
Histological Variants of Squamous and Basal Cell Carcinoma in Squamates and Chelonians: A Comprehensive Classification

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissues
2.2. Histopathology
2.3. Immunohistochemistry
2.4. Statistical Analysis
3. Results
3.1. Histological Re-Classification
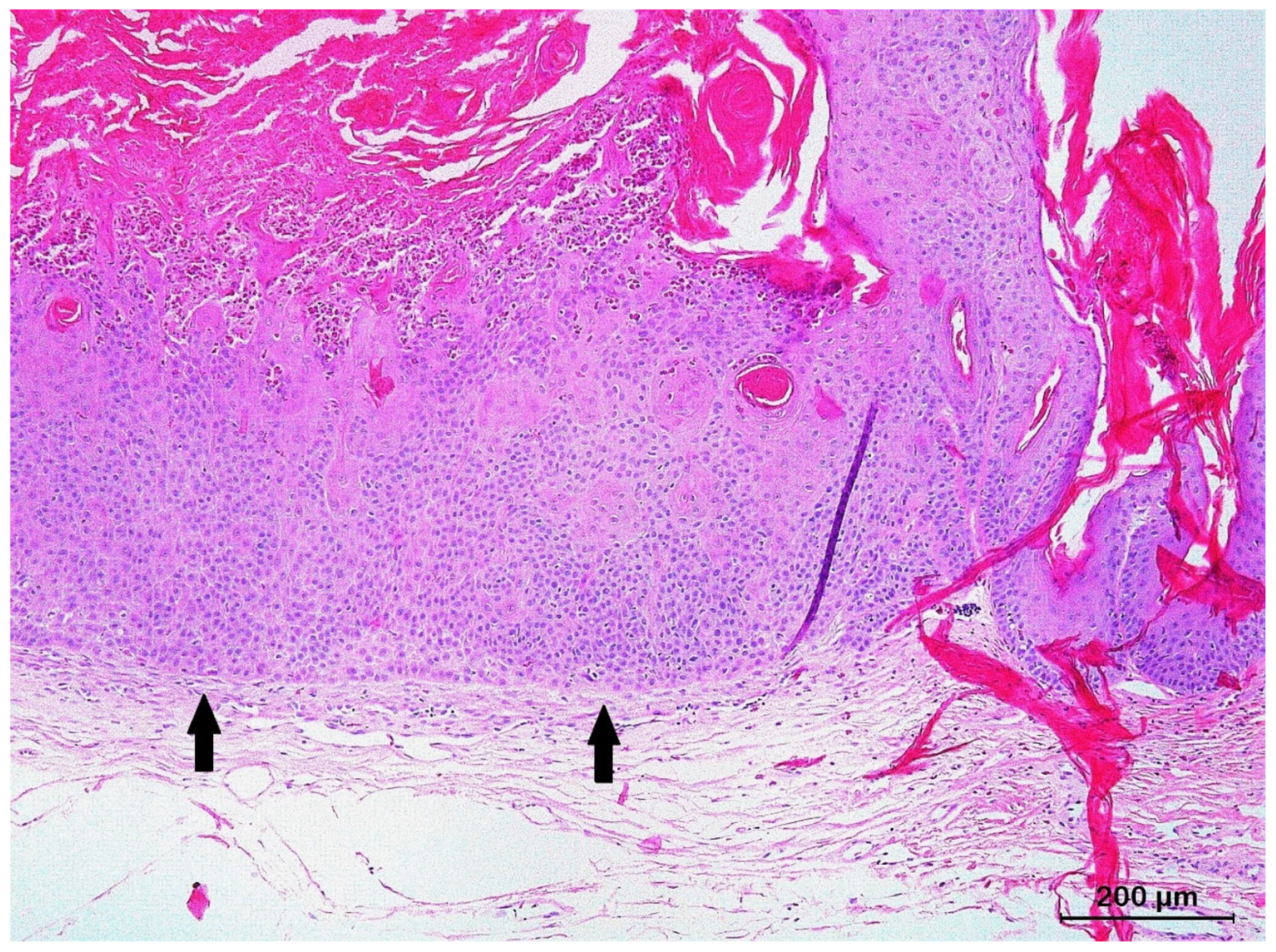
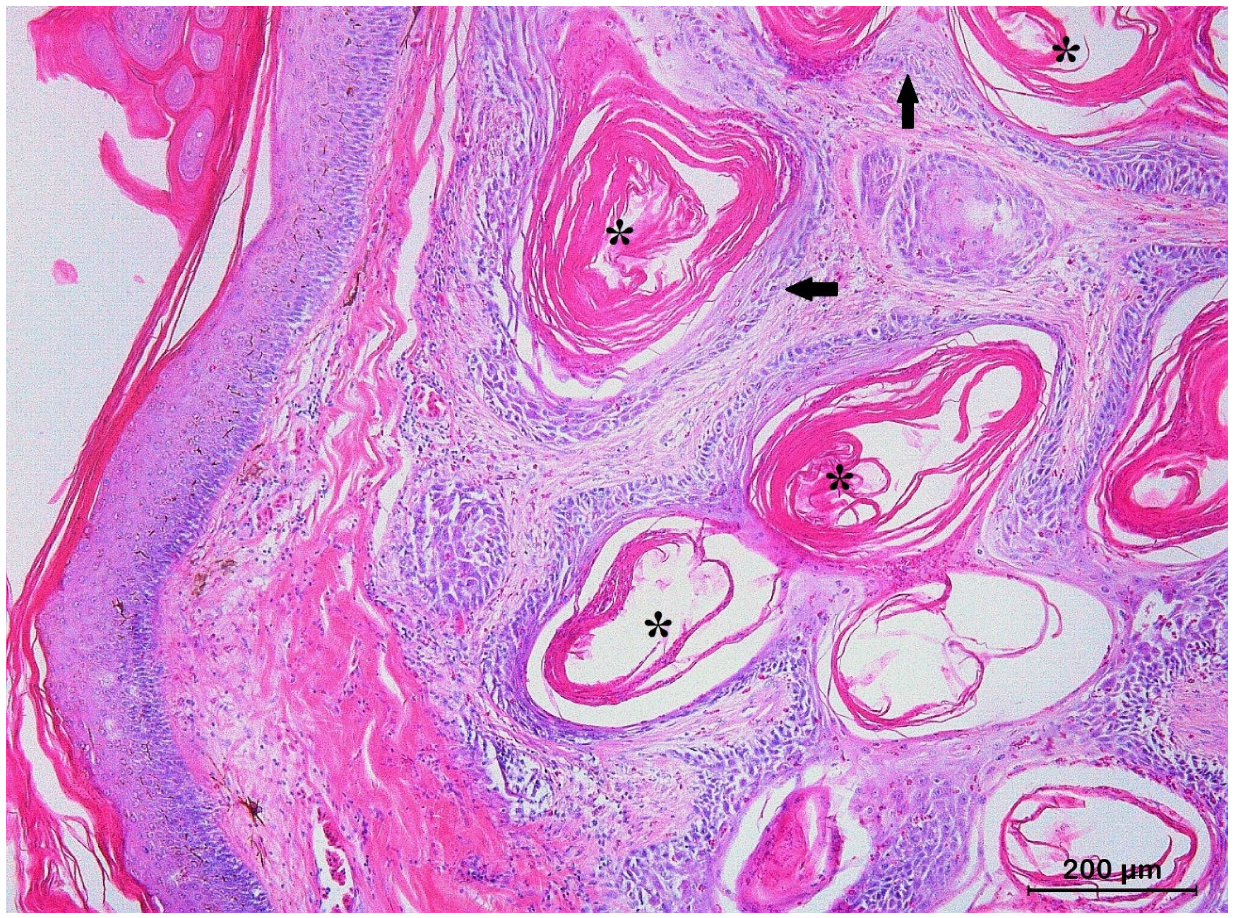
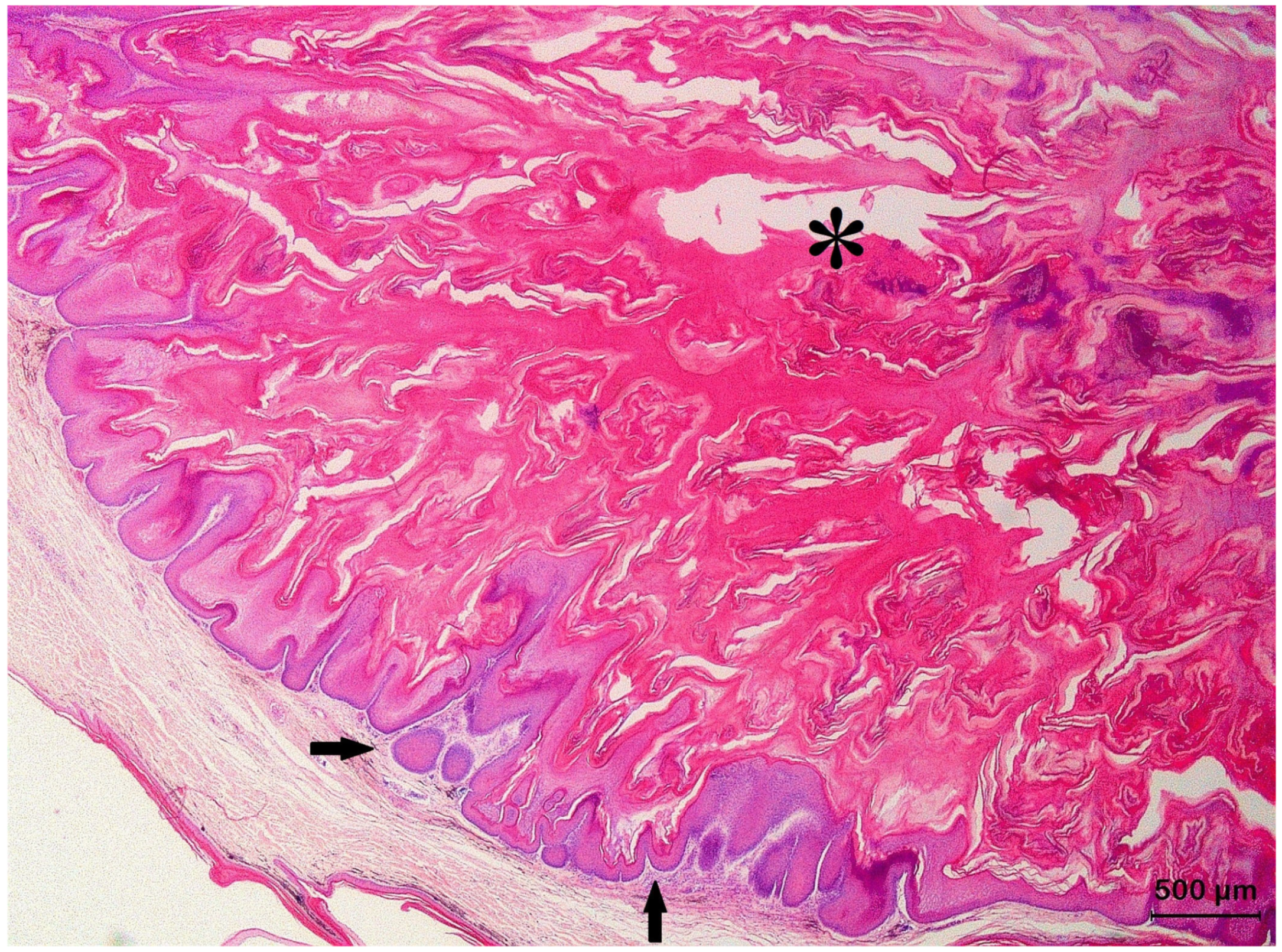
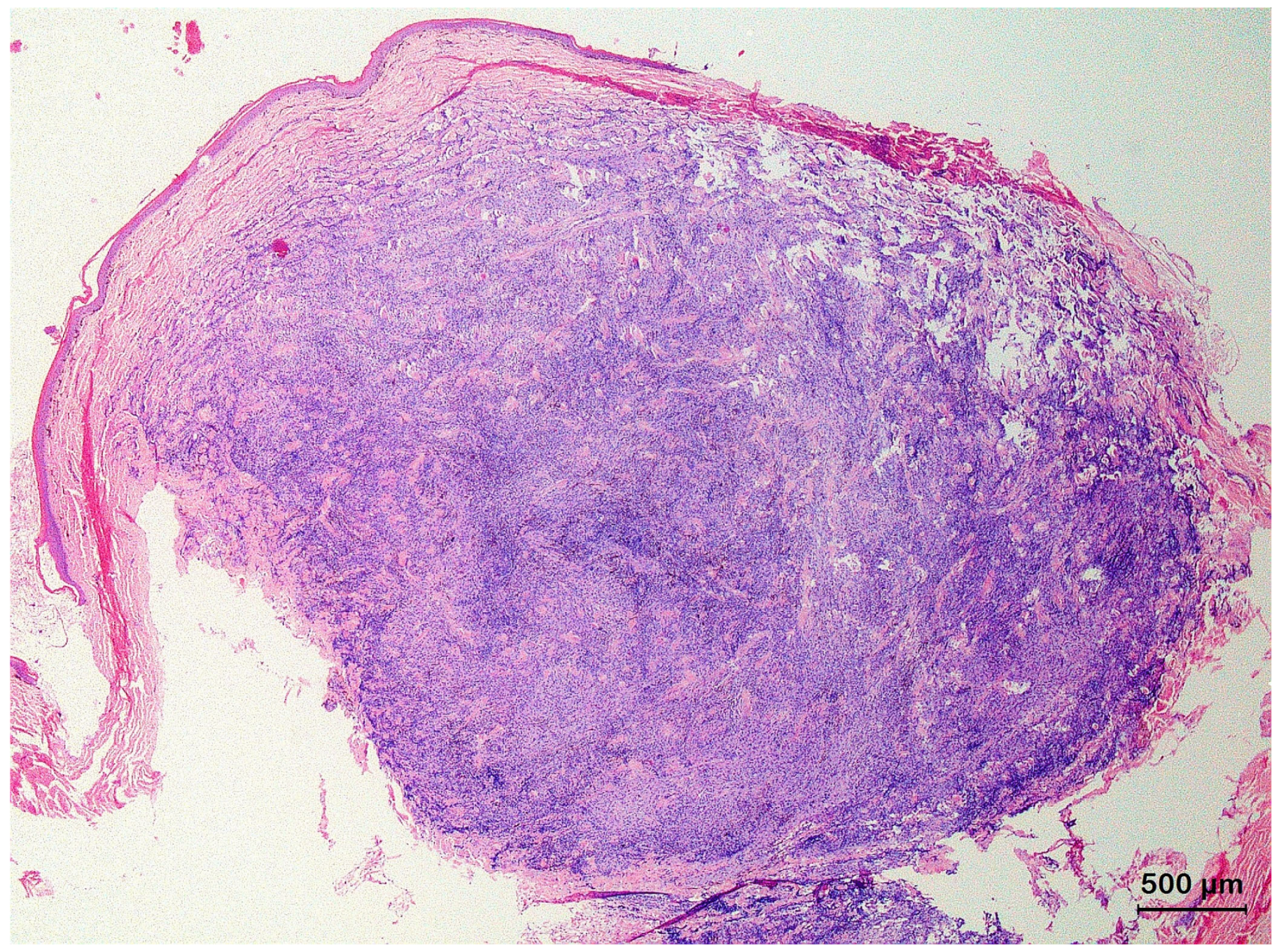
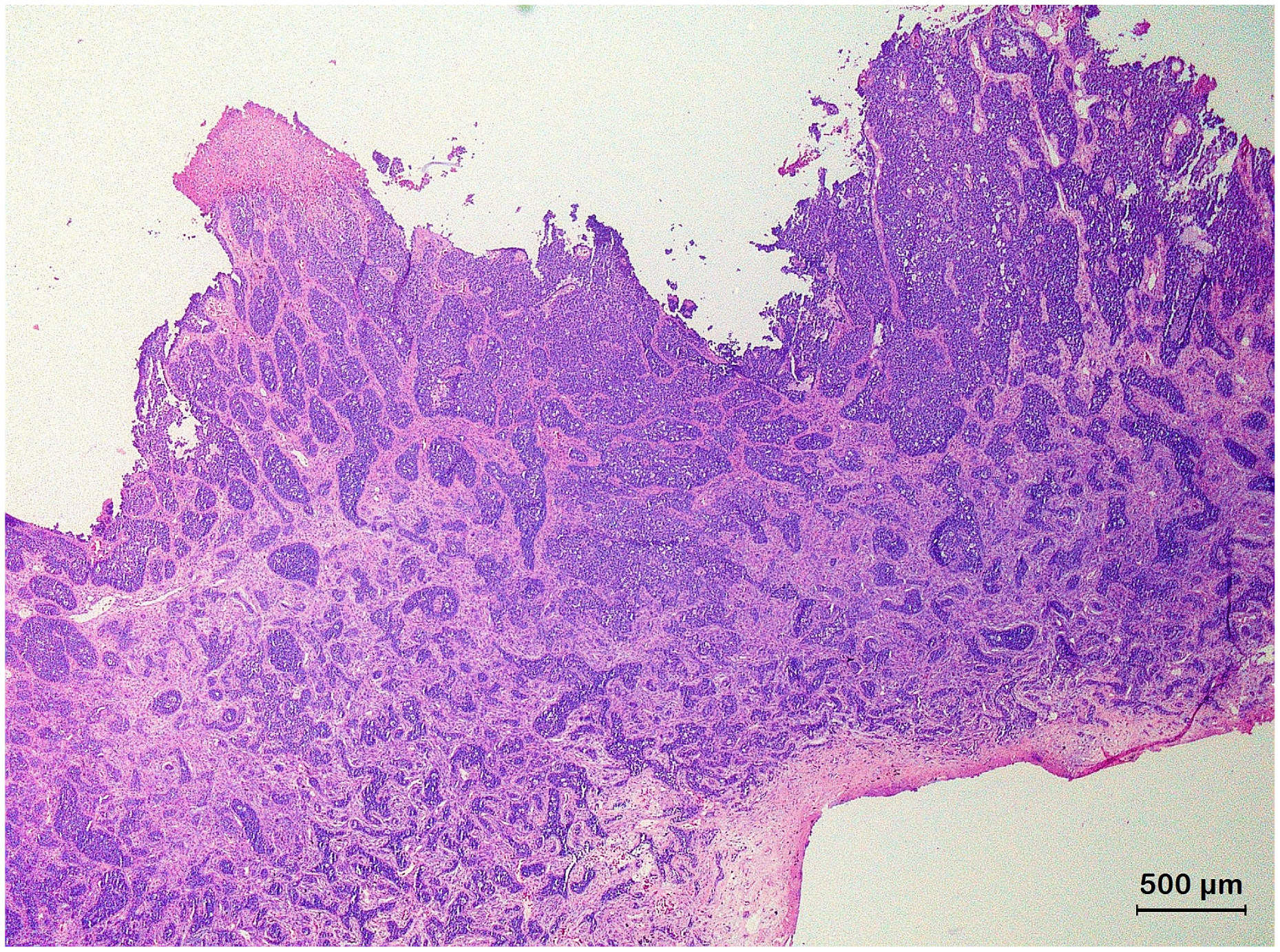
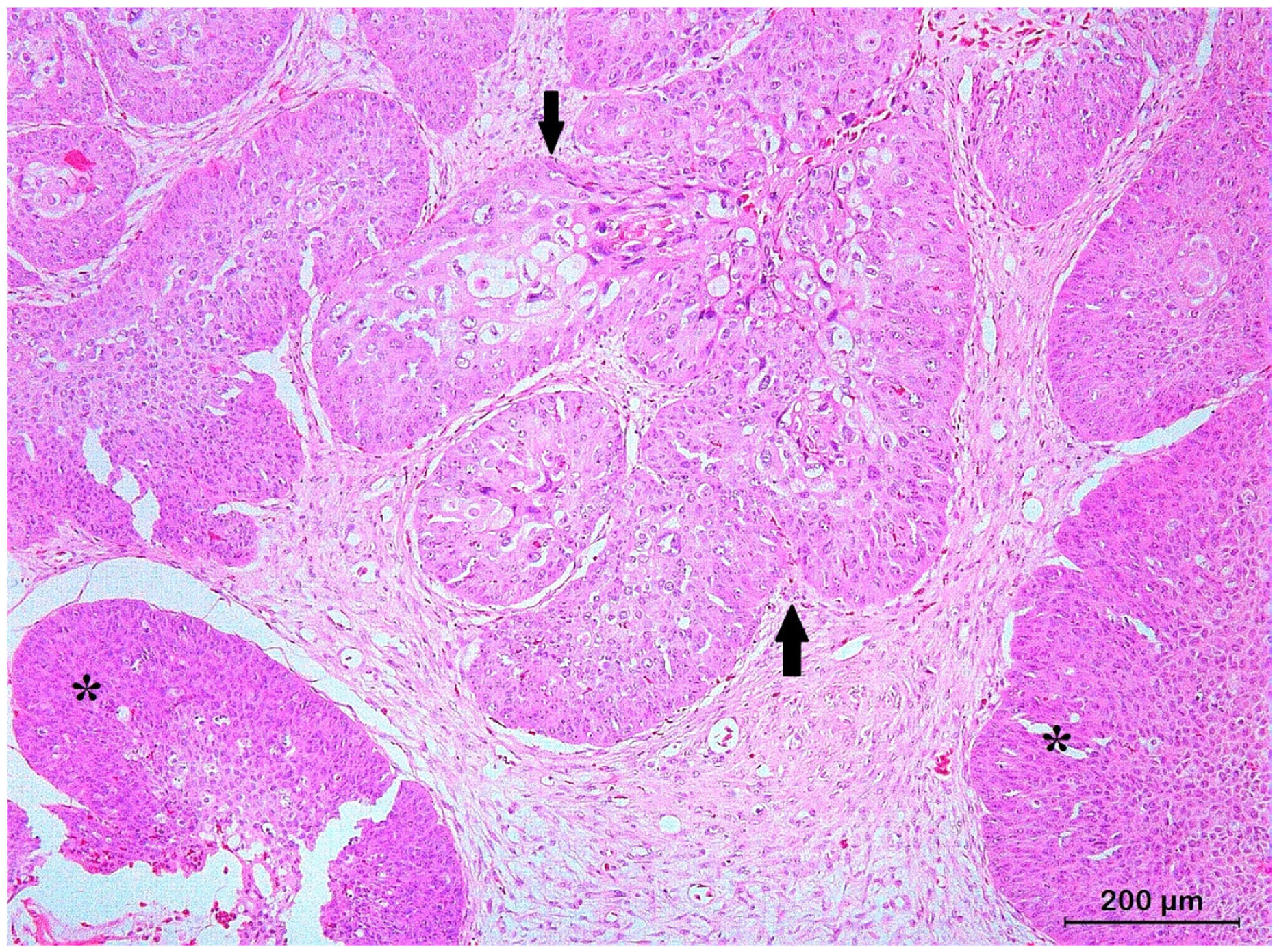
3.2. Squamous Cell Carcinoma and Its Histological Variants
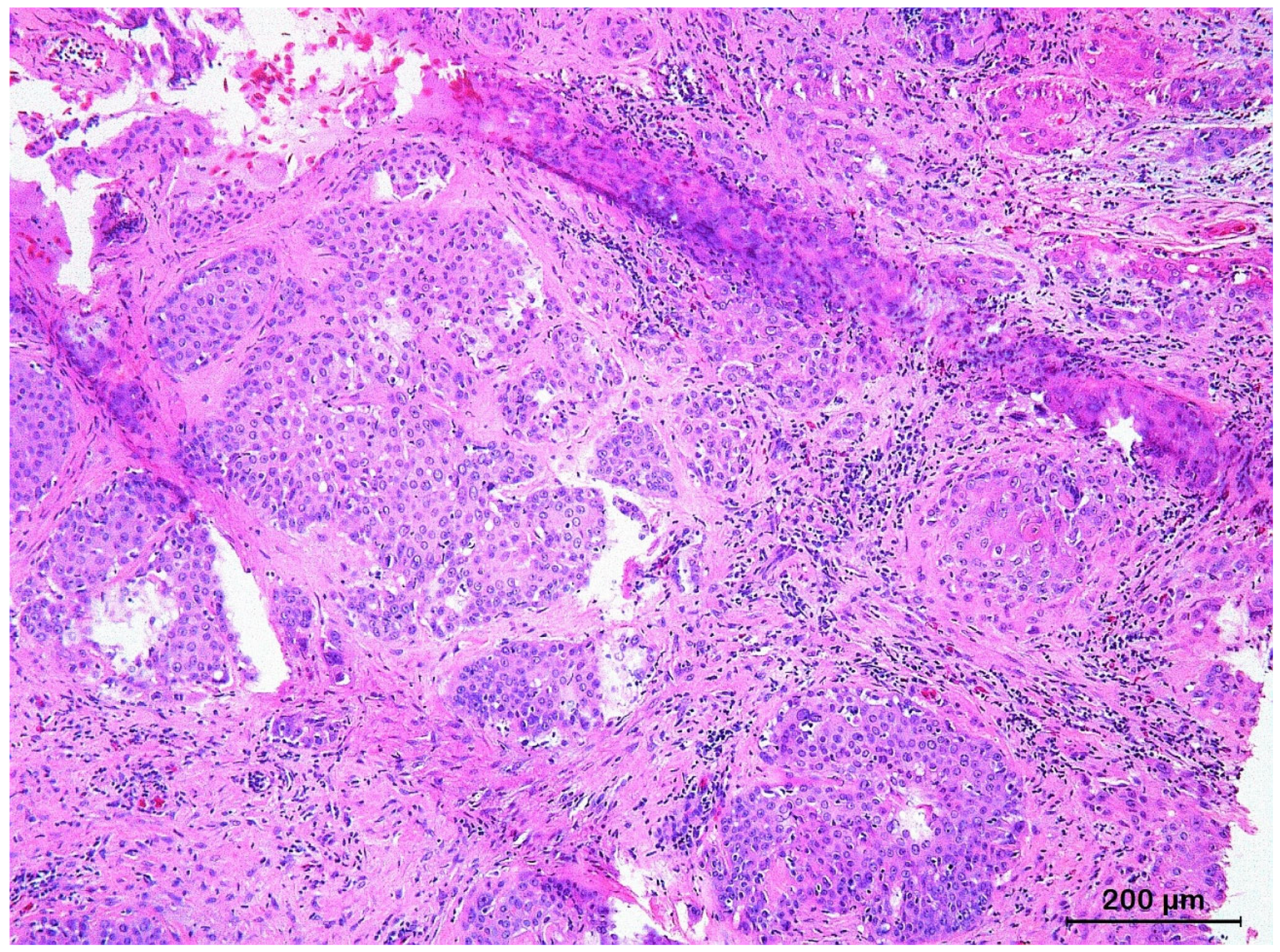
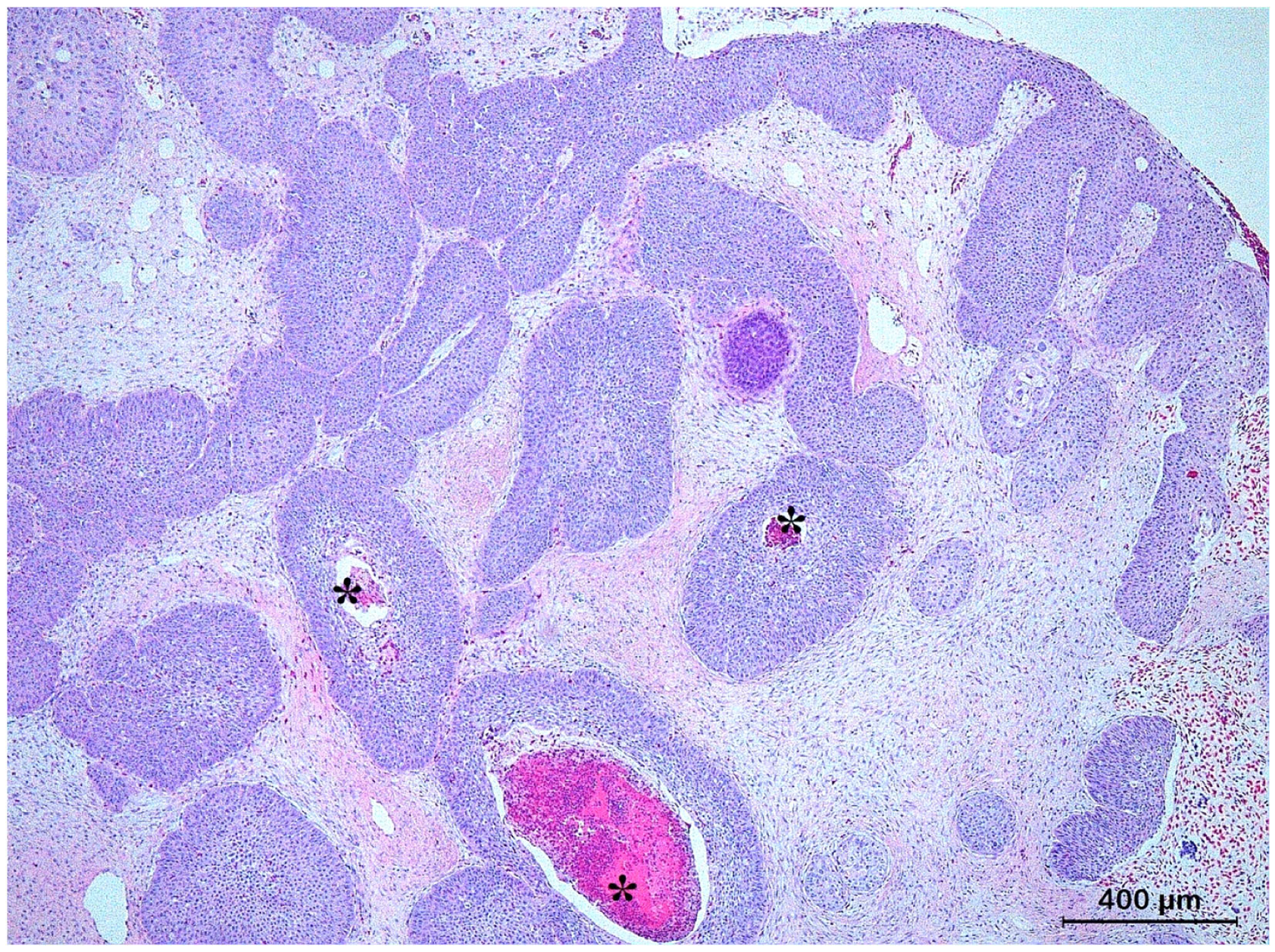
3.3. Basal Cell Carcinoma and Its Histological Variants
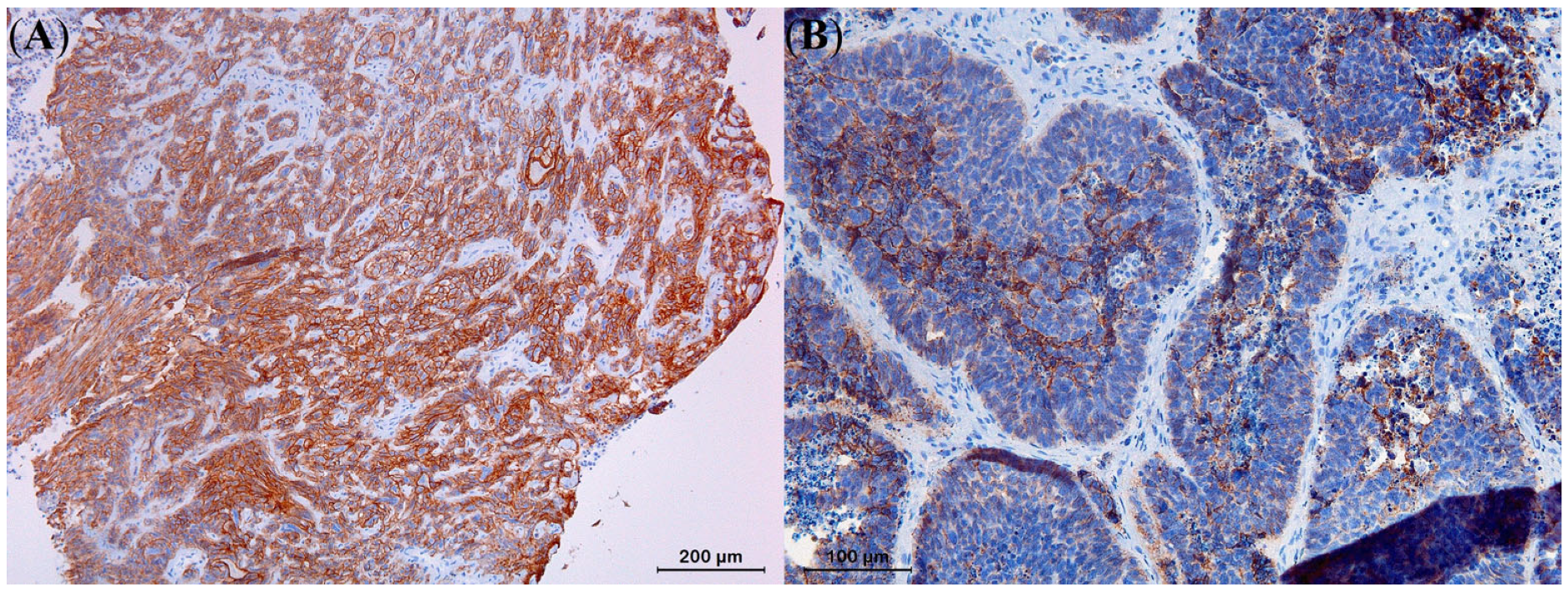
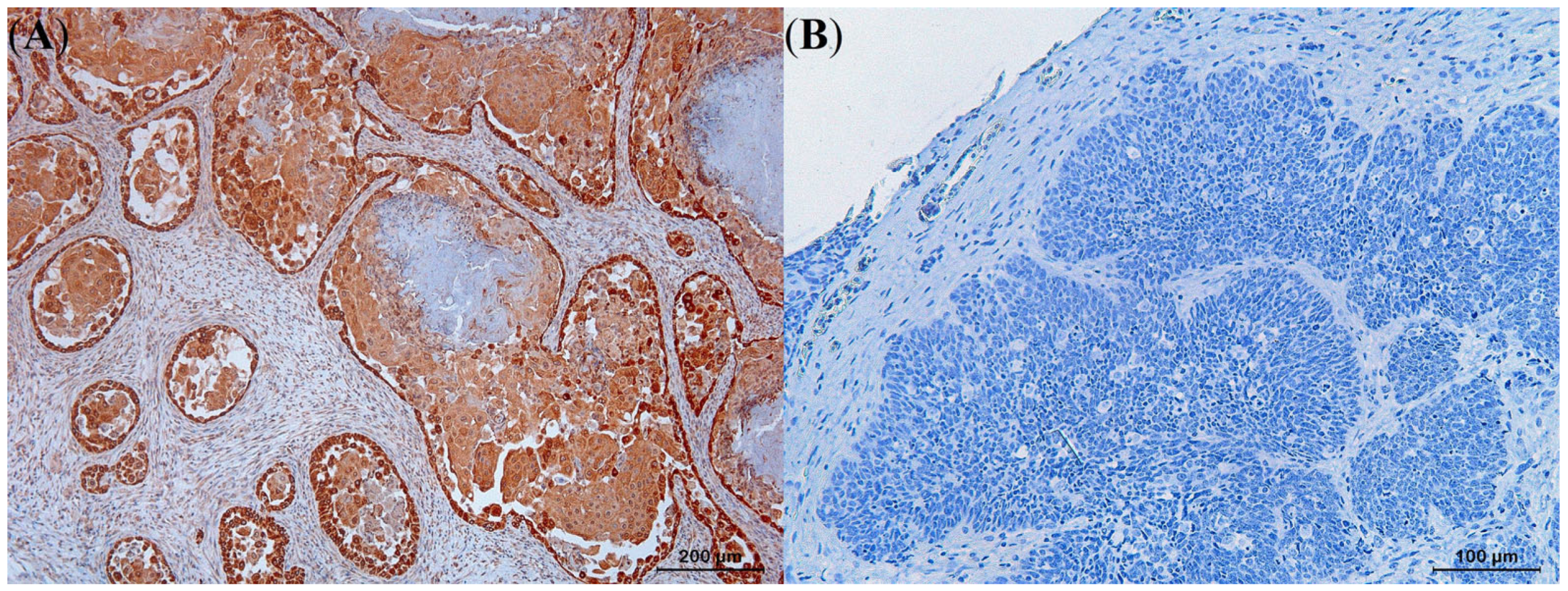
3.4. Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garner, M.M.; Hernandez-Divers, S.M.; Raymond, J.T. Reptile neoplasia: A retrospective study of case submissions to a specialty diagnostic service. Vet. Clin. N. Am. Exot. Anim. Pract. 2004, 7, 653–671. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Divers, S.M.; Garner, M.M. Neoplasia of reptiles with an emphasis on lizards. Vet. Clin. N. Am. Exot. Anim. Pract. 2003, 6, 251–273. [Google Scholar] [CrossRef] [PubMed]
- Hannon, D.E.; Garner, M.M.; Reavill, D.R. Squamous cell carcinomas in inland bearded dragons (Pogona vitticeps). J. Herpetol. Med. Surg. 2012, 21, 101–108. [Google Scholar] [CrossRef]
- Sykes, J.M.; Trupkiewicz, J.G. Reptile neoplasia at the Philadelphia zoological garden, 1901–2002. J. Zoo Wildl. Med. 2006, 37, 11–19. [Google Scholar] [CrossRef]
- Kubiak, M.; Denk, D.; Stidworthy, M.F. Retrospective review of neoplasms of captive lizards in the United Kingdom. Vet. Rec. 2020, 186, 28–37. [Google Scholar] [CrossRef]
- Christman, J.; Devau, M.; Wilson-Robles, H.; Hoppes, S.; Rech, R.; Russell, K.E.; Heatley, J.J. Oncology of reptiles: Diseases, diagnosis, and treatment. Vet. Clin. N. Am. Exot. Anim. Pract. 2017, 20, 87–110. [Google Scholar] [CrossRef]
