
Multimodal Communication in the Human–Cat Relationship: A Pilot Study

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
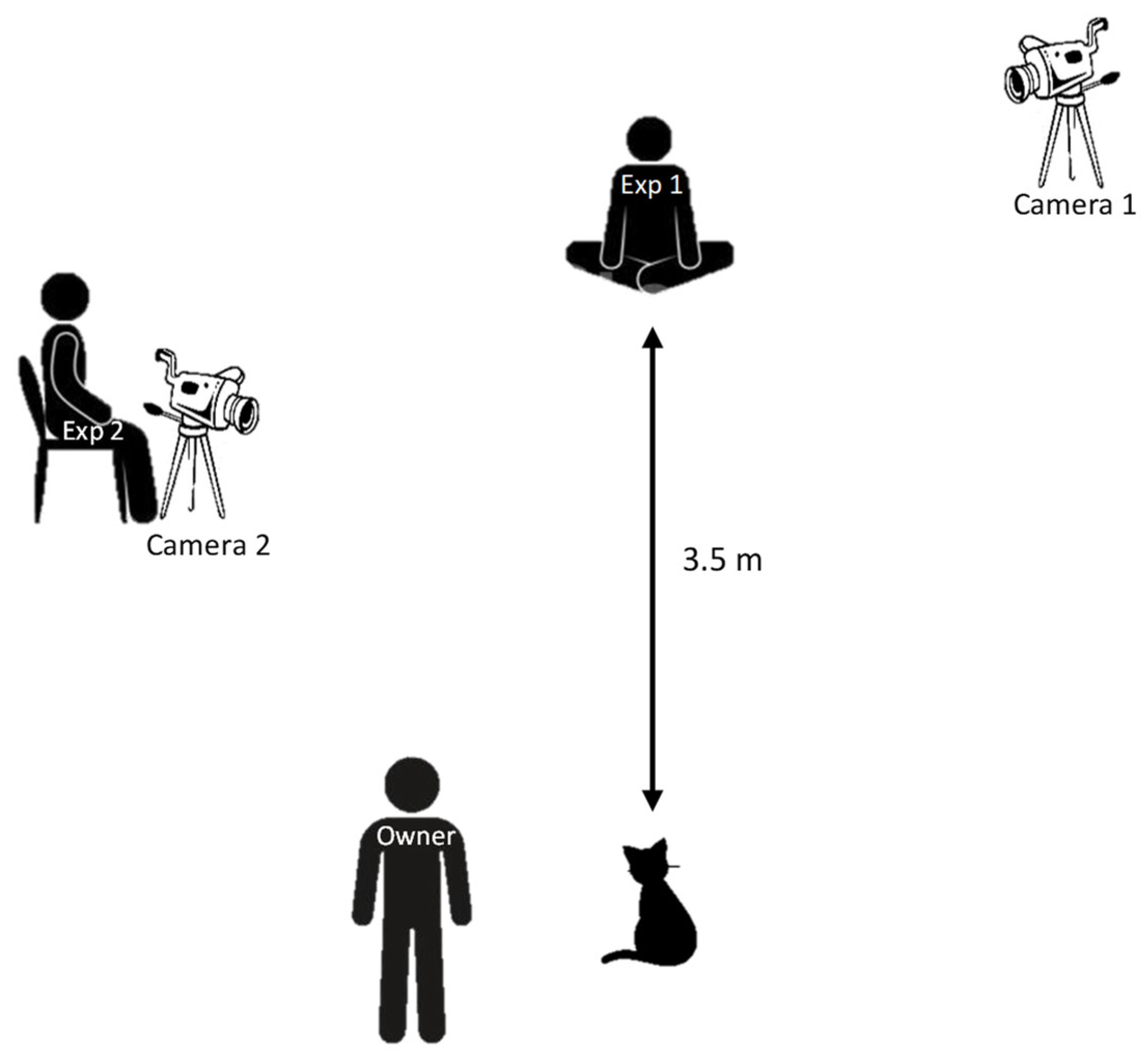
2.2. Apparatus
2.3. Procedure
2.3.1. Habituation Phase
2.3.2. Testing Conditions
2.4. Behavioural Analysis
2.5. Statistical Analysis
3. Result
3.1. Latency of Approach
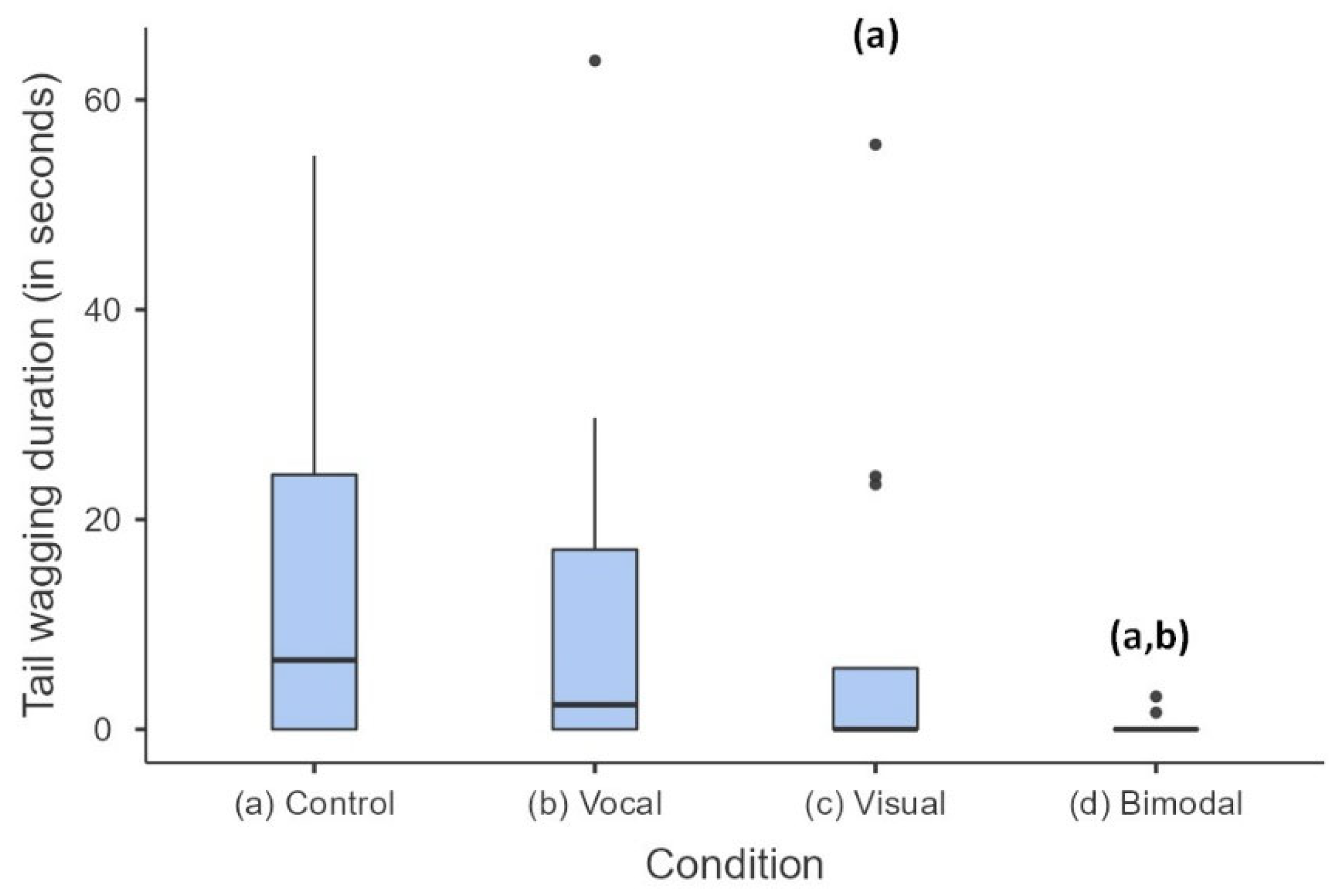
3.2. Other Behavioural Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humphrey, T.; Proops, L.; Forman, J.; Spooner, R.; McComb, K. The Role of Cat Eye Narrowing Movements in Cat–Human Communication. Sci. Rep. 2020, 10, 16503. [Google Scholar] [CrossRef] [PubMed]
- Miklósi, Á.; Pongrácz, P.; Lakatos, G.; Topál, J.; Csányi, V. A Comparative Study of the Use of Visual Communicative Signals in Interactions Between Dogs (Canis familiaris) and Humans and Cats (Felis catus) and Humans. J. Comp. Psychol. 2005, 119, 179–186. [Google Scholar] [CrossRef]
- Albuquerque, N.; Guo, K.; Wilkinson, A.; Savalli, C.; Otta, E.; Mills, D. Dogs Recognize Dog and Human Emotions. Biol. Lett. 2016, 12, 20150883. [Google Scholar] [CrossRef] [PubMed]
- Aria, M.; Alterisio, A.; Scandurra, A.; Pinelli, C.; D’Aniello, B. The Scholar’s Best Friend: Research Trends in Dog Cognitive and Behavioral Studies. Anim. Cogn. 2021, 24, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Cannas, S.; Mattiello, S.; Battini, M.; Ingraffia, S.I.; Cadoni, D.; Palestrini, C. Evaluation of Maine Coon cat behavior during three different management situations. J. Vet. Behav. 2020, 37, 93–100. [Google Scholar] [CrossRef]
- D’Aniello, B.; Scandurra, A.; Alterisio, A.; Valsecchi, P.; Prato-Previde, E. The importance of gestural communication: A study of human–dog communication using incongruent information. Anim. Cogn. 2016, 19, 1231–1235. [Google Scholar] [CrossRef]
- D’Aniello, B.; Fierro, B.; Scandurra, A.; Pinelli, C.; Aria, M.; Semin, G.R. Sex Differences in the Behavioral Responses of Dogs Exposed to Human Chemosignals of Fear and Happiness. Anim. Cogn. 2021, 24, 299–309. [Google Scholar] [CrossRef] [PubMed]
- De Mouzon, C.; Gonthier, M.; Leboucher, G. Discrimination of Cat-Directed Speech from Human-Directed Speech in a Population of Indoor Companion Cats (Felis catus). Anim. Cogn. 2023, 26, 611–619. [Google Scholar] [CrossRef]
