
Modeling Co-Infection by Streptococcus suis and Haemophilus parasuis Reveals Influences on Biofilm Formation and Host Response
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Mono-Culture and Co-Culture Planktonic Growth Assays In Vitro
2.3. Establishment of Single and Mixed Biofilms In Vitro
2.4. Measurement of Biofilm Biomass
2.5. Observation of Biofilms by Confocal Laser Scanning Microscopy
2.6. Observation of Biofilms by Scanning Electron Microscope
2.7. Analysis of Gene Expression in Mixed Planktonic and Biofilm Cells
2.8. Antibiotic Susceptibility of Planktonic and Biofilm Cultures
2.9. Establishment of Mixed Infection Mice Model
2.10. Determination of Live Bacteria in Organs
2.11. Inflammatory Factor Expression Assay
2.12. Statistical Analysis and Interpretative Criteria
3. Results
3.1. Competition between S. suis and H. parasuis during Co-Culture In Vitro
3.2. Formation of Mixed Biofilms of S. suis and H. parasuis In Vitro
3.2.1. Single or Mixed Biofilms were Visualized by CLSM
3.2.2. 3D images of Single and Mixed Biofilms Visualized by Scanning Electron Microscopy
3.2.3. S. suis and H. parasuis Were Inhibited When Cultured in Mixed Biofilms
3.3. Mixed Culture of S. suis and H. parasuis Decrease Antibiotic Susceptibility
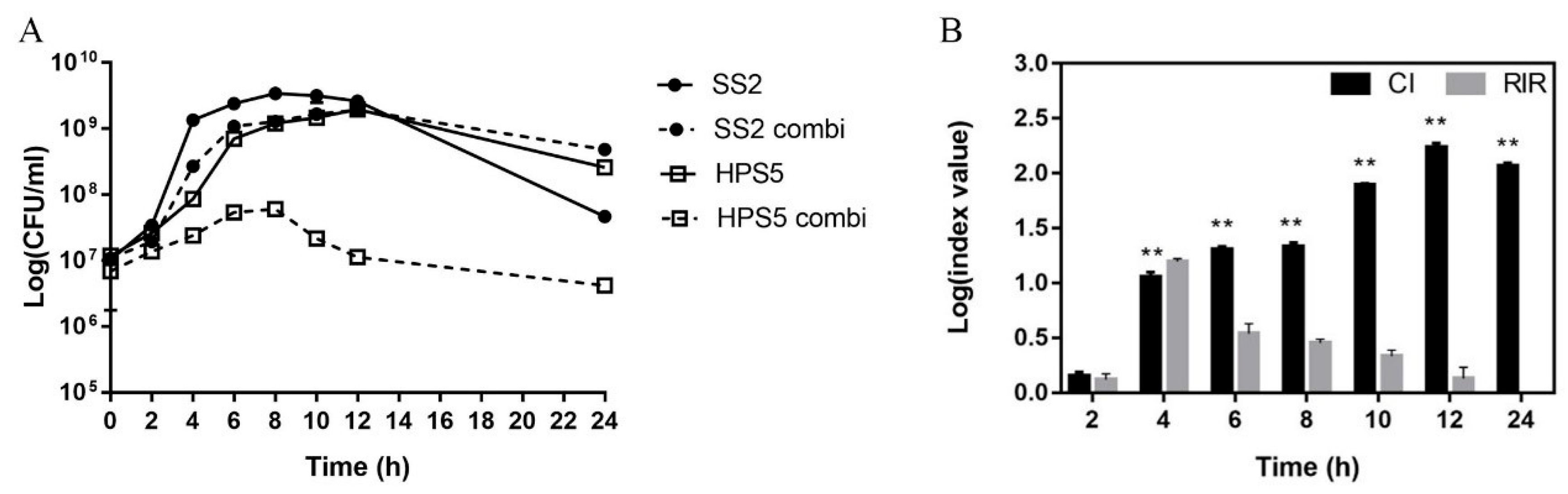
3.4. H. parasuis Has a Significant Effect on the Virulence of S. suis in In-Vitro Co-Culture
3.5. Coexistence of S. suis and H. parasuis Increase the Bacterial Load in Mice
3.6. Single and Mixed Infections Affect Inflammatory Factor Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primers Sequence (5′–3′) | Target Gene |
|---|---|---|
| CPS2-F | ATTGGTAGGCACTGTCGTTGGTC | cps2 |
| CPS2-R | AGAACTTAGCATTGTTGCGGTGG | cps2 |
| EF-F | TCCAATCACAGATCCAGATAGCG | ef |
| EF-R | CTGACCCATTTGGACCATCTAAG | ef |
| MRP-F | CAAGGAAAGTGAACAGAACGAGC | mrp |
| MRP-A | TAGTCGTCCAAACCTGAGTAGCG | mrp |
| PDH-F | CGCGAATTCATGCAACAAATCCGTGAT | pdh |
| PDH-R | CCCTCGAGGCTAGTCTACAAACACATC | pdh |
| LUXS-F | ATGAAAAAAGAAGTCACT | luxs |
| LUXS-R | TTAGATTGGTTTTCTTTC | luxs |
| 16S RNA-F | GTTGCGAACGGGTGAGTAA | 16sRNA |
| 16S RNA-R | TCTCAGGTCGGCTATGTATCG | 16sRNA |
| CAPD-F | ATGTTAATGCCATTAATTTATTCATTG | CapD |
| CAPD-R | TCGAACCGATAGAACCAGCAGCACCAGTC | CapD |
| CLPX-F | AGAGTGAGGGCGTTGAGT | clpX |
| CLPX-R | TTCTTGTTTCGGGTGTTT | clpX |
| GROUP 1-F | TTTAGGTAAAGATAAGCAAGGAAATCC | Group 1 |
| GROUP 1-R | CCACACAAAACCTACCCCTCCTCC | Group 1 |
| LUXS-F | GCATCAGCAAGAGAATGTTCCT | luxs |
| LUXS-R | ATGTCGTTAATTGGTTCACCTTCA | luxs |
| OMPP2-F | ATGAAAAAAACACTAGTAGCA | ompP2 |
| OMPP2-R | TTACCATAATACACGTAAACC | ompP2 |