- Heckers, K.O.; Aupperle, H.; Schmidt, V.; Pees, M. Melanophoromas and iridophoromas in reptiles. J. Comp. Pathol. 2012, 146, 258–268. [Google Scholar] [CrossRef]
- Solanes, F.; Chiers, K.; Kik, M.J.L.; Hellebuyck, T. Gross, histologic and immunohistochemical characteristics of keratoacanthomas in lizards. Animals 2023, 13, 398–413. [Google Scholar] [CrossRef]
- Monahan, C.F.; Garner, M.M.; Kiupel, M. Chromatophoromas in Reptiles. Vet. Sci. 2022, 9, 115–137. [Google Scholar] [CrossRef]
- Meyer, J.; Kolodziejek, J.; Häbich, A.C.; Dinhopl, N.; Richter, B. Multicentric Squamous Cell Tumors in Panther Chameleons (Furcifer pardalis). J. Exot. Pet. Med. 2019, 29, 166–172. [Google Scholar] [CrossRef]
- Gordon, R. Skin cancer: An overview of epidemiology and risk factors. Semin. Oncol. Nurs. 2013, 29, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Gross, T.L.; Ihrke, P.J.; Walder, E.J.; Affolter, V.K. Neoplasms and other tumors. In Skin Diseases of the Dog and Cat: Clinical and Histopathologic Diagnosis, 2nd ed.; Gross, T.L., Ihrke, P.J., Walder, E.J., Affolter, V.K., Eds.; Blackwell Science: Oxford, UK, 2005; Volume 1, pp. 581–598. [Google Scholar]
- Ho, N.T.; Smith, K.C.; Dobromylskyj, M.J. Retrospective study of more than 9000 feline cutaneous tumours in the UK: 2006–2013. J. Feline Med. Surg. 2018, 20, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Hellebuyck, T.; Ducatelle, R.; Bosseler, L.; Van Caelenberg, A.; Versnaeyen, H.; Chiers, K.; Martel, A. Basal cell carcinoma in two Hermann’s tortoises (Testudo hermanni). J. Vet. Diagn. Investig. 2016, 28, 750–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolino, G.; Donati, M.; Didona, D.; Mercuri, S.; Cantisani, C. Histology of non-melanoma skin cancers: An update. Biomedicines 2017, 5, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, K.A.; Ashack, K.A.; Khachemoune, A. Cutaneous squamous cell carcinoma: A review of high-risk and metastatic disease. Am. J. Clin. Dermatol. 2016, 17, 491–508. [Google Scholar] [CrossRef]
