, K.; Chuammitri, P.; Punyapornwithaya, V.; Boonprasert, K.; Tankaew, P.; Angkawanish, T.; Namwongprom, K.; Arjkumpa, O.; et al. Clinical Characteristics of Elephant Endotheliotropic Herpesvirus (EEHV) Cases in Asian Elephants (Elephas maximus) in Thailand during 2006–2019. Vet. Q. 2021, 41, 268–279. [Google Scholar] [CrossRef]
- Kim, S.; Kang, S. Serum Albumin Levels: A Simple Answer to a Complex Problem? Are We on the Right Track of Assessing Metabolic Syndrome? Endocrinol. Metab. 2013, 28, 17. [Google Scholar] [CrossRef]
- Perrin, K.L.; Kristensen, A.T.; Bertelsen, M.F.; Denk, D. Retrospective Review of 27 European Cases of Fatal Elephant Endotheliotropic Herpesvirus-Haemorrhagic Disease Reveals Evidence of Disseminated Intravascular Coagulation. Sci. Rep. 2021, 11, 14173. [Google Scholar] [CrossRef]
- Crnogaj, M.; Cerón, J.J.; Šmit, I.; Kiš, I.; Gotić, J.; Brkljačić, M.; Matijatko, V.; Rubio, C.P.; Kučer, N.; Mrljak, V. Relation of Antioxidant Status at Admission and Disease Severity and Outcome in Dogs Naturally Infected with Babesia canis canis. BMC Vet. Res. 2017, 13, 114. [Google Scholar] [CrossRef]
- Costantini, D.; Verhulst, S. Does High Antioxidant Capacity Indicate Low Oxidative Stress? Funct. Ecol. 2009, 23, 506–509. [Google Scholar] [CrossRef]
- Naraoka, M.; Li, Y.; Katagai, T.; Ohkuma, H. Effects of Apple Polyphenols on Oxidative Stress and Cerebral Vasospasm after Subarachnoid Hemorrhage in a Rabbit Double Hemorrhage Model. Brain Hemorrhages 2020, 1, 54–58. [Google Scholar] [CrossRef]
- Edwards, K.L.; Miller, M.A.; Carlstead, K.; Brown, J.L. Relationships between Housing and Management Factors and Clinical Health Events in Elephants in North American Zoos. PLoS ONE 2019, 14, e0217774. [Google Scholar] [CrossRef]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The Use of Leukocyte Profiles to Measure Stress in Vertebrates: A Review for Ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Drescher, B.; Bai, F. Neutrophil in Viral Infections, Friend or Foe? Virus Res. 2013, 171, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, N.; Sahu, S.K.; Das, A.K.; Mohapatra, D.; Panda, S.K.; Gupta, S.K.; Behera, B.K.; Pahari, A.; Dash, M. Elephant Endotheliotropic Herpesvirus Hemorrhagic Disease Outbreak in an Indian Zoo. J. Zoo Wildl. Med. 2021, 52, 1286–1297. [Google Scholar] [CrossRef]
- Iyer, M.L.; Molter, C.M.; Flanagan, J.P.; Bauer, K.L.; Bernardy, R.; Hoffman, D.; Parkinson, L.; Brainard, B.M.; Evans, T.S.; Pursell, T.; et al. Novel Diagnostic and Therapeutic Approaches to Elephant Endotheliotroic Herpesvirus 1A Hemorrhagic Disease in a Captive Juvenile Asian Elephant (Elephas maximus). J. Zoo Wildl. Med. 2022, 53, 232–240. [Google Scholar] [CrossRef]