- Ford, G.; Guo, K.; Mills, D. Human facial expression affects a dog’s response to conflicting directional gestural cues. Behav. Proc. 2019, 159, 80–85. [Google Scholar] [CrossRef]
- Galvan, M.; Vonk, J. Man’s Other Best Friend: Domestic Cats (F. silvestris catus) and Their Discrimination of Human Emotion Cues. Anim. Cogn. 2016, 19, 193–205. [Google Scholar] [CrossRef]
- Gergely, A.; Petró, E.; Oláh, K.; Topál, J. Auditory–Visual Matching of Conspecifics and Non-Conspecifics by Dogs and Human Infants. Animals 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.; Racca, A.; Scaf, B.; Virányi, Z.; Range, F. Discrimination of Familiar Human Faces in Dogs (Canis familiaris). Learn. Motiv. 2013, 44, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Watanabe, A.; Takagi, S.; Arahori, M.; Saito, A. Cats beg for food from the human who looks at and calls to them: Ability to understand humans’ attentional states. Psychologia 2016, 59, 112–120. [Google Scholar] [CrossRef]
- Pongrácz, P.; Szapu, J.S.; Faragó, T. Cats (Felis silvestris catus) Read Human Gaze for Referential Information. Intelligence 2019, 74, 43–52. [Google Scholar] [CrossRef]
- Prato-Previde, E.; Cannas, S.; Palestrini, C.; Ingraffia, S.; Battini, M.; Ludovico, L.A.; Ntalampiras, S.; Presti, G.; Mattiello, S. What’s in a Meow? A Study on Human Classification and Interpretation of Domestic Cat Vocalizations. Animals 2020, 10, 2390. [Google Scholar] [CrossRef] [PubMed]
- Quaranta, A.; d’Ingeo, S.; Amoruso, R.; Siniscalchi, M. Emotion Recognition in Cats. Animals 2020, 10, 1107. [Google Scholar] [CrossRef]
- Scandurra, A.; Alterisio, A.; Marinelli, L.; Mongillo, P.; Semin, G.R.; D’Aniello, B. Effectiveness of verbal and gestural signals and familiarity with signal-senders on the performance of working dogs. Appl. Anim. Behav. Sci. 2017, 191, 78–83. [Google Scholar] [CrossRef]
- Scandurra, A.; Pinelli, C.; Fierro, B.; Di Cosmo, A.; D’Aniello, B. Multimodal signaling in the visuo-acoustic mismatch paradigm: Similarities between dogs and children in the communicative approach. Anim. Cogn. 2020, 23, 833–841. [Google Scholar] [CrossRef]