| 16S RNA-F | GGCTTCGTCACCCTCTGT | 16sRNA |
| 16S RNA-R | GTGATGAGGAAGGGTGGTGT | 16sRNA |
| IL-6-F | CTGCAAGAGACTTCCATCCAG | IL-6 |
| IL-6-R | AGTGGTATAGACAGGTCTGTTGG | IL-6 |
| IL-12-F | GTCCTCAGAAGCTAACCATCTCC | IL-12 |
| IL-12-R | CCAGAGCCTATGACTCCATGTC | IL-12 |
| IL-1β-F | GAAATGCCACCTTTTGACAGTG | IL-1β |
| IL-1β-R | TGGATGCTCTCATCAGGACAG | IL-1β |
| MCP-F | GCATCCACGTGTTGGCTCA | MCP-1 |
| MCP-R | CTCCAGCCTACTCATTGGGATC | MCP-1 |
| TNF-α-F | CAGGCGGTGCCTATGTCTC | TNF-α |
| TNF-α-R | CGATCACCCCGAAGTTCAGTAG | TNF-α |
| IFN-γ-F | ACAGCAAGGCGAAAAAGGATG | IFN-γ |
| IFN-γ-R | TGGTGGACCACTCGGATGA | IFN-γ |
| GAPDH-F | AGGTCGGTGTGAACGGATTTG | gapdh |
| GAPDH-R | GGGGTCGTTGATGGCAACA | gapdh |
References
- Dutkiewicz, J.; Zajac, V.; Sroka, J.; Wasinski, B.; Cisak, E.; Sawczyn, A.; Kloc, A.; Wojcik-Fatla, A. Streptococcus suis: A re-emerging pathogen associated with occupational exposure to pigs or pork products. Part II—Pathogenesis. Ann. Agric. Environ. Med. 2018, 25, 186–203. [Google Scholar] [CrossRef]
- Dutkiewicz, J.; Sroka, J.; Zajac, V.; Wasinski, B.; Cisak, E.; Sawczyn, A.; Kloc, A.; Wojcik-Fatla, A. Streptococcus suis: A re-emerging pathogen associated with occupational exposure to pigs or pork products. Part I—Epidemiology. Ann. Agric. Environ. Med. 2017, 24, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, W.; Wu, Z.; Lu, C. Reduced virulence is an important characteristic of biofilm infection of Streptococcus suis. FEMS Microbiol. Lett. 2011, 316, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; Lu, C.; Sun, J.; Yan, Y. Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl. Environ. Microbiol. 2011, 77, 8272–8279. [Google Scholar] [CrossRef]
- Lee, K.W.K.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 2014, 8, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Tang, C.; Liao, M.; Yue, H. Update on the pathogenesis of Haemophilus parasuis infection and virulence factors. Vet. Microbiol. 2014, 168, 1–7. [Google Scholar] [CrossRef]
- Macedo, N.; Rovira, A.; Torremorell, M. Haemophilus parasuis: Infection, immunity and enrofloxacin. Vet. Res. 2015, 46, 128. [Google Scholar] [CrossRef] [PubMed]
- Bello-Orti, B.; Costa-Hurtado, M.; Martinez-Moliner, V.; Segales, J.; Aragon, V. Time course Haemophilus parasuis infection reveals pathological differences between virulent and non-virulent strains in the respiratory tract. Vet. Microbiol. 