- Ibrahim, H.M.M. Oxidative Stress Associated with Spasmodic, Flatulent, and Impaction Colic in Draft Horses. J. Equine Vet. Sci. 2014, 34, 1205–1210. [Google Scholar] [CrossRef]
- El-Ashker, M.; El-Khodery, S.; Metwally, N.; Hussein, H.; El-Boshy, M. Prognostic Significance of Oxidative Stress Markers in Colitis Associated with Phenylbutazone Administration in Draft Horses. J. Equine Vet. Sci. 2012, 32, 146–152. [Google Scholar] [CrossRef]
- Merl, S.; Scherzer, S.; Palme, R.; Möstl, E. Pain Causes Increased Concentrations of Glucocorticoid Metabolites in Horse Feces. J. Equine Vet. Sci. 2000, 20, 586–590. [Google Scholar] [CrossRef]
- Mair, T.S.; Sherlock, C.E.; Boden, L.A. Serum Cortisol Concentrations in Horses with Colic. Vet. J. 2014, 201, 370–377. [Google Scholar] [CrossRef]
- Chiaradia, E.; Avellini, L.; Rueca, F.; Spaterna, A.; Porciello, F.; Antonioni, M.T.; Gaiti, A. Physical Exercise, Oxidative Stress and Muscle Damage in Racehorses. Comp. Biochem. Physiol. B Biochem. 1998, 119, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Prasad, G.; Dhillon, M.S.; Khullar, M.; Nagi, O.N. Evaluation of Oxidative Stress after Fractures. A Preliminary Study. Acta Orthop. Belg. 2003, 69, 546–551. [Google Scholar] [PubMed]
- Sheweita, S.A.; Khoshhal, K.I. Calcium Metabolism and Oxidative Stress in Bone Fractures: Role of Antioxidants. Curr. Drug Metab. 2007, 8, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, M.A.; Hammouda, O.; Matran, R.; Robin, S.; Fabre, C. Changes in Oxidative Stress Markers and Biological Markers of Muscle Injury with Aging at Rest and in Response to an Exhaustive Exercise. PLoS ONE 2014, 9, e90420. [Google Scholar] [CrossRef]
- Buddi, R.; Lin, B.; Atilano, S.R.; Zorapapel, N.C.; Kenney, M.C.; Brown, D.J. Evidence of Oxidative Stress in Human Corneal Diseases. J. Histochem. Cytochem. 2002, 50, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Shoham, A.; Hadziahmetovic, M.; Dunaief, J.L.; Mydlarski, M.B.; Schipper, H.M. Oxidative Stress in Diseases of the Human Cornea. Free Radic. Biol. Med. 2008, 45, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Chaurasia, S.; Roy, S. Oxidative Stress in the Pathogenesis of Corneal Endothelial Dystrophies and Other Corneal Diseases. React. Oxyg. Species 2018, 6, 299–310. [Google Scholar] [CrossRef]