- Zargaran, M.; Baghaei, F. A clinical, histopathological and immunohistochemical approach to the bewildering diagnosis of keratoacanthoma. J. Dent. 2014, 15, 91–97. [Google Scholar]
- Murphy, G.; Beer, T.; Cerio, R.; Kao, G.; Nagore, E.; Pulitzer, M. Keratinocytic/epidermal tumours. In World Health Organization (WHO) Classification of Skin Tumours, 4th ed.; Elder, D., Massi, D., Scolyer, R., Willemze, R., Eds.; International Agency for Research on Cancer (IARC): Argonay, France, 2018; Volume 11, pp. 36–38. [Google Scholar]
- Vantuchová, Y.; Čuřík, R. Histological types of basal cell carcinoma. Scr. Med. Fac. Med. Univ. Brun Masaryk 2006, 79, 261–270. [Google Scholar]
- Stratigos, A.; Garbe, C.; Lebbe, C.; Malvehy, J.; Del Marmol, V.; Pehamberger, H.; Peris, K.; Becker, J.C.; Zalaudek, I.; Saiag, P.; et al. Diagnosis and treatment of invasive squamous cell carcinoma of the skin: European consensus-based interdisciplinary guideline. Eur. J. Cancer 2015, 51, 1989–2007. [Google Scholar] [CrossRef]
- Zehnder, A.M.; Swift, L.A.; Sundaram, A.; Speer, B.L.; Olsen, G.P.; Hawkins, M.G.; Paul-Murphy, J. Clinical features, treatment, and outcomes of cutaneous and oral squamous cell carcinoma in avian species. J. Am. Vet. Med. Assoc. 2018, 252, 309–315. [Google Scholar] [CrossRef]
- Orós, J.; López-Yánez, M.; Rodríguez, F.; Calabuig, P.; Castro, P.L. Immunohistochemical staining patterns of alpha-keratins in normal tissues from two reptile species: Implications for characterization of squamous cell carcinomas. BMC Vet. Res. 2018, 14, 219. [Google Scholar] [CrossRef]
- Erdogan Bamac, O.; Seckin Arun, S. Histological and immunohistochemical evaluation of epithelial and mesenchymal tumors of psittacines. Med. Weter. 2020, 76, 165–169. [Google Scholar]
- Jones, A.L.; Suárez-Bonnet, A.; Mitchell, J.A.; Ramirez, G.A.; Stidworthy, M.F.; Priestnall, S.L. Avian papilloma and squamous cell carcinoma: A histopathological, immunohistochemical and virological study. J. Comp. Pathol. 2020, 175, 13–23. [Google Scholar] [CrossRef]