2014, 170, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.; Pijoan, C. Haemophilus parasuis: New trends on diagnosis, epidemiology and control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Li, C.; Wang, C.; Liu, Y.; Wang, G.; He, X.; Hu, L.; Liu, Y.; Cui, M.; et al. Secondary Haemophilus parasuis infection enhances highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection-mediated inflammatory responses. Vet. Microbiol. 2017, 204, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.; Oh, C.; Lee, K.; Cho, K.H. Survey of porcine respiratory disease complex-associated pathogens among commercial pig farms in Korea via oral fluid method. J. Vet. Sci. 2017, 18, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Denoncourt, A.; Letendre, C.; Auger, J.-P.; Segura, M.; Aragon, V.; Lacouture, S.; Gottschalk, M. Limited Interactions between Streptococcus Suis and Haemophilus Parasuis in In Vitro Co-Infection Studies. Pathogens 2018, 7, 7. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Costerton, W.J. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Hansen, M.F.; Roder, H.L.; Wang, N.; Burmolle, M.; He, G. Mixed-species biofilms in the food industry: Current knowledge and novel control strategies. Crit. Rev. Food Sci. Nutr. 2019, 60, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wuertz, S.; Okabe, S.; Hausner, M. Microbial communities and their interactions in biofilm systems: An overview. Water Sci. Technol. 2004, 49, 327. [Google Scholar] [CrossRef]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol. Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef]
- Reddinger, R.M.; Luke-Marshall, N.R.; Sauberan, S.L.; Hakansson, A.P.; Campagnari, A.A. Streptococcus pneumoniae Modulates Staphylococcus aureus Biofilm Dispersion and the Transition from Colonization to Invasive Disease. MBio 2018, 9, e02089-17. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; De Nicola, S.; Verginelli, F.; Fiscarelli, E.; Di Bonaventura, G. Cooperative pathogenicity in cystic fibrosis: Stenotrophomonas maltophilia modulates Pseudomonas aeruginosa virulence in mixed biofilm. Front. Microbiol. 2015, 6, 951. [Google Scholar] [CrossRef] [PubMed]
- Cope, E.K.; Goldstein-Daruech, N.; Kofonow, J.M.; Christensen, L.; McDermott, B.; Monroy, F.; Palmer, J.N.; Chiu, A.G.; Shirtliff, M.E.; Cohen, N.A.; et al. Regulation of virulence gene expression resulting from Streptococcus pneumoniae and nontypeable Haemophilus influenzae interactions in chronic disease. PLoS ONE 2011, 6, e28523. [Google Scholar] [CrossRef] [PubMed]
- Grenier, D.; Grignon, L.; Gottschalk, M. Characterisation of biofilm formation by a Streptococcus suis meningitis isolate. Vet. J. 2009, 179, 292–295. [Google Scholar] [CrossRef] [PubMed]
- M100-S25; Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2015.