- Kraiwong, N.; Sanyathitiseree, P.; Boonprasert, K.; Diskul, P.; Charoenphan, P.; Pintawong, W.; Thayananuphat, A. Anterior Ocular Abnormalities of Captive Asian Elephants (Elephas maximus indicus) in Thailand. Vet. Ophthalmol. 2016, 19, 269–274. [Google Scholar] [CrossRef]
- Moseley, R.; Hilton, J.R.; Waddington, R.J.; Harding, K.G.; Stephens, P.; Thomas, D.W. Comparison of Oxidative Stress Biomarker Profiles between Acute and Chronic Wound Environments. Wound Repair Regen. 2004, 12, 419–429. [Google Scholar] [CrossRef]
- Firth, C.A.; Laing, A.D.; Baird, S.K.; Pearson, J.; Gieseg, S.P. Inflammatory Sites as a Source of Plasma Neopterin: Measurement of High Levels of Neopterin and Markers of Oxidative Stress in Pus Drained from Human Abscesses. Clin. Biochem. 2008, 41, 1078–1083. [Google Scholar] [CrossRef]
- Fujii, T.; Tokuda, S.; Nakazawa, Y.; Kurozumi, S.; Obayashi, S.; Yajima, R.; Shirabe, K. Implications of Low Serum Albumin as a Prognostic Factor of Long-Term Outcomes in Patients with Breast Cancer. In Vivo 2020, 34, 2033–2036. [Google Scholar] [CrossRef]
- Tussing-Humphreys, L.; Pustacioglu, C.; Nemeth, E.; Braunschweig, C. Rethinking Iron Regulation and Assessment in Iron Deficiency, Anemia of Chronic Disease, and Obesity: Introducing Hepcidin. J. Acad. Nutr. Diet. 2012, 112, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Liczner, G.; Cadelski, K.; Kołnierzak, T.; Nowaczewska, M.; Malinowski, B. Anemia of Chronic Diseases: Wider Diagnostics—Better Treatment? Nutrients 2020, 12, 1784. [Google Scholar] [CrossRef] [PubMed]
- Washington, I.M.; Van Hoosier, G. Clinical Biochemistry and Hematology. In The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Elsevier: Amsterdam, The Netherlands, 2012; pp. 57–116. ISBN 978-0-12-380920-9. [Google Scholar]
- Greene, W.; Mikota, S.; Pitcairn, J.; Ryer, M. Clinical Management of a Complete Gastrointestinal Obstruction and Ileus in a Geriatric Female Asian Elephant (Elephas maximus). J. Zoo Biol. 2019, 2, 01–04. [Google Scholar] [CrossRef]
- Palanivelrajan, M.; Prathipa, A.; Senthilkumar, A.; Jayathangaraj, M.G. Therapeutic Management of Arthritis in a Temple Elephant (Elephas maximus). J. Wildl. Res. 2015, 3, 19–20. [Google Scholar]
- Nath, I.; Samantara, S.; Cheeran, J.V.; Dangolla, A.; Panda, S.K. Surgical Management of Temporal Bursitis in a Captive Asian Elephant. Gajah 2011, 42. [Google Scholar]
- Purnama, M.T.E.; Hendrawan, D.; Wicaksono, A.P.; Fikri, F.; Purnomo, A.; Chhetri, S. Risk Factors, Hematological and Biochemical Profile Associated with Colic in Delman Horses in Gresik, Indonesia. F1000Research 2021, 10, 950. [Google Scholar] [CrossRef]