- Beer, T.W.; Shepherd, P.; Theaker, J.M. Ber EP4 and epithelial membrane antigen aid distinction of basal cell, squamous cell and basosquamous carcinomas of the skin. Histopathology 2000, 37, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, M.; Zavattaro, E.; Sadeghi, M. Immunohistochemistry expression of EMA, CD10, CEA, and Bcl-2 in distinguishing cutaneous basal cell from squamous cell carcinoma: A systematic review. Gulhane Med. J. 2020, 62, 63–71. [Google Scholar] [CrossRef]
- Alhumaidi, A. Practical immunohistochemistry of epithelial skin tumor. Indian J. Dermatol. Venereol. Leprol. 2012, 78, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Compton, L.A.; Murphy, G.F.; Lian, C.G. Diagnostic immunohistochemistry in cutaneous neoplasia: An update. Dermatopathology 2015, 2, 15–42. [Google Scholar] [CrossRef]
- Nagamine, E.; Hirayama, K.; Matsuda, K.; Okamoto, M.; Ohmachi, T.; Uchida, K.; Kadosawa, T.; Taniyama, H. Invasive front grading and epithelial-mesenchymal transition in canine oral and cutaneous squamous cell carcinomas. Vet. Pathol. 2017, 54, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Thaiwong, T.; Sledge, D.G.; Collins-Webb, A.; Kiupel, M. Immunohistochemical characterization of canine oral papillary squamous cell carcinoma. Vet. Pathol. 2018, 55, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Sanz Ressel, B.L.; Massone, A.R.; Barbeito, C.G. Immunohistochemical expression of selected phosphoproteins of the mTOR signalling pathway in canine cutaneous squamous cell carcinoma. Vet. J. 2019, 245, 41–48. [Google Scholar] [CrossRef]
- Gál, J.; Mándoki, M. Adenoma of the cloacal scent gland in a California Kingsnake (Lampropeltis getulus californiae). Acta Vet. Hung. 2012, 60, 459–463. [Google Scholar] [CrossRef]
- Petterino, C.; Bedin, M.; Podestà, G.; Ratto, A. Undifferentiated tumor in the ovary of a corn snake (Elaphe guttata guttata). Vet. Clin. Pathol. 2006, 35, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.M.; Garner, M.M.; Chilton, J.A.; Jacobson, E.R.; Kiupel, M. Gastric neuroendocrine carcinomas in bearded dragons (Pogona vitticeps). Vet. Pathol. 