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Liu, B.; Wang, S.; Li, J.; Gong, S.; Sun, L.; Li, Y. Pdh modulate virulence through reducing stress tolerance and biofilm formation of serotype 2. Virulence 2019, 10, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zumaquero, A.; Ortiz-Martin, I.; Beuzon, C.R. Competitive index in mixed infections: A sensitive and accurate assay for the genetic analysis of Pseudomonas syringae-plant interactions. Mol. Plant. Pathol. 2007, 8, 437–450. [Google Scholar] [CrossRef]
- Russel, J.; Roder, H.L.; Madsen, J.S.; Burmolle, M.; Sorensen, S.J. Antagonism correlates with metabolic similarity in diverse bacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 10684–10688. [Google Scholar] [CrossRef]
- Reece, E.; Doyle, S.; Greally, P.; Renwick, J.; McClean, S. Aspergillus fumigatus Inhibits Pseudomonas aeruginosa in Co-culture: Implications of a Mutually Antagonistic Relationship on Virulence and Inflammation in the CF Airway. Front. Microbiol. 2018, 9, 1205. [Google Scholar] [CrossRef]
- Kreth, J.; Merritt, J.; Shi, W.; Qi, F. Competition and Coexistence between Streptococcus mutans and Streptococcus sanguinis in the Dental Biofilm. J. Bacteriol. 2005, 187, 7193–7203. [Google Scholar] [CrossRef]
- An, D.; Danhorn, T.; Fuqua, C.; Parsek, M.R. Quorum sensing and motility mediate interactions between Pseudomonas aeruginosa and Agrobacterium tumefaciens in biofilm cocultures. Proc. Natl. Acad. Sci. USA 2006, 103, 3828–3833. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Sun, L.; Grenier, D.; Yi, L. The LuxS/AI-2 system of Streptococcus suis. Appl. Microbiol. Biotechnol. 2018, 102, 7231–7238. [Google Scholar] [CrossRef]
- Zhang, B.; Ku, X.; Zhang, X.; Zhang, Y.; Chen, G.; Chen, F.; Zeng, W.; Li, J.; Zhu, L.; He, Q. The AI-2/luxS Quorum Sensing System Affects the Growth Characteristics, Biofilm Formation, and Virulence of Haemophilus parasuis. Front. Cell Infect. Microbiol. 2019, 9, 62. [Google Scholar] [CrossRef]
- Egland, P.G.; Palmer, R.J., Jr.; Kolenbrander, P.E. Interspecies communication in Streptococcus gordonii-Veillonella atypica biofilms: Signaling in flow conditions requires juxtaposition. Proc. Natl. Acad. Sci. USA 2004, 101, 16917–16922. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Sun, L.; Grenier, D.; Yi, L. Streptococcus suis biofilm: Regulation, drug-resistance mechanisms, and disinfection strategies. Appl. Microbiol. Biotechnol. 2018, 102, 9121–9129. [Google Scholar] [CrossRef]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, C.E.; Hong, W.; Pang, B.; Weimer, K.E.; Juneau, R.A.; Turner, J.; Swords, W.E. Indirect pathogenicity of Haemophilus influenzae and Moraxella catarrhalis in polymicrobial otitis media occurs via interspecies quorum signaling. mBio 2010, 1, e00102-10. [Google Scholar] [CrossRef]
- Gabrilska, R.A.; Rumbaugh, K.P. Biofilm models of polymicrobial infection. Future Microbiol. 2015, 10, 1997–2015. [Google Scholar] [CrossRef] [PubMed]
- García-Bayona, L.; Comstock, L.E. Bacterial antagonism in host-associated microbial communities. Science 2018, 361, eaat2456. [Google Scholar] [CrossRef]
- Duan, K.; Dammel, C.; Stein, J.; Rabin, H.; Surette, M.G. Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol. Microbiol. 2003, 50, 1477–1491. [Google Scholar] [CrossRef]
- Domínguez-Punaro, M.C.; Segura, M.; Plante, M.M.; Lacouture, S.; Rivest, S.; Gottschalk, M. Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J. Immunol. 2007, 179, 1842–1854. [Google Scholar] [CrossRef]