- Olaogun, S.C.; Oyetoyinbo, T.E. Lameness and Its Associated Hematological Features among Nigerian Breeds of Goats in Ibadan, Nigeria. Niger. J. Anim. Sci. 2020, 22, 315–324. [Google Scholar]
- Katica, M.; Šeho-Alić, A.; Čelebičić, M.; Prašović, S.; Alić, A. Histopathological and Hematological Changes Caused by Head Abscess in Rat. Veterinaria 2019, 68, 151–156. [Google Scholar]
- Preena, P.; Aravind, B. Case Report on the Medical Management of Corneal Opacity in a Captive Elephant Tusker. Adv. Anim. Vet. Sci. 2017, 8, 342–345. [Google Scholar]
- Lassaline, M.E.; Brooks, D.E.; Ollivier, F.J.; Komaromy, A.M.; Kallberg, M.E.; Gelatt, K.N. Equine Amniotic Membrane Transplantation for Corneal Ulceration and Keratomalacia in Three Horses. Vet. Ophthalmol. 2005, 8, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Lores, M.; Rakestraw, P.; De Rijck, M.; Yarbrough, T. Use of Autologous Fascia Lata Graft to Repair a Complex Corneal Ulcer in a Mare. Ir. Vet. J. 2020, 73, 7. [Google Scholar] [CrossRef]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal 2015, 23, 1144–1170. [Google Scholar] [CrossRef]
- Singh, S.; Verma, S.K.; Kumar, S.; Ahmad, M.K.; Nischal, A.; Singh, S.K.; Dixit, R.K. Evaluation of Oxidative Stress and Antioxidant Status in Chronic Obstructive Pulmonary Disease. Scand. J. Immunol. 2017, 85, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, E.; Youssef, C.; Rita, B.; Kaoutar, B.; Islah, L.; Najia, E.; Abdarrahmane, B.; Bernard, F. Impact of Transport Distance on Stress Biomarkers Levels in Dromedary Camel (Camelus dromedarius). Emir. J. Food Agric 2015, 27, 507. [Google Scholar] [CrossRef]
- Myint, K.; Jayakumar, R.; Hoe, S.-Z.; Lam, S.-K. Cortisol, β-Endorphin and Oxidative Stress Markers in Healthy Medical Students in Response to Examination Stress. Biomed. Res. 2017, 28, 6. [Google Scholar]
- Mohamed, R.H.; Khalphallah, A.; Nakada, K.; Elmeligy, E.; Hassan, D.; Ebissy, E.A.; Ghandour, R.A.; Mousa, S.A.; Hassaneen, A.S.A. Clinical and Correlated Responses among Steroid Hormones and Oxidant/Antioxidant Biomarkers in Pregnant, Non-Pregnant and Lactating CIDR-Pre-Synchronized Dromedaries (Camelus dromedarius). Vet. Sci. 2021, 8, 247. [Google Scholar] [CrossRef]
- Asker, S.; Asker, M.; Sarikaya, E.; Sunnetcioglu, A.; Aslan, M.; Demir, H. Oxidative Stress Parameters and Their Correlation with Clinical, Metabolic and Polysomnographic Parameters in Severe Obstructive Sleep Apnea Syndrome. Int. J. Clin. Exp. Med. 2015, 8, 11449–11455. [Google Scholar]
