
Influence of Seasonality and Culture Stage of Farmed Nile Tilapia (Oreochromis niloticus) with Monogenean Parasitic Infection
 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Fish Collection and Analysis (Biotic Factors)
2.2. Collection and Analysis of Abiotic Factors
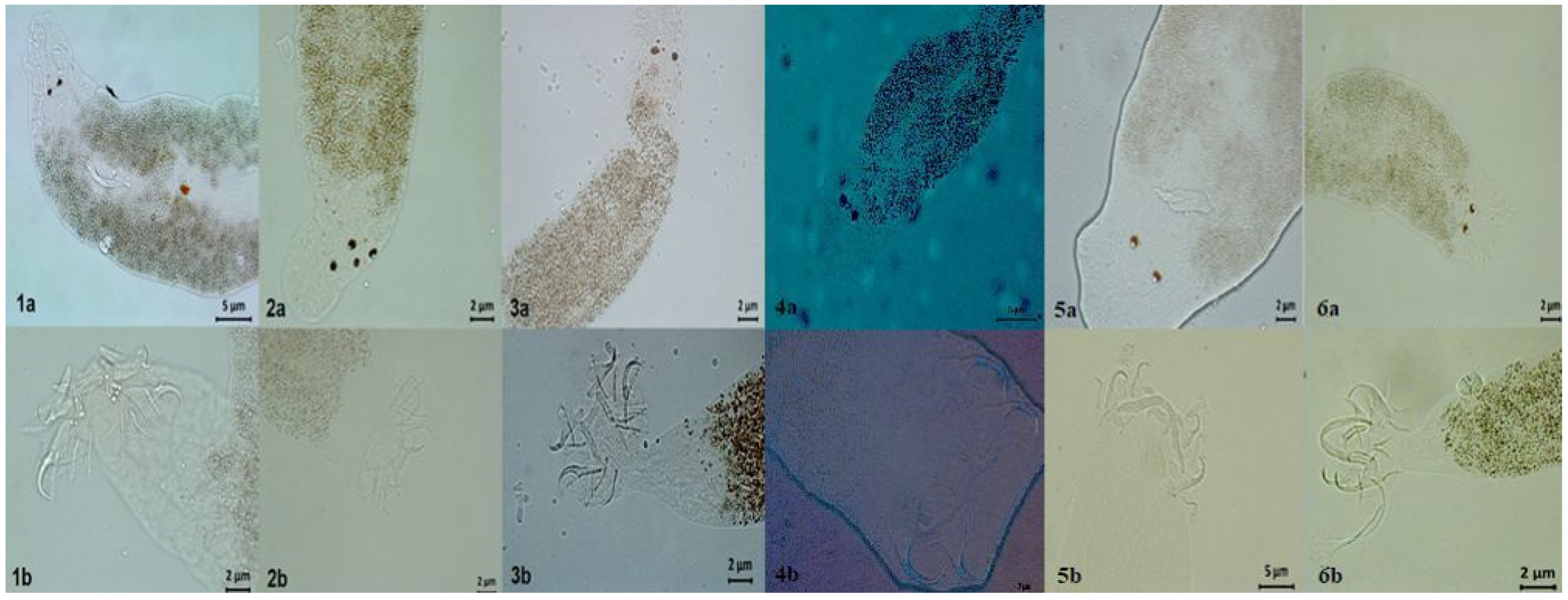
2.3. Parasitological Analysis
2.4. Statistical Analysis
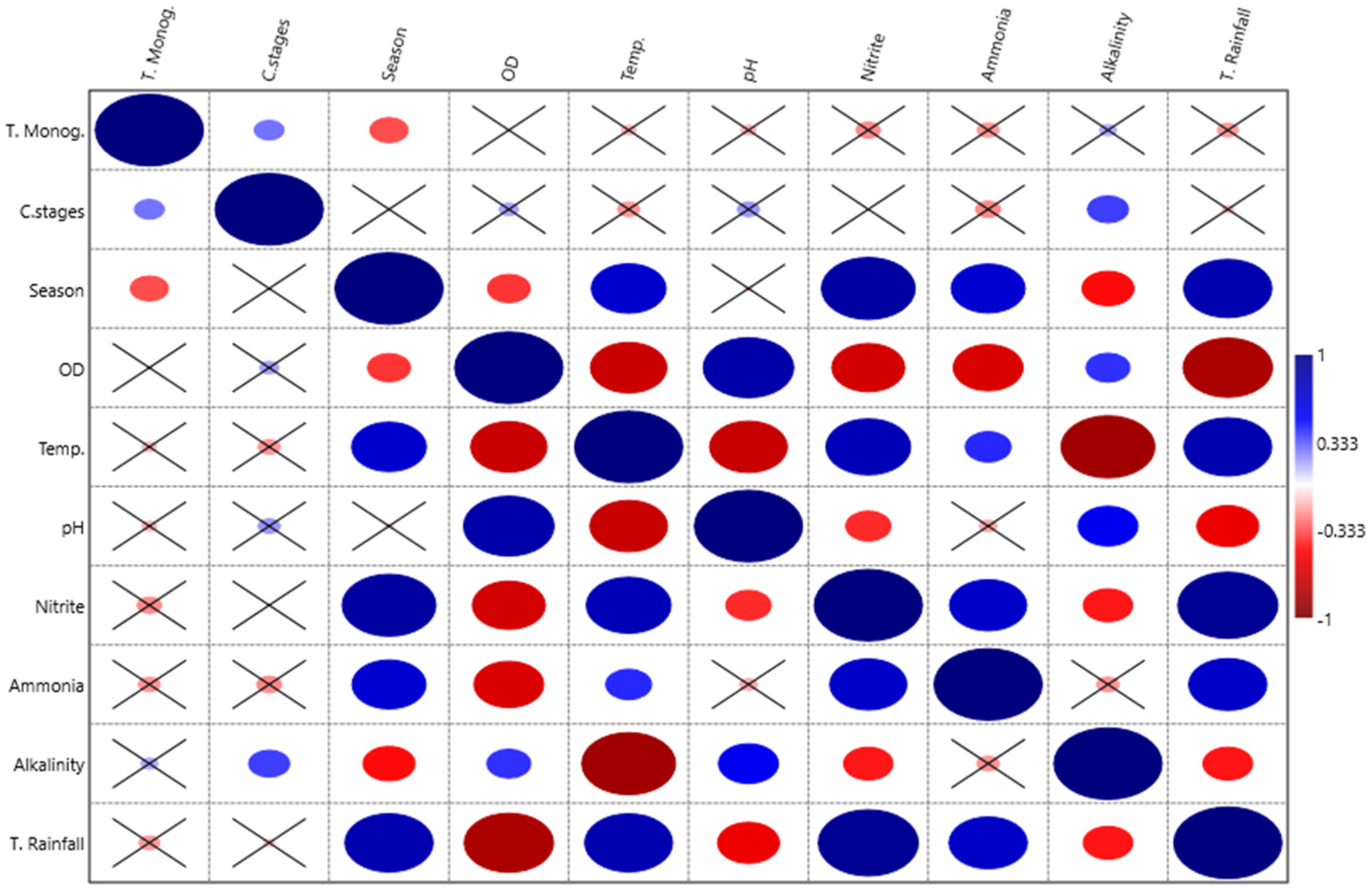
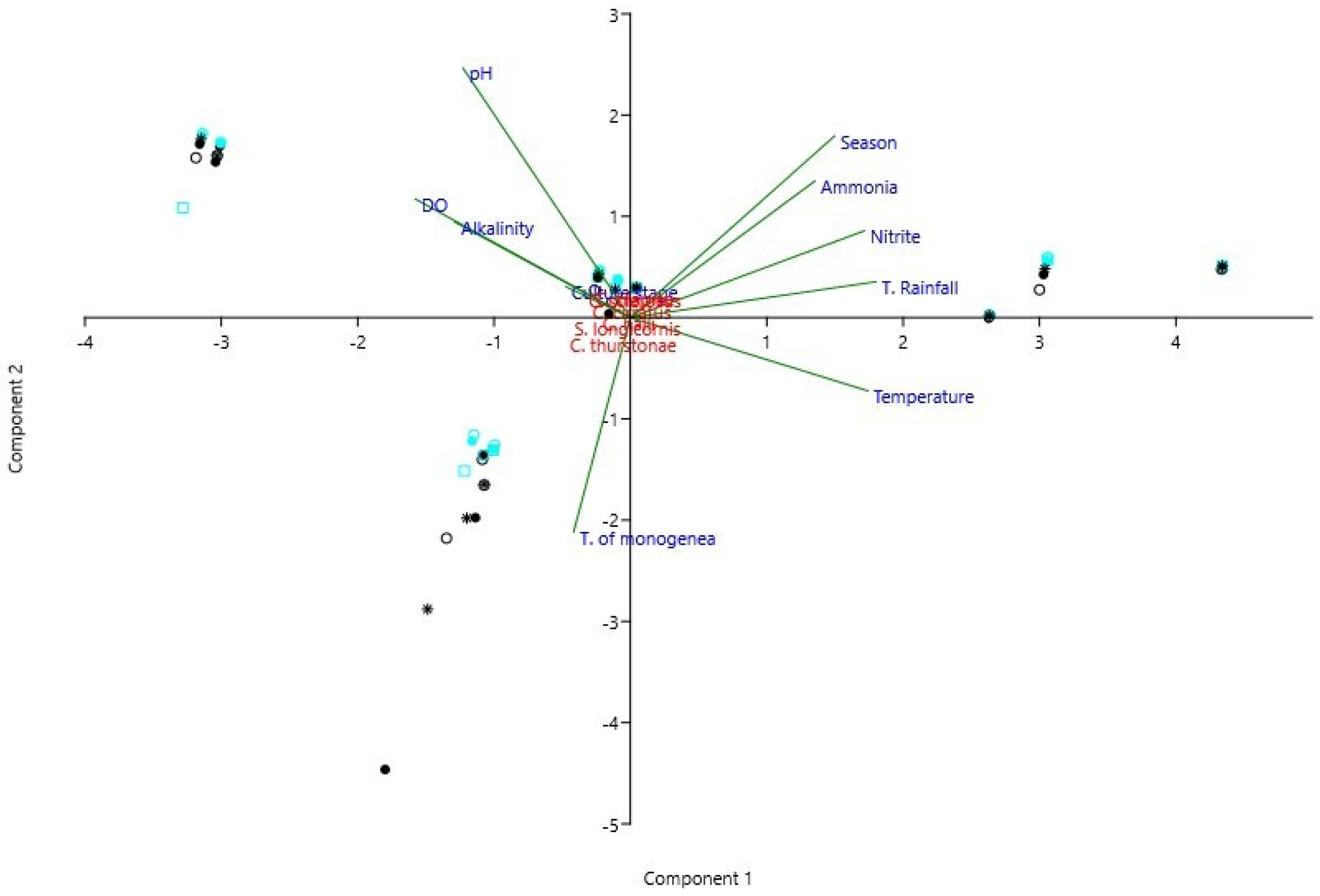
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roriz, G.D.; Delphino, M.K.V.C.; Gardner, I.A.; Gonçalves, V.S.P. Characterization of Tilapia Farming in Net Cages at a Tropical Reservoir in Brazil. Aquac. Rep. 2017, 6, 43–48. [Google Scholar] [CrossRef]
- Ergens, R. Nine Species of the Genus Cichlidogyrus Paperna, 1960 (Monogenea: Ancyrocephalinae) from Egyptian Fishes. Folia Parasitol. 1981, 28, 205–214. [Google Scholar]
- Jiménez-García, M.I.; Vidal-Martínez, V.M.; López-Jiménez, S. Monogeneans in Introduced and Native Cichlids in México: Evidence for Transfer. J. Parasitol. 2001, 87, 907–909. [Google Scholar] [CrossRef] [PubMed]
- Pariselle, A.; Euzet, L. Systematic Revision of Dactylogyridean Parasites (Monogenea) from Cichlid Fishes in Africa, the Levant and Madagascar. Zoosystema 2009, 31, 849–898. [Google Scholar] [CrossRef]
- Shinn, A.P.; Avenant-Oldewage, A.; Bondad-Reantaso, M.G.; Cruz-Laufer, A.J.; García-Vásquez, A.; Hernández-Orts, J.S.; Kutchta, R.; Longshaw, M.; Metselaar, M.; Pariselle, A.; et al. A global review of problematic and pathogenic parasites of farmed tilapia. Rev. Aquac. 2023, 15, 92–153. [Google Scholar] [CrossRef]
- Jiménez-García, I.; Rojas-García, C.R.; Mendoza-Franco, E.F. Ectoparasitic Infection in Nile Tilapia (Oreochromis Niloticus) Fry during Male Reversal in Veracruz, México. Int. Aquat. Res. 2020, 12, 197–207. [Google Scholar]
- Acosta-Pérez, V.-J.; Vega-Sánchez, V.; Fernández-Martínez, T.-E.; Zepeda-Velázquez, A.-P.; Reyes-Rodríguez, N.-E.; Ponce-Noguez, J.-B.; Peláez-Acero, A.; de-la-Rosa-Arana, J.-L.; Gómez-De-Anda, F.-R. Physicochemical Water Quality Influence on the Parasite Biodiversity in Juvenile Tilapia (Oreochromis spp.) Farmed at Valle Del Mezquital in the Central-Eastern Socioeconomic Region of Mexico. Pathogens 2022, 11, 1076. [Google Scholar] [CrossRef]