- Takagi, S.; Arahori, M.; Chijiiwa, H.; Saito, A.; Kuroshima, H.; Fujita, K. Cats Match Voice and Face: Cross-Modal Representation of Humans in Cats (Felis catus). Anim. Cogn. 2019, 22, 901–906. [Google Scholar] [CrossRef]
- Vitale, K.R.; Udell, M.A.R. The Quality of Being Sociable: The Influence of Human Attentional State, Population, and Human Familiarity on Domestic Cat Sociability. Behav. Proc. 2019, 158, 11–17. [Google Scholar] [CrossRef]
- Gácsi, M.; McGreevy, P.; Kara, E.; Miklósi, Á. Effects of Selection for Cooperation and Attention in Dogs. Behav. Brain Funct. 2009, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.; Tomasello, M. Human-like Social Skills in Dogs? Trends Cogn. Sci. 2005, 9, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Adachi, I.; Kuwahata, H.; Fujita, K. Dogs Recall Their Owner’s Face upon Hearing the Owner’s Voice. Anim. Cogn. 2007, 10, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Call, J.; Bräuer, J.; Kaminski, J.; Tomasello, M. Domestic Dogs (Canis familiaris) Are Sensitive to the Attentional State of Humans. J. Comp. Psychol. 2003, 117, 257–263. [Google Scholar] [CrossRef]
- Gácsi, M.; Miklósi, Á.; Varga, O.; Topál, J.; Csányi, V. Are Readers of Our Face Readers of Our Minds? Dogs (Canis familiaris) Show Situation-Dependent Recognition of Human’s Attention. Anim. Cogn. 2004, 7, 144–153. [Google Scholar] [CrossRef]
- Miklösi, Á.; Polgárdi, R.; Topál, J.; Csányi, V. Use of Experimenter-given Cues in Dogs. Anim. Cogn. 1998, 1, 113–121. [Google Scholar] [CrossRef]
- Racca, A.; Amadei, E.; Ligout, S.; Guo, K.; Meints, K.; Mills, D. Discrimination of Human and Dog Faces and Inversion Responses in Domestic Dogs (Canis familiaris). Anim. Cogn. 2010, 13, 525–533. [Google Scholar] [CrossRef]
- Schwab, C.; Huber, L. Obey or Not Obey? Dogs (Canis familiaris) Behave Differently in Response to Attentional States of Their Owners. J. Comp. Psychol. 2006, 120, 169–175. [Google Scholar] [CrossRef]
- Soproni, K.; Miklósi, A.; Topál, J.; Csányi, V. Comprehension of Human Communicative Signs in Pet Dogs (Canis familiaris). J. Comp. Psychol. 2001, 115, 122–126. [Google Scholar] [CrossRef]
- Ben-Aderet, T.; Gallego-Abenza, M.; Reby, D.; Mathevon, N. Dog-Directed Speech: Why Do We Use It and Do Dogs Pay Attention to It? Proc. R. Soc. B 2017, 284, 20162429. [Google Scholar] [CrossRef]