- D’Alessandro, A.; Fu, X.; Reisz, J.A.; Stone, M.; Kleinman, S.; Zimring, J.C.; Busch, M.; for the Recipient Epidemiology and Donor Evaluation Study-III (REDS III). Ethyl Glucuronide, a Marker of Alcohol Consumption, Correlates with Metabolic Markers of Oxidant Stress but Not with Hemolysis in Stored Red Blood Cells from Healthy Blood Donors. Transfusion 2020, 60, 1183–1196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Area | Location | Number of Elephants | Number of Camps | Elephant Activities | Main Diet |
|---|---|---|---|---|---|
| North | Chiang Mai | 104 | 20 | Elephant trekking, observation, tourist feeding, bathing, playing in mud, elephant show | Fresh grasses, corn stalks |
| North | Lampang | 16 | 1 | Elephant trekking, observation, tourist feeding, bathing, elephant show | Fresh grasses, corn stalks |
| West | Kanchanaburi | 4 | 2 | Elephant trekking, tourist feeding, elephant show, bathing | Pineapple leaves, fresh grasses |
| Center | Phra Nakorn Si Ayutthaya | 3 | 1 | Elephant trekking, tourist feeding, elephant show | Pineapple leaves, fresh grasses |
| East | Surin | 2 | 2 | Tourist feeding, elephant show | Fresh grasses, corn stalks |
| East | Chonburi | 8 | 1 | Elephant trekking, tourist feeding, elephant show | Pineapple leaves, fresh grasses |
| Biomarkers | Range | Overall | Age Group | Sex | Season | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Juvenile 1–5 Years | Subadult 6–10 Years | Adult 11–45 Years | Aged >46 Years | Male | Female | Summer | Rainy | Winter | |||
| ROS (mg H2O2/L) | 1.91–2.47 | 2.18 ± 0.16 (n = 137) | 2.13 ± 0.17 (n = 45) | 2.18 ± 0.17 (n = 36) | 2.22 ± 0.13 (n = 45) | 2.24 ± 0.09 (n = 11) | 2.16 ± 0.17 (n = 45) | 2.19 ± 0.15 (n = 92) | 2.29 ± 0.13 (n = 33) | 2.11 ± 0.17 (n = 53) | 2.19 ± 0.11 (n = 51) |
| MDA (nmol/mL) | 0.93–2.49 | 1.76 ± 0.47 (n = 137) | 1.77 ± 0.49 (n = 45) | 1.98 ± 0.48 (n = 36) | 1.59 ± 0.43 (n = 45) | 1.63 ± 0.19 (n = 11) | 1.89 ± 0.51 (n = 45) | 1.69 ± 0.45 (n = 92) | 1.89 ± 0.35 (n = 33) | 1.77 ± 0.49 (n = 53) | 1.65 ± 0.51 (n = 51) |
| Serum albumin (g/dL) | 2.73–3.96 | 3.33 ± 0.36 (n = 136) | 3.22 ± 0.37 (n = 45) | 3.45 ± 0.31 (n = 36) | 3.29 ± 0.36 (n = 44) | 3.54 ± 0.32 (n = 11) | 3.40 ± 0.33 (n = 45) | 3.29 ± 0.37 (n = 91) | 3.35 ± 0.38 (n = 33) | 3.32 ± 0.34 (n = 53) | 3.32 ± 0.36 (n = 50) |
| GPx (U/L) | 0.14–2.08 | 1.16 ± 0.56 (n = 136) | 1.16 ± 0.53 (n = 45) | 1.28 ± 0.62 (n = 36) | 1.08 ± 0.53 (n = 44) | 1.03 ± 0.53 (n = 11) | 1.09 ± 0.51 (n = 45) | 1.19 ± 0.58 (n = 91) | 1.29 ± 0.56 (n = 33) | 1.25 ± 0.57 (n = 53) | 0.97 ± 0.49 (n = 50) |