2009, 46, 1109–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargo, N. Basal cell and squamous cell carcinoma. Semin. Oncol. Nurs. 2003, 19, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—A review. Diagn. Pathol. 2014, 9, 221–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldschmidt, M.H.; Goldschmidt, K.H. Epithelial and melanocytic tumors of the skin. In Tumors in Domestic Animals, 5th ed.; Meuten, D.J., Ed.; John Wiley & Sons: Oxford, UK, 2017; Volume 1, pp. 88–148. [Google Scholar]
- Misago, N.; Inoue, T.; Koba, S.; Narisawa, Y. Keratoacanthoma and other types of squamous cell carcinoma with crateriform architecture: Classification and identification. J. Dermatol. 2013, 40, 443–452. [Google Scholar] [CrossRef]
- Yanofsky, V.R.; Mercer, S.E.; Phelps, R.G. Histopathological variants of cutaneous squamous cell carcinoma: A review. J. Skin Cancer 2011, 2011, 210813. [Google Scholar] [CrossRef] [Green Version]
- Putti, T.C.; Teh, M.; Lee, Y.S. Biological behavior of keratoacanthoma and squamous cell carcinoma: Telomerase activity and COX-2 as potential markers. Mod. Pathol. 2004, 17, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Alibardi, L.; Dalla Valle, L.; Toffolo, V.; Toni, M. Scale keratin in lizard epidermis reveals amino acid regions homologous with avian and mammalian epidermal proteins. Anat. Rec. 2006, 288, 734–752. [Google Scholar] [CrossRef]
- Alibardi, L.; Toni, M. Immunolocalization and characterization of beta-keratins in growing epidermis of chelonians. Tissue Cell 2006, 38, 53–63. [Google Scholar] [CrossRef]
- Marzuka, A.G.; Book, S.E. Basal cell carcinoma: Pathogenesis, epidemiology, clinical features, diagnosis, histopathology, and management. Yale J. Biol. Med. 2015, 88, 167–179. [Google Scholar]
- Rajabi, H.; Kufe, D. MUC1-C oncoprotein integrates a program of EMT, epigenetic reprogramming and immune evasion in human carcinomas. Biochim. Biophys. Acta 2017, 1868, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, M.; Mohamadzaheri, E.; Khazaei, S.; Najafi, F.; Vaisi-Raygani, A.; Rahbar, M.; Sadeghi, M. Comparison of EMA, CEA, CD10 and Bcl-2 biomarkers by immunohistochemistry in squamous cell