- Paperna, I. Parasites, Infections and Diseases of Fishes in Africa: An Update; CIFA Technical Paper; CABI: Wallingford, UK, 1996; Volume 31, p. 177. [Google Scholar]
- Jerônimo, G.T.; Speck, G.M.; Cechinel, M.; Gonçalves, E.; Martins, M.L. Seasonal Variation on the Ectoparasitic Communities of Nile Tilapia Cultured in Three Regions in Southern Brazil. Braz. J. Biol. 2011, 71, 365–373. [Google Scholar] [CrossRef]
- Jerônimo, G.T.; Ventura, A.S.; Pádua, S.B.; Porto, E.L.; Ferreira, L.C.; Ishikawa, M.M.; Martins, M.L. Parasitological Assessment in Hybrids Serrasalmidae Fish Farmed in Brazil. Braz. J. Vet. Parasitol. 2020, 29, s1984–s29612020084. [Google Scholar] [CrossRef]
- Cruz, M.G.; Jerônimo, G.T.; Bentes, S.P.C.; Gonçalves, L.U. Trichlorfon Is Effective against Dawestrema Cycloancistrium and Does Not Alter the Physiological Parameters of Arapaima (Arapaima Gigas): A Large Neotropical Fish from the Amazon. J. Fish Dis. 2022, 45, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-H.; Shoemaker, C.A.; Klesius, P.H. Evaluation of the Link between Gyrodactylosis and Streptococcosis of Nile Tilapia, Oreochromis Niloticus (L.). J. Fish Dis. 2007, 30, 233–238. [Google Scholar] [CrossRef]
- Tavares-Dias, M.; Martins, M.L. An Overall Estimation of Losses Caused by Diseases in the Brazilian Fish Farms. J. Paras. Dis. 2017, 41, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, L.D.; Gouveia, E.J.; Leal, F.C.; Figueiró, C.S.M.; Rojas, S.S.; Russo, M.R. Responses of Monogenean Species to Variations in Abiotic Parameters in Tilapiculture. J. Helminthol. 2020, 94, e186. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Pellitero, P. Fish Immunity and Parasite Infections: From Innate Immunity to Immunoprophylactic Prospects. Vet. Immunol. Immunopathol. 2008, 126, 171–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, C.; Yang, T. Killing Effect of Fish Serum and Mucus on Gyrodactylus Cichlidarum (Platyhelminthes, Monogenea) Mediated by Complement. Aquaculture 2023, 567, 739248. [Google Scholar] [CrossRef]
- Rubiogodoy, M.; Porter, R.; Tinsley, R. Evidence of Complement-Mediated Killing of Discocotyle Sagittata (Platyhelminthes, Monogenea) Oncomiracidia. F. Shell Immunol. 2004, 17, 95–103. [Google Scholar] [CrossRef]