- Benjamin, A.; Slocombe, K. ‘Who’s a Good Boy?!’ Dogs Prefer Naturalistic Dog-Directed Speech. Anim. Cogn. 2018, 21, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, S.; Gilbert, C.; Amy, M.; Leboucher, G. Pet-Directed Speech Draws Adult Dogs’ Attention More Efficiently than Adult-Directed Speech. Sci. Rep. 2017, 7, 4980. [Google Scholar] [CrossRef]
- Merola, I.; Lazzaroni, M.; Marshall-Pescini, S.; Prato-Previde, E. Social Referencing and Cat–Human Communication. Anim. Cogn. 2015, 18, 639–648. [Google Scholar] [CrossRef]
- Saito, A.; Shinozuka, K. Vocal Recognition of Owners by Domestic Cats (Felis catus). Anim. Cogn. 2013, 16, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Chijiiwa, H.; Arahori, M.; Saito, A.; Fujita, K.; Kuroshima, H. Socio-Spatial Cognition in Cats: Mentally Mapping Owner’s Location from Voice. PLoS ONE 2021, 16, e0257611. [Google Scholar] [CrossRef]
- Trösch, M.; Cuzol, F.; Parias, C.; Calandreau, L.; Nowak, R.; Lansade, L. Horses Categorize Human Emotions Cross-Modally Based on Facial Expression and Non-Verbal Vocalizations. Animals 2019, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- Beauchaud, M. La Communication Animale. In Ethologie Animale. Une Approche Biologique du Comportement, 2nd ed.; Darmaillacq, A.S., Lévy, F., Eds.; De Boeck Supérieur: Louvain-la-Neuve, France, 2020; pp. 127–148. [Google Scholar]
- Partan, S.; Marler, P. Communication Goes Multimodal. Science 1999, 283, 1272–1273. [Google Scholar] [CrossRef]
- Baraud, I.; Deputte, B.L.; Pierre, J.-S.; Blois-Heulin, C. Informative Value of Vocalizations during Multimodal Interactions in Red-Capped Mangabeys. In Discovering Hidden Temporal Patterns in Behavior and Interaction; Magnusson, M.S., Burgoon, J.K., Casarrubea, M., Eds.; Neuromethods; Springer: New York, NY, USA, 2016; Volume 111, pp. 255–277. ISBN 978-1-4939-3248-1. [Google Scholar]
- Turner, D.C. The Mechanics of Social Interactions Between Cats and Their Owners. Front. Vet. Sci. 2021, 8, 650143. [Google Scholar] [CrossRef]
- Cafazzo, S.; Natoli, E. The Social Function of Tail up in the Domestic Cat (Felis silvestris catus). Behav. Proc. 2009, 80, 60–66. [Google Scholar] [CrossRef]