carcinoma and basal cell carcinoma of the skin. Asian Pac. J. Cancer Prev. 2016, 17, 1379–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordi, J.; Romagosa, C.; Tavassoli, F.A.; Nogales, F.; Palacin, A.; Condom, E.; Torné, A.; Cardesa, A. CD10 expression in epithelial tissues and tumors of the gynecologic tract: A useful marker in the diagnosis of mesonephric, trophoblastic, and clear cell tumors. Am. J. Surg. Pathol. 2003, 27, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Peat, T.J.; Edmondson, E.F.; Miller, M.A.; DuSold, D.M.; Ramos-Vara, J.A. Pax8, Napsin A, and CD10 as immunohistochemical markers of canine renal cell carcinoma. Vet. Pathol. 2017, 54, 588–594. [Google Scholar] [CrossRef]
- Aouacheria, A.; Navratil, V.; Combet, C. Database and bioinformatic analysis of Bcl-2 family proteins and BH3-Only proteins. Methods Mol. Biol. 2019, 1877, 23–43. [Google Scholar]
- Tsujii, M.; Kawano, S.; Dubois, R.N. Cyclooxygenase-2 expression in human colon cancer cells increases metastatic potential. Proc. Natl. Acad. Sci. USA 1997, 94, 3336–3340. [Google Scholar] [CrossRef] [Green Version]
- Karagece Yalçin, Ü.; Seçkin, S. The expression of p53 and Cox-2 in basal cell carcinoma, squamous cell carcinoma and actinic keratosis cases. Turkish J. Pathol. 2012, 28, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Lyakhovitsky, A.; Barzilai, A.; Fogel, M.; Trau, H.; Huszar, M. Expression of E-cadherin and beta-catenin in cutaneous squamous cell carcinoma and its precursors. Am. J. Dermatopathol. 2004, 26, 372–378. [Google Scholar] [CrossRef]
- Papadavid, E.; Pignatelli, M.; Zakynthinos, S.; Krausz, T.; Chu, A.C. The potential role of abnormal E-cadherin and a-, b- and g -catenin immunoreactivity in the determination of the biological behaviour of keratoacanthoma. Br. J. Dermatol. 2001, 145, 582–589. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Initial Diagnosis | Re-Classification | |
|---|---|---|---|---|
| Final Diagnosis | Histological Variant | |||
| Bearded dragon (Pogona vitticeps) (n = 9) | Skin | 6 SCC | 4 SCC | 3 WD SCC |
| 1 KA | ||||