- Buchmann, K. Binding and Lethal Effect of Complement from Oncorhynchus Mykiss on Gyrodactylus Derjavini (Platyhelminthes: Monogenea). Dis. Aquat. Organ. 1998, 32, 195–200. [Google Scholar] [CrossRef]
- Buchmann, K.; Bresciani, J. Monogenea (Phylum Platyhelminthes). In Fish Diseases and Disorders: Protozoan and Metazoan Infections; Oxfordshire: Wallingford, UK, 2006; Volume 1, pp. 297–344. [Google Scholar]
- Esch, G.W.; Shostak, A.W.; Marcogliese, D.J.; Goater, T.M. Patterns and Processes in Helminth Parasite Communities: An Overview. In Parasite Communities: Patterns and Processes; Springer: Dordrecht, The Netherlands, 1990; pp. 1–19. [Google Scholar]
- Rohde, K.; Hayward, C.; Heap, M. Aspects of the Ecology of Metazoan Ectoparasites of Marine Fishes. Int. J. Parasitol. 1995, 25, 945–970. [Google Scholar] [CrossRef]
- Lamková, K.; Simková, A.; Palíková, M.; Jurajda, P.; Lojek, A. Seasonal Changes of Immunocompetence and Parasitism in Chub (Leuciscus Cephalus), a Freshwater Cyprinid Fish. Parasitol. Res. 2007, 101, 775–789. [Google Scholar] [CrossRef]
- Akoll, P.; Fioravanti, M.L.; Konecny, R.; Schiemer, F. Infection Dynamics of Cichlidogyrus Tilapiae and C. Sclerosus (Monogenea, Ancyrocephalinae) in Nile Tilapia (Oreochromis Niloticus L.) from Uganda. J. Helminthol. 2012, 86, 302–310. [Google Scholar] [CrossRef]
- Hoai, T.D. Reproductive Strategies of Parasitic Flatworms (Platyhelminthes, Monogenea): The Impact on Parasite Management in Aquaculture. Aquac. Int. 2020, 28, 421–447. [Google Scholar] [CrossRef]
- Jerônimo, G.T.; Cruz, M.G.; Bertaglia, E.D.A.; Furtado, W.E.; Martins, M.L. Fish Parasites Can Reflect Environmental Quality in Fish Farms. Rev. Aquac. 2022, 14, 1558–1571. [Google Scholar] [CrossRef]
- Félix-Cuencas, L.; García-Trejo, J.F.; López-Tejeida, S.; León-Ramírez, J.J.D.; Soto-Zarazúa, G.M. Effect of three productive stages of tilapia (Oreochromis niloticus) under hyper-intensive recirculation aquaculture system on the growth of tomato (Solanum lycopersicum). Lat. Am. J. Aquat. Res. 2011, 49, 689–701. [Google Scholar] [CrossRef]
- Poulin, R.; Leung, T.L.F. Body size, trophic level, and the use of fish as transmission routes by parasites. Oecologia 2011, 166, 731–738. [Google Scholar] [CrossRef]