- Seitz, M.; Beineke, A.; Seele, J.; Fulde, M.; Valentin-Weigand, P.; Baums, C.G. A novel intranasal mouse model for mucosal colonization by Streptococcus suis serotype 2. J. Med. Microbiol. 2012, 61, 1311–1318. [Google Scholar] [CrossRef]
- Williams, A.E.; Blakemore, W.F.; Alexander, T.J. A murine model of Streptococcus suis type 2 meningitis in the pig. Res. Vet. Sci. 1988, 45, 394–399. [Google Scholar] [CrossRef]
- Domínguez-Punaro, M.C.; Koedel, U.; Hoegen, T.; Demel, C.; Klein, M.; Gottschalk, M. Severe cochlear inflammation and vestibular syndrome in an experimental model of Streptococcus suis infection in mice. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 2391–2400. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Y.; Wen, Y.; Lau, G.W.; Huang, X.; Wu, R.; Yan, Q.; Huang, Y.; Zhao, Q.; Cao, S. HtrA Is Important for Stress Resistance and Virulence in Haemophilus parasuis. Infect. Immun. 2016, 84, 2209–2219. [Google Scholar] [CrossRef]
- Niu, K.; Meng, Y.; Liu, M.; Ma, Z.; Lin, H.; Zhou, H.; Fan, H. Phosphorylation of GntR reduces Streptococcus suis oxidative stress resistance and virulence by inhibiting NADH oxidase transcription. PLoS Pathog. 2023, 19, e1011227. [Google Scholar] [CrossRef]
- Gao, T.; Tan, M.; Liu, W.; Zhang, C.; Zhang, T.; Zheng, L.; Zhu, J.; Li, L.; Zhou, R. GidA, a tRNA Modification Enzyme, Contributes to the Growth, and Virulence of Streptococcus suis Serotype 2. Front. Cell. Infect. Microbiol. 2016, 6, 44. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, Y.; Zhou, J.; Zhu, L.; Gu, Y.; Zhang, X.; Li, X.; Feng, W. Roles of the Putative Type IV-like Secretion System Key Component VirD4 and PrsA in Pathogenesis of Streptococcus suis Type 2. Front. Cell. Infect. Microbiol. 2016, 6, 172. [Google Scholar] [CrossRef] [PubMed]
- Dai, K.; Yang, Z.; Ma, X.; Chang, Y.F.; Cao, S.; Zhao, Q.; Huang, X.; Wu, R.; Huang, Y.; Wen, Y.; et al. Deletion of Polyamine Transport Protein PotD Exacerbates Virulence in Glaesserella (Haemophilus) parasuis in the Form of Non-biofilm-generated Bacteria in a Murine Acute Infection Model. Virulence 2021, 12, 520–546. [Google Scholar] [CrossRef] [PubMed]







| Bacterial Combinations | MIC (μg/mL) (Planktonic) | MBC (μg/mL) (Planktonic) | MIC (μg/mL) (Biofilm) | MBC (μg/mL) (Biofilm) |
|---|---|---|---|---|
| Amoxicillin | ||||
| SS2 | 0.3125 | 2.5 | 5 | >320 |
| HPS5 | 0.1625 | 0.625 | 10 | >320 |
| SS2 + HPS5 | 5 | 10 | 10 | >320 |
| Gentamicin | ||||
| SS2 | 10 | 40 | 5 | >320 |
| HPS5 | 5 | 10 | 10 | >320 |
| SS2 + HPS5 | 20 | 40 | 20 | >320 |
| Enrofloxacin | ||||
| SS2 | 0.3125 | 0.625 | 1.25 | >320 |
| HPS5 | 0.1625 | 0. 625 | 0.625 | >320 |
| SS2 + HPS5 | 0.625 | 20 | 1.25 | >320 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, M.; Zuo, J.; Shen, Y.; Yuan, S.; Gao, S.; Wang, Y.; Wang, Y.; Yi, L. Modeling Co-Infection by Streptococcus suis and Haemophilus parasuis Reveals Influences on Biofilm Formation and Host Response. Animals 2023, 13, 1511. https://doi.org/10.3390/ani13091511
Gao M, Zuo J, Shen Y, Yuan S, Gao S, Wang Y, Wang Y, Yi L. Modeling Co-Infection by Streptococcus suis and Haemophilus parasuis Reveals Influences on Biofilm Formation and Host Response. Animals. 2023; 13(9):1511. https://doi.org/10.3390/ani13091511
Chicago/Turabian StyleGao, Mengxia, Jing Zuo, Yamin Shen, Shuo Yuan, Shuji Gao, Yuxin Wang, Yang Wang, and Li Yi. 2023. "Modeling Co-Infection by Streptococcus suis and Haemophilus parasuis Reveals Influences on Biofilm Formation and Host Response" Animals 13, no. 9: 1511. https://doi.org/10.3390/ani13091511