| 1 BCC | 1 Infiltrating BCC | |||
| 1 Non-neoplastic | Gingival fibrous hyperplasia | |||
| MCJ | 2 SCC | 2 SCC | 1 WD SCC | |
| 1 MD SCC | ||||
| Oral | 1 BCC | 1 BCC | 1 Solid BCC | |
| Panther chameleon (Furcifer pardalis) (n = 5) | Skin | 4 SCC | 4 SCC | 4 KA |
| MCJ | 1 SCC | 1 SCC | 1 SCC in situ | |
| Veiled chameleon (Chamaeleo calyptratus) (n = 2) | Skin | 1 SCC | 1 SCC | 1 KA |
| MCJ | 1 SCC | 1 BCC | 1 Keratotic BCC | |
| Brown anole (Anolis sagrei) (n = 2) | Skin | 2 SCC | 1 SCC | 1 WD SCC |
| 1 BCC | 1 Keratotic BCC | |||
| Common blue-tongued skink (Tiliqua scincoides) (n = 1) | Skin | 1 SCC | 1 Non-neoplastic | Cystic mass lined by pseudostratified ciliated epithelium |
| Green iguana (Iguana iguana) (n = 1) | Skin | 1 SCC | 1 BCC | 1 Keratotic BCC |
| Von Höhnel’s chameleon (Trioceros hoehnelii) (n = 1) | Skin | 1 SCC | 1 BCC | 1 Keratotic BCC |
| Boa constrictor (Boa constrictor) (n = 1) | Skin | 1 SCC | 1 BCC | 1 Keratotic BCC |
| False map turtle (Graptemys pseudogeographica) (n = 1) | Skin | 1 SCC | 1 Non-neoplastic | Pyogranulomatous dermatitis |
| Yellow-belied slider (Trachemys scripta scripta) (n = 1) | Skin | 1 SCC | 1 SCC | 1 WD SCC |
| Hermann’s tortoise (Testudo hermanni) (n = 9) | Skin | 1 SCC | 1 BCC | 1 Infiltrating BCC |
| Shell | 2 SCC | 1 SCC | 1 WD SCC | |
| 3 BCC | 4 BCC | 1 Solid BCC | ||
| 2 Infiltrating BCC | ||||
| 1 BSCC | ||||
| MCJ | 1 SCC | 1 SCC | 1 MD SCC | |
| Oral | 2 BCC | 2 BCC | 2 Solid BCC | |
| African spurred tortoise (Centrochelys sulcata) (n = 1) | Oral | 1 SCC | 1 SCC | 1 MD SCC |
| Red-eared slider (T. scripta elegans) (n = 1) | Skin | 1 BCC | 1 BCC | 1 Solid BCC |
| A (Percentage of Positive Cells) | B (Intensity of Staining) | IRS Score (A × B) |
|---|---|---|
| 0 = no positive cells | 0 = no color reaction | 0–1 = negative expression |
| 1 ≤ 10% positive cells | 1 = mild reaction | 2–3 = poor expression |
| 2 = 10–50% positive cells | 2 = moderate reaction | 4–8 = moderate expression |
| 3 = 51–80% positive cells | 3 = intense reaction | 9–12 = strong expression |