- Eiras, J.D.C.; Takemoto, R.M.; Pavanelli, G.C. Métodos de Estudo e Técnicas Laboratoriais Em Parasitologia de Peixes; EDUEM: Maringá, Brazil, 2000; Volume 1. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeo.Electron. 2001, 4, 9. [Google Scholar]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for Parasitologists—A Primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- Paperna, I.; Thurston, J.P. Monogenetic Trematodes Collected from Cichlid Fish in Uganda; Including the Description of Five New Species of Cichlidogyrus. Ver. Zool. Bot. Afr. 1969, 79, 15–33. [Google Scholar]
- Douëllou, L. Monogeneans of the Genus Cichlidogyrus Paperna, 1960 (Dactylogyridae: Ancyrocephalinae) from Cichlid Fishes of Lake Kariba (Zimbabwe) with Descriptions of Five New Species. Syst. Parasitol. 1993, 25, 159–186. [Google Scholar] [CrossRef]
- Pariselle, A.; Bilong Bilong, C.F.; Euzet, L. Four New Species of Cichlidogyrus Paperna, 1960 (Monogenea, Ancyrocephalidae), All Gill Parasites from African Mouthbreeder Tilapias of the Genera Sarotherodon and Oreochromis (Pisces, Cichlidae), with a Redescription of C. thurstonae Ergens, 1981. Syst. Parasitol. 2003, 56, 201–210. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575. [Google Scholar] [CrossRef]
- Ranzani-Paiva, M.J.; Felizardo, N.N.; Luque, J.L. Parasitological and Hematological Analysis of Nile Tilapia Oreochromis Niloticus Linnaeus, 1757 from Guarapiranga Reservoir, São Paulo State, Brazil. Acta Sci. Biol. Sci. 2005, 27, 231–237. [Google Scholar] [CrossRef]
- Ghiraldell, L.; Martins, M.L.; Jerônimo, G.T.; Yamashita, M.M.; Adamante, W.d.B. Ectoparasites Communities from Oreochromis Niloticus Cultivated in the State of Santa Catarina, Brazil. J. Fish Aquat. Sci. 2006, 1, 181–190. [Google Scholar] [CrossRef]
- Lizama, M.D.L.A.P.; Takemoto, R.M.; Ranzani-Paiva, M.J.T.; Ayroza, L.M.S.; Pavanelli, G.C. Relação Parasito-Hospedeiro Em Peixes de Pisciculturas Da Região de Assis, Estado de São Paulo, Brasil. 2. Piaractus Mesopotamicus (Holmberg, 1887). Acta Sci. Biol. Sci. 2008, 29, 437–445. [Google Scholar] [CrossRef]
- Pantoja MF, W.M.; Neves, L.R.; Dias, M.R.D.; Marinho, R.G.B.; Montagner, D.; Tavares-Dias, M. Protozoan and Metazoan Parasites of Nile Tilapia Oreochromis Niloticus Cultured in Brazil. Rev. MVZ Cordoba 2012, 17, 2812–2819. [Google Scholar] [CrossRef]