| Catalase (U/mL) | 2.99–21.86 | 13.05 ± 5.54 (n = 137) | 11.71 ± 4.31 (n = 45) | 13.68 ± 5.87 (n = 36) | 13.95 ± 6.35 (n = 45) | 12.72 ± 4.95 (n = 11) | 13.3 ± 6.05 (n = 45) | 12.9 ± 5.30 (n = 92) | 8.96 ± 3.96 (n = 33) | 12.9 ± 4.99 (n = 53) | 15.80 ± 5.35 (n = 51) |
| fGCMs (ng/g feces) | 22.23–82.70 | 51.97 ± 17.62 (n = 85) | 52.13 ± 17.10 (n = 17) | 55.59 ± 14.68 (n = 16) | 51.12 ± 17.02 (n = 41) | 49.61 ± 25.08 (n = 11) | 51.67 ± 15.48 (n = 22) | 52.07 ± 18.43 (n = 63) | 44.58 ± 13.88 (n = 21) | 58.08 ± 17.42 (n = 16) | 53.16 ± 18.37 (n = 48) |
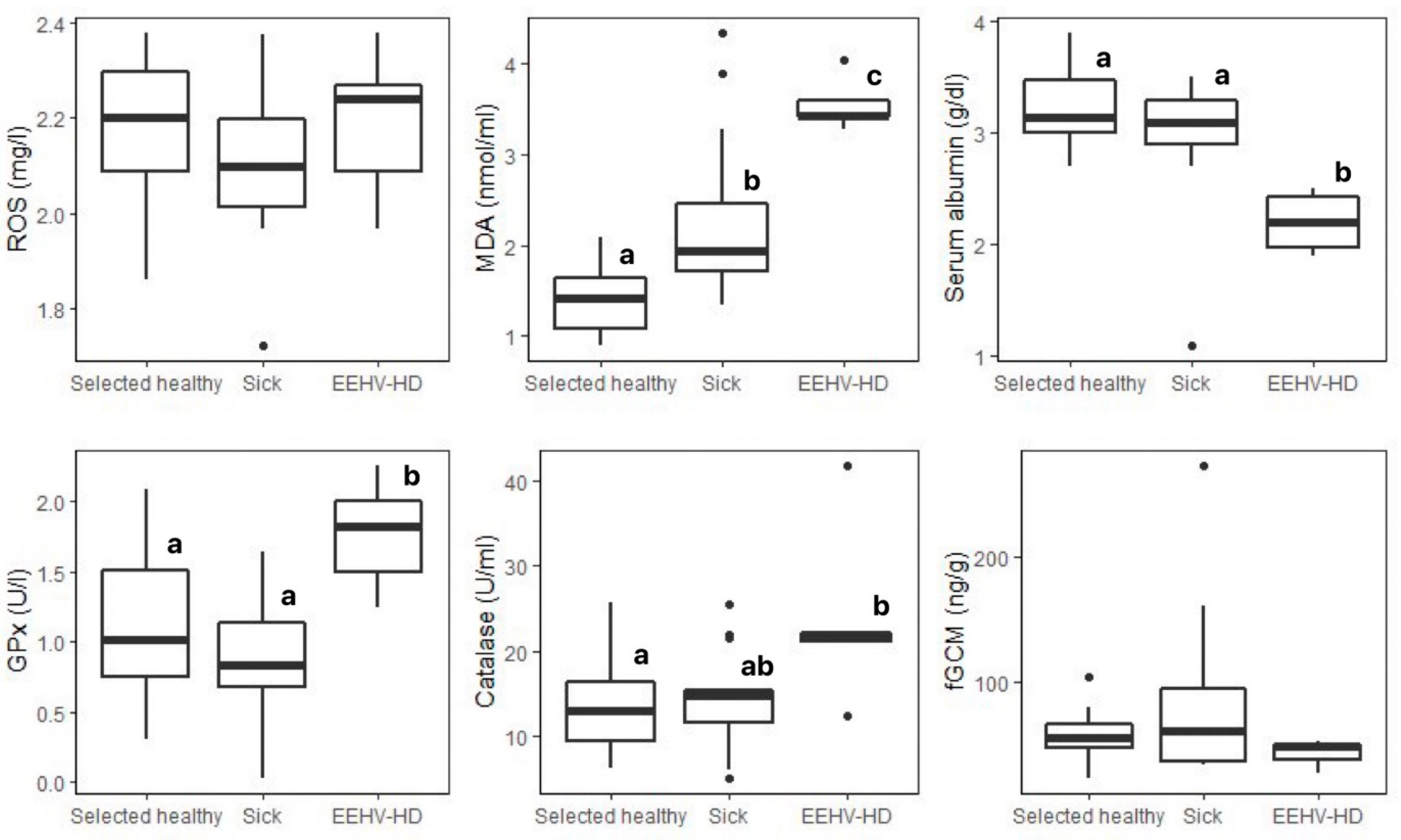
| Biomarkers | Selected Healthy (n = 30) 1 | Sick 2 (n = 15) | EEHV-HD (n = 5) | H Statistic | p-Value |
|---|---|---|---|---|---|
| ROS (mg/L) | 2.18 ± 0.14 (1.86–2.38) | 2.10 ± 0.16 (1.72–2.37) | 2.19 ± 0.16 (1.97–2.38) | 2.637 | 0.267 |
| MDA (nmol/mL) | 1.43 ± 0.35 a (0.90–2.49) | 2.26 ± 0.91 b (1.34–4.33) | 3.55 ± 0.29 c (3.28–4.03) | 22.972 | <0.01 ** |
| Albumin (g/dL) | 3.23 ± 0.33 a (2.70–3.90) | 3.00 ± 0.62 a (1.10–3.50) | 2.20 ± 0.29 b (1.90–2.50) | 10.643 | <0.01 ** |
| GPx (U/L) | 1.13 ± 0.47 a (0.30–2.09) | 0.89 ± 0.39 a (0.30–1.64) | 1.76 ± 0.40 b (1.24–2.25) | 9.729 | <0.01 ** |
| Catalase (U/mL) | 13.25 ± 4.85 a (6.33–25.64) | 14.35 ± 5.56 ab (5.13–25.51) | 23.81 ± 10.76 b (12.52–41.78) | 6.083 | 0.048 * |
| fGCMs (ng/g) | 55.43 ± 19.01 (22.15–103.23) | 88.14 ± 77.05 (32.30–273.68) | 42.00 ± 13.63 (26.48–52.03) | 2.222 | 0.329 |
| Parameter | Unit | Range (Healthy) | Health Status | |||
|---|---|---|---|---|---|---|
| Healthy (n = 137) | Sick 1 (n = 15) | EEHV-HD (n = 5) | ||||
| RBCs | PCV | % | 27.62–47.52 | 37.51 ± 5.97 | 32.82 ± 2.29 | 35.80 ± 7.56 |
| Hemoglobin | g/dL | 10.29–16.10 | 13.31 ± 1.89 | 11.87 ± 0.78 | 12.34 ± 3.06 | |
| RBC count | ×106 cells/µL | 2.39–3.91 | 3.15 ± 0.45 | 2.70 ± 0.20 | 3.24 ± 0.65 | |
| MCV | fl | 106.30–129.08 | 117.72 ± 6.68 | 121.85 ± 3.65 | 110.98 ± 3.43 | |
| MCHCs | g/dL | 34.62–36.82 | 35.80 ± 0.73 | 36.16 ± 0.89 | 34.20 ± 2.83 | |
| WBCs | WBC count | cells/µL | 7130.58–20,368.19 | 13,972.19 ± 4040.26 | 12,467.50 ± 4457.69 | 18,950.00 ± 11,131.23 |
| Heterophil | cells/µL | 974.61–5383.52 | 3506.49 ± 1596.61 | 3672.50 ± 1942.69 | 9502.80 ± 4755.95 | |
| Lymphocyte | cells/µL | 686.23–9985.99 | 5529.07 ± 2590.80 | 4029.42 ± 1642.62 | 5909.40 ± 2778.33 | |
| Monocyte | cells/µL | 422.91–7459.65 | 4173.14 ± 2066.36 | 4112.42 ± 1833.32 | 3351.40 ± 4078.38 | |
| Eosinophil | cells/µL | 0–598.60 | 268.08 ± 267.25 | 626.75 ± 1331.35 | 186.40 ± 215.53 | |