- Cameron-Beaumont, C. Visual and Tactile Communication in the Domestic Cat (Felis silvestris catus) and Undomesticated Small Felids. Ph.D. Thesis, University of Southampton, Southampton, UK, 1997. [Google Scholar]
- Crowell-Davis, S. Cat Behaviour: Social Organization, Communication and Development. In The Welfare of Cats; Rochlitz, I., Ed.; Animal Welfare; Springer: Dordrecht, The Netherlands, 2007; Volume 3. [Google Scholar] [CrossRef]
- Deputte, B.L.; Jumelet, E.; Gilbert, C.; Titeux, E. Heads and Tails: An Analysis of Visual Signals in Cats, Felis catus. Animals 2021, 11, 2752. [Google Scholar] [CrossRef]
- Bradshaw, J.; Cameron-Beaumont, C. The Signalling Repertoire of the Domestic Cat and Its Undomesticated Relatives. In The Domestic Cat: The Biology of Its Behaviour, 2nd ed; Turner, D., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 67–93. [Google Scholar]
- Yeon, S.C.; Kim, Y.K.; Park, S.J.; Lee, S.S.; Lee, S.Y.; Suh, E.H.; Houpt, K.A.; Chang, H.H.; Lee, H.C.; Yang, B.G.; et al. Differences between Vocalization Evoked by Social Stimuli in Feral Cats and House Cats. Behav. Proc. 2011, 87, 183–189. [Google Scholar] [CrossRef]
- Nibblett, B.M.; Ketzis, J.K.; Grigg, E.K. Comparison of Stress Exhibited by Cats Examined in a Clinic versus a Home Setting. Appl. Anim. Behav. Sci. 2015, 173, 68–75. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A Free, Versatile Open-source Event-logging Software for Video/Audio Coding and Live Observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Stanton, L.A.; Sullivan, M.S.; Fazio, J.M. A Standardized Ethogram for the Felidae: A Tool for Behavioral Researchers. Appl. Anim. Behav. Sci. 2015, 173, 3–16. [Google Scholar] [CrossRef]
- The Jamovi Project, Jamovi Version 2.2. 2021. Available online: https://www.jamovi.org (accessed on 8 March 2022).
- Pereira, D.G.; Afonso, A.; Melo Medeiros, F. Overview of Friedman’s test and post-hoc analysis. Commun. Stat. -Simul. Comput. 2015, 44, 2636–2653. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Version 4.0; R Core Team: Vienna, Austria, 2021; Available online: https://cran.r-project.org (accessed on 8 March 2022).
- Pohlert, T. PMCMR: Calculate Pairwise Multiple Comparisons of Mean Rank Sums. [R Package]. 2018. Available online: https://cran.r-project.org/package=PMCMR (accessed on 8 March 2022).