- Martins, M.L.; deSá, A.R.S.; Jerônimo, G.T.; Tancredo, K.R.; Gonçalves, E.L.T.; Bambi, D.; Speck, G.M.; Sandim, A.M. Microhabitat Preference and Seasonality of Gill Monogeneans in Nile Tilapia Reared in Southern Brazil. Neotrop. Helminthol. 2014, 8, 47–58. [Google Scholar]
- Zago, A.C.; Franceschini, L.; Garcia, F.; Schalch, S.H.C.; Gozi, K.S.; da Silva, R.J. Ectoparasites of Nile Tilapia (Oreochromis Niloticus) in Cage Farming in a Hydroelectric Reservoir in Brazil. Braz. J. Vet. Parasitol. 2014, 23, 171–178. [Google Scholar] [CrossRef]
- Brito, Y.C.T.; Silva-Souza, Â.T. Temporal Variation of Monogenoideans Component Community in the Gills of Oreochromis Niloticus (Cichlidae) in Fish Farming in Northern Parana State, Brazil. Panam. J. Aquat. Sci. 2017, 12, 333–342. [Google Scholar]
- Aparecido, L.E.O.; de Rolim, G.S.; Richetti, J.; de Souza, P.S.; Johann, J.A. Köppen, Thornthwaite and Camargo Climate Classifications for Climatic Zoning in the State of Paraná, Brazil. Ciência Agrotecnologia 2016, 40, 405–417. [Google Scholar] [CrossRef]
- Gannicott, A.M.; Tinsley, R.C. Environmental Effects on Transmission of Discocotyle Sagittata (Monogenea): Egg Production and Development. Parasitology 1998, 117, 499–504. [Google Scholar] [CrossRef]
- Da Silva, P.E.; Santos-Silva, C.M.; Spyrides, M.H.C.; Andrade, L.d.M.B. Precipitation and Air Temperature Extremes in the Amazon and Northeast Brazil. Int. J. Clim. 2019, 39, 579–595. [Google Scholar] [CrossRef]
- Pech, D.; Aguirre-Macedo, M.L.; Lewis, J.W.; Vidal-Martínez, V.M. Rainfall Induces Time-Lagged Changes in the Proportion of Tropical Aquatic Hosts Infected with Metazoan Parasites. Int. J. Parasitol. 2010, 40, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Fey, D.; Benítez-Villa, G.E.; Pérez-Ponce de León, G.; Rubio-Godoy, M. Population Dynamics of Cichlidogyrus Spp. and Scutogyrus Sp. (Monogenea) Infecting Farmed Tilapia in Veracruz, México. Aquaculture 2015, 443, 11–15. [Google Scholar] [CrossRef]
- Le Roux, L.; Avenant-Oldewage, A.; Van der Walt, F.C. Aspects of the ecology of Cichlidogyrus philander collected from Pseudocrenilabrus philander philander from the Padda Dam, Gauteng, South Africa. Afr. Zool. 2011, 46, 103–116. [Google Scholar] [CrossRef]