| Basophil | cells/µL | 0–100.33 | 33.10 ± 62.36 | 24.83 ± 86.02 | Not found | |
| H:L ratio | >0–1.47 | 0.79 ± 0.63 | 1.07 ± 0.63 | 1.59 ± 0.26 | ||
| M:H ratio | >0–2.73 | 1.39 ± 0.88 | 1.31 ± 0.64 | 0.30 ± 0.24 | ||
| Platelets | Platelet count | ×103 cells/µL | 151.08–552.62 | 361.38 ± 120.19 | 400.25 ± 65.50 | 118.40 ± 103.71 |
| Blood chemistry | BUN | mg/dL | 4.44–15.31 | 10.25 ± 3.42 | 8.67 ± 2.80 | 12.80 ± 2.00 |
| Creatinine | mg/dL | 0.90–1.89 | 1.42 ± 0.29 | 1.34 ± 0.39 | 2.20 ± 0.33 | |
| ALT | U/L | 0–2.11 | 0.92 ± 1.03 | 4.15 ± 9.92 | 5.75 ± 3.59 | |
| ALP | U/L | 0–306.92 | 154.59 ± 93.68 | 100.80 ± 73.26 | 113.00 ± 28.08 | |
| Total serum protein | g/dL | 6.81–9.91 | 8.33 ± 0.85 | 8.10 ± 0.82 | 5.57 ± 0.76 | |
| Parameters | ROS | MDA | Albumin | GPx | Catalase | fGCMs | RBCs | WBCs | HETs | LYMs | MONOs | PLTs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROS | ||||||||||||
| MDA | −0.024 | |||||||||||
| Albumin | 0.09 | 0.26 ** | ||||||||||
| GPx | 0.15 | 0.29 ** | 0.02 | |||||||||
| Catalase | −0.09 | 0.15 | 0.18 | 0.17 | ||||||||
| fGCMs | −0.36 ** | −0.15 | −0.08 | −0.08 | 0.23 * | |||||||
| RBCs | 0.27 ** | 0.27 ** | 0.23 ** | 0.22 * | 0.18 * | −0.33 ** | ||||||
| WBCs | −0.01 | 0.06 | −0.03 | 0.16 | 0.11 | 0.04 | −0.01 | |||||
| HETs | −0.11 | −0.14 | −0.18 * | 0.05 | 0.01 | 0.17 | −0.25 ** | 0.57 ** | ||||
| LYMs | 0.11 | 0.30 ** | 0.19 * | 0.10 | 0.17 | −0.11 | 0.19 * | 0.58 ** | 0.01 | |||
| MONOs | 0.10 | −0.07 | −0.07 | 0.18 * | 0.07 | −0.21 | 0.04 | 0.46 ** | 0.05 | −0.06 | ||
| PLTs | −0.25 ** | −0.10 | −0.18 * | 0.10 | −0.04 | 0.30 ** | −0.23 ** | 0.63 ** | 0.52 ** | 0.11 | 0.44 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosaruk, W.; Brown, J.L.; Towiboon, P.; Punyapornwithaya, V.; Pringproa, K.; Thitaram, C. Measures of Oxidative Status Markers in Relation to Age, Sex, and Season in Sick and Healthy Captive Asian Elephants in Thailand. Animals 2023, 13, 1548. https://doi.org/10.3390/ani13091548
Kosaruk W, Brown JL, Towiboon P, Punyapornwithaya V, Pringproa K, Thitaram C. Measures of Oxidative Status Markers in Relation to Age, Sex, and Season in Sick and Healthy Captive Asian Elephants in Thailand. Animals. 2023; 13(9):1548. https://doi.org/10.3390/ani13091548
Chicago/Turabian StyleKosaruk, Worapong, Janine L. Brown, Patcharapa Towiboon, Veerasak Punyapornwithaya, Kidsadagon Pringproa, and Chatchote Thitaram. 2023. "Measures of Oxidative Status Markers in Relation to Age, Sex, and Season in Sick and Healthy Captive Asian Elephants in Thailand" Animals 13, no. 9: 1548. https://doi.org/10.3390/ani13091548