- Ratuski, A.S.; Makowska, I.J.; Dvorack, K.R.; Weary, D.M. Using Approach Latency and Anticipatory Behaviour to Assess Whether Voluntary Playpen Access Is Rewarding to Laboratory Mice. Sci. Rep. 2021, 11, 18683. [Google Scholar] [CrossRef] [PubMed]
- Bensoussan, S.; Tigeot, R.; Lemasson, A.; Meunier-Salaün, M.-C.; Tallet, C. Domestic Piglets (Sus scrofa domestica) Are Attentive to Human Voice and Able to Discriminate Some Prosodic Features. Appl. Anim. Behav. Sci. 2019, 210, 38–45. [Google Scholar] [CrossRef]
- Mertens, C.; Turner, D.C. Experimental Analysis of Human-Cat Interactions During First Encounters. Anthrozoös 1988, 2, 83–97. [Google Scholar] [CrossRef]
- Uccheddu, S.; Miklósi, Á.; Gintner, S.; Gácsi, M. Comparing Pears to Apples: Unlike Dogs, Cats Need Habituation before Lab Tests. Animals 2022, 12, 3046. [Google Scholar] [CrossRef]
- Kiley-Worthington, M. The Tail Movements of Ungulates, Canids and Felids With Particular Reference To Their Causation and Function as Displays. Behaviour 1976, 56, 69–114. [Google Scholar] [CrossRef]
- Ellis, S.L.H.; Swindell, V.; Burman, O.H.P. Human Classification of Context-Related Vocalizations Emitted by Familiar And Unfamiliar Domestic Cats: An Exploratory Study. Anthrozoös 2015, 28, 625–634. [Google Scholar] [CrossRef]
- Bouma, E.M.C.; Reijgwart, M.L.; Dijkstra, A. Family Member, Best Friend, Child or ‘Just’ a Pet, Owners’ Relationship Perceptions and Consequences for Their Cats. IJERPH 2021, 19, 193. [Google Scholar] [CrossRef]
- Eriksson, M.; Keeling, L.J.; Rehn, T. Cats and Owners Interact More with Each Other after a Longer Duration of Separation. PLoS ONE 2017, 12, e0185599. [Google Scholar] [CrossRef] [PubMed]
- Finka, L.R.; Ward, J.; Farnworth, M.J.; Mills, D.S. Owner Personality and the Wellbeing of Their Cats Share Parallels with the Parent-Child Relationship. PLoS ONE 2019, 14, e0211862. [Google Scholar] [CrossRef] [PubMed]
- Vitale, K.R.; Behnke, A.C.; Udell, M.A.R. Attachment Bonds between Domestic Cats and Humans. Curr. Biol. 2019, 29, R864–R865. [Google Scholar] [CrossRef]