- Ibrahim, M.M. Variation in Parasite Infracommunies of Tilapia Zillii in Relation to Some Biotic and Abiotic Factors. Int. J. Zoo Re. 2012, 8, 59–70. [Google Scholar] [CrossRef]
- Zapata, A.; Diez, B.; Cejalvo, T.; Gutiérrez-de Frías, C.; Cortés, A. Ontogeny of the Immune System of Fish. Fish Shellfish. Immunol. 2006, 20, 126–136. [Google Scholar] [CrossRef]
- Muñoz, G.; Zamora, L. Ontogenetic Variation in Parasite Infracommunities of the Clingfish Sicyases Sanguineus(Pisces: Gobiesocidae). J. Parasitol. 2011, 97, 14–19. [Google Scholar] [CrossRef]
- Zander, C.D. Four-Year Monitoring of Parasite Communities in Gobiid Fishes of the South-Western Baltic. Parasitol. Res. 2003, 90, 502–511. [Google Scholar] [CrossRef]
- Suliman, E.A.M.; Al-Harbi, A.H. Prevalence and seasonal variation of ectoparasites in cultured Nile tilapia Oreochromis niloticus in Saudi Arabia. J. Parasit. Dis. 2016, 40, 1487–1493. [Google Scholar] [CrossRef]
- Santos, G.G.; da Silva, A.; Sousa, B.O.; Tavares-Dias, M. Parasite fauna of tambaqui reared in net-cages at two stocking densities. Bol. Inst.Pesca 2019, 45, e492. [Google Scholar] [CrossRef]
- Bowden, T.J. Modulation of the Immune System of Fish by Their Environment. Fish Shellfish. Immunol. 2008, 25, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.G.; Pante, E.; Levesque, E.M.; Roumillat, W.A.; de Buron, I. Metamicrocotyla Macracantha, a Polyopisthocotylid Gill Parasite of the Striped Mullet, Mugil Cephalus: Population Dynamics in South Carolina Estuaries. Parasitol. Res. 2008, 102, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- Shah, H.B.; Yousuf, A.R.; Chishti, M.Z.; Ahmad, F. Seasonal Changes in Infrapopulations of Diplozoon Kashmirensis Kaw, 1950 (Monogenea: Diplozoidae) along a Eutrophic Gradient. Parasitol. Res. 2013, 112, 3347–3356. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, L.; Quilichini, Y.; Marchand, B. Sparicotyle Chrysophrii (Van Beneden and Hesse 1863) (Monogenea: Polyopisthocotylea) Parasite of Cultured Gilthead Sea Bream Sparus Aurata (Linnaeus 1758) (Pisces: Teleostei) from Corsica: Ecological and Morphological Study. Parasitol. Res. 2010, 107, 389–398. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Indices | Autumn | Winter | Spring | Summer | p Value |
|---|---|---|---|---|---|---|
| C. sclerosus | N | 28 a | 7 a | 4 a | 10 a | 0.0006 |