| 4 ≥ 80% positive cells | Final IRS score (A × B): 0–12 1 | |
| Neoplasm | Total | Mitotic Index | Degree of Nuclear Atypia | ||||
|---|---|---|---|---|---|---|---|
| 0–2 | 3–4 | ≥5 | <30% | 30–60% | >60% | ||
SCC
| 17 | 15 (88.2%) | 1 (5.9%) | 1 (5.9%) | 11 (64.7%) | 4 (23.5%) | 2 (11.8%) |
| 1 | 0 | 0 | 1 | 1 | 0 | 0 | |
| 7 | 6 | 1 | 0 | 3 | 4 | 0 | |
| 3 | 3 | 0 | 0 | 1 | 0 | 2 | |
| 6 | 6 | 0 | 0 | 6 | 0 | 0 | |
BCC
| 15 | 7 (46.7%) | 7 (46.7%) | 1 (6.7%) | 8 (53.3%) | 3 (20%) | 4 (26.7%) |
| 5 | 4 | 1 | 0 | 4 | 0 | 1 | |
| 5 | 2 | 2 | 1 | 4 | 1 | 0 | |
| 4 | 1 | 3 | 0 | 0 | 2 | 2 | |
| 1 | 0 | 1 | 0 | 0 | 0 | 1 | |
| Neoplasm | Total | E-Cadherin (IRS Score) (%) | COX-2 (IRS Score) (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Neg | Poor | Mod | Strong | Neg | Poor | Mod | Strong | |||
| SCC | 17 | 0 (0%) | 0 (0%) | 5 (29.4%) | 12 (70.6%) | 0 (0%) | 0 (0%) | 10 (58.8%) | 7 (41.2%) | |
| 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | |
| ||||||||||
| 7 | 0 | 0 | 5 | 2 | 0 | 0 | 3 | 4 | |
| 3 | 0 | 0 | 0 | 3 | 0 | 0 | 1 | 2 | |
| 6 | 0 | 0 | 0 | 6 | 0 | 0 | 6 | 0 | |
| BCC | 15 | 0 (0%) | 3 (20%) | 12 (80%) | 0 (0%) | 7 (46.7%) | 4 (26.7%) | 4 (26.7%) | 0 (0%) | |
| 5 | 0 | 1 | 4 | 0 | 4 | 1 | 0 | 0 | |
| 5 | 0 | 1 | 4 | 0 | 0 | 1 | 4 | 0 | |
| 4 | 0 | 1 | 3 | 0 | 3 | 1 | 0 | 0 | |
| 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solanes Vilanova, F.; Hellebuyck, T.; Chiers, K. Histological Variants of Squamous and Basal Cell Carcinoma in Squamates and Chelonians: A Comprehensive Classification. Animals 2023, 13, 1327. https://doi.org/10.3390/ani13081327
Solanes Vilanova F, Hellebuyck T, Chiers K. Histological Variants of Squamous and Basal Cell Carcinoma in Squamates and Chelonians: A Comprehensive Classification. Animals. 2023; 13(8):1327. https://doi.org/10.3390/ani13081327
Chicago/Turabian StyleSolanes Vilanova, Ferran, Tom Hellebuyck, and Koen Chiers. 2023. "Histological Variants of Squamous and Basal Cell Carcinoma in Squamates and Chelonians: A Comprehensive Classification" Animals 13, no. 8: 1327. https://doi.org/10.3390/ani13081327