- Vitale Shreve, K.R.; Mehrkam, L.R.; Udell, M.A.R. Social Interaction, Food, Scent or Toys? A Formal Assessment of Domestic Pet and Shelter Cat (Felis silvestris catus) Preferences. Behav. Proc. 2017, 141, 322–328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behaviour | Description | Information |
|---|---|---|
| Vocalizing | Cat is producing sounds originating from the throat and mouth | Vocal signal, emotional state |
| Blinking | Cat blinking (eyes narrowing) | Visual signal, emotional state |
| Looking away | Cat is avoiding eye contact by looking elsewhere | Visual signal, emotional state |
| Sniffing | Cat is smelling the ground or any object | Visual signal, emotional state |
| Grooming | Cat is quickly licking breastplate, flank or genitals | Emotional state |
| Locomotion | Cat is moving around the room (more than one-step displacements) | Exploratory behaviour |
| Ear(s) moving | Ear movement in any direction | Sensitivity to signal |
| Experimenter | Cat is looking towards experimenter 1 | Sensitivity to signal |
| Owner | Cat is looking towards owner | Emotional state |
| Tail wagging | Tail is slowly moving from side to side, in a continuous lateral movement | Emotional state |
| Tail up | Tail is held in an upright position | Visual signal, emotional state |
| Behaviour | χ2 | df | p |
|---|---|---|---|
| Vocalizing | 0.931 | 3 | 0.818 |
| Blinking | 6 | 3 | 0.112 |
| Looking away | 4.02 | 3 | 0.260 |
| Sniffing | 1.91 | 3 | 0.591 |
| Grooming | 5.00 | 3 | 0.172 |
| Locomotion | 4.16 | 3 | 0.245 |
| Ear moving | 4.79 | 3 | 0.188 |
| Experimenter | 5.12 | 3 | 0.163 |
| Owner | 4.23 | 3 | 0.237 |
| Tail wagging | 8.81 | 3 | 0.032 |
| Tail up | 1.05 | 3 | 0.788 |
| Latency for approaching | 10.8 | 3 | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mouzon, C.; Leboucher, G. Multimodal Communication in the Human–Cat Relationship: A Pilot Study. Animals 2023, 13, 1528. https://doi.org/10.3390/ani13091528
de Mouzon C, Leboucher G. Multimodal Communication in the Human–Cat Relationship: A Pilot Study. Animals. 2023; 13(9):1528. https://doi.org/10.3390/ani13091528
Chicago/Turabian Stylede Mouzon, Charlotte, and Gérard Leboucher. 2023. "Multimodal Communication in the Human–Cat Relationship: A Pilot Study" Animals 13, no. 9: 1528. https://doi.org/10.3390/ani13091528