| P% | 31.7% a | 10.0% b | 6.7% b | 11.7% b | 0.0015 | |
| MI ± SD | 1.47 ± 0.69 | 1.75 ± 0.95 | 1.00 ± 0.00 | 1.42 ± 0.78 | 0.422 | |
| MA ± SD | 0.46 ± 0.79 | 0.17 ± 0.59 | 0.06 ± 0.25 | 0.16 ± 0.52 | 0.675 | |
| C. thurstonae | N | 1130 a | 77 b | 109 b | 57 b | 0.000 |
| P% | 76.7% a | 30.0% b | 31.7% b | 20.0% b | <0.0001 | |
| MI ± SD | 24.57 ± 30.80 a | 6.41 ± 7.89 b | 5.73 ± 13.21 b | 4.75 ± 3.04 b | 0.001 | |
| MA ± SD | 18.83 ± 28.87 a | 1.92 ± 5.14 b | 1.81 ± 7.77 b | 0.95 ± 2.32 b | 0.002 | |
| C. tilapiae | N | 1 | 0 | 3 | 1 | 0.841 |
| P% | 1.7% | 0.0% | 3.3% | 1.7% | 0.9090 | |
| MI ± SD | 1.00 ± NA | - | 1.50 ± 0.70 | 1.00 ± NA | - | |
| MA ± SD | 0.01 ± 0.12 | 0.00 ± 0.00 | 0.05 ± 0.28 | 0.01 ± 0.12 | 0.468 | |
| S. longicornis | N | 558 a | 48 b | 37 b | 33 b | 0.0000 |
| P% | 65.0% a | 32.5% b | 25.0% b | 15.0% b | <0.0001 | |
| MI ± SD | 14.31 ± 13.12 a | 3.69 ± 2.78 b | 2.46 ± 1.40 b | 3.66 ± 2.91 b | 0.001 | |
| MA ± SD | 9.30 ± 12.58 a | 1.20 ± 2.33 b | 0.61 ± 1.27 b | 0.55 ± 1.70 b | 0.001 | |
| C. halli | N | 529 a | 96 ab | 112 ab | 94 b | 0.0013 |
| P% | 78.3% a | 45.0% b | 41.7% b | 21.7% b | <0.0001 | |
| MI ± SD | 11.26 ± 12.43 a | 5.33 ± 5.47 b | 4.48 ± 7.50 b | 7.23 ± 8.35 b | 0.028 | |
| MA ± SD | 8.81 ± 11.93 a | 2.40 ± 4.50 b | 1.86 ± 5.27 b | 1.56 ± 4.82 b | 0.001 | |
| C. cirratus | N | 106 a | 200 a | 41 ab | 9 b | 0.0000 |
| P% | 30.0% b | 47.5% a | 25.0% b | 10.0% b | 0.0004 | |
| MI ± SD | 5.88 ± 5.02 | 10.53 ± 21.36 | 2.73 ± 2.12 | 1.50 ± 0.83 | 0.69 | |
| MA ± SD | 1.76 ± 3.83 | 5.00 ± 15.46 | 0.68 ± 1.57 | 0.15 ± 0.51 | 0.174 | |
| Total | N | 2352 a | 428 b | 306 bc | 204 c | 0.000 |
| Species | Indices | Fingerling | Juvenile | Adult | p Value |
|---|---|---|---|---|---|
| C. sclerosus | N | 1 b | 16 b | 32 a | 0.0003 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 1.7% b | 15.0% ab | 26.2% a | 0.0001 | |
| MI±SD | 1.00 ± NA | 1.33 ± 0.65 | 1.52 ± 0.75 | - | |
| MA±SD | 0.01 ± 0.12 b | 0.20 ± 0.53 ab | 0.40 ± 0.77 a | 0.007 | |
| C. thurstonae | N | 95 b | 333 a | 945 a | 0.000 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 30.0% | 40.0% | 48.8% | 0.0841 | |
| MI±SD | 5.27 ± 3.95 b | 10.41 ± 13.36 ab | 24.23 ± 33.68 a | 0.046 | |
| MA±SD | 1.58 ± 3.23 | 4.16 ± 9.81 | 11.81 ± 26.35 | 0.08 | |
| C. tilapiae | N | 1 | 2 | 2 | 0.8418 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 1.7% | 1.2% | 2.5% | 1.000 | |
| MI±SD | 1.00 ± NA | 2.00 ± NA | 1.00 ± 0.00 | - | |
| MA±SD | 0.01 ± 0.12 | 0.02 ± 0.22 | 0.02 ± 0.15 | 0.959 | |
| S. longicornis | N | 12 | 164 | 500 | 0.0564 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 11.7% b | 40.0% a | 46.2% a | <0.0001 | |
| MI±SD | 1.71 ± 1.49 b | 5.12 ± 6.73 a | 13.51 ± 12.98 a | 0.016 | |
| MA±SD | 0.20 ± 0.73 b | 2.05 ± 4.91 a | 6.25 ± 11.08 a | 0.014 | |
| C. halli | N | 171 c | 199 b | 461 a | 0.0000 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 33.3% b | 32.5% b | 71.2% a | <0.0001 | |
| MI±SD | 8.55 ± 6.33 | 7.65 ± 10.72 | 8.08 ± 11.26 | 0.965 | |
| MA±SD | 2.85 ± 5.42 | 2.48 ± 7.02 | 5.76 ± 10.17 | 0.154 | |
| C. cirratus | N | 0 b | 49 b | 307 a | 0.0000 |
| Kn | 1.00 ± 0.03 | 1.00 ± 0.02 | 0.99 ± 0.01 | 0.9982 | |
| P% | 0.0% b | 22.5% b | 50.0% a | <0.0001 | |
| MI±SD | 0 | 2.72 ± 1.87 | 7.67 ± 15.21 | 0.146 | |
| MA±SD | 0.00 ± 0.00 b | 0.61 ± 1.43 b | 3.83 ± 11.36 a | 0.032 | |
| Total | N | 280 c | 763 b | 2247 a | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertaglia, E.d.A.; Furtado, W.E.; Silva e Souza, Â.T.; Fernandes, M.C.; Pereira, S.A.; Brasil, E.M.; Mouriño, J.L.P.; Jerônimo, G.T.; Martins, M.L. Influence of Seasonality and Culture Stage of Farmed Nile Tilapia (Oreochromis niloticus) with Monogenean Parasitic Infection. Animals 2023, 13, 1525. https://doi.org/10.3390/ani13091525
Bertaglia EdA, Furtado WE, Silva e Souza ÂT, Fernandes MC, Pereira SA, Brasil EM, Mouriño JLP, Jerônimo GT, Martins ML. Influence of Seasonality and Culture Stage of Farmed Nile Tilapia (Oreochromis niloticus) with Monogenean Parasitic Infection. Animals. 2023; 13(9):1525. https://doi.org/10.3390/ani13091525
Chicago/Turabian StyleBertaglia, Elisabeth de Aguiar, William Eduardo Furtado, Ângela Teresa Silva e Souza, Manoela Clemente Fernandes, Scheila Anelise Pereira, Elenice Martins Brasil, José Luiz Pedreira Mouriño, Gabriela Tomas Jerônimo, and Maurício Laterça Martins. 2023. "Influence of Seasonality and Culture Stage of Farmed Nile Tilapia (Oreochromis niloticus) with Monogenean Parasitic Infection" Animals 13, no. 9: 1525. https://doi.org/10.3390/ani13091525