
Characterization of Bacillus pumilus Strains Isolated from Bovine Uteri

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Culture and Growth Kinetics
2.2. Keratinase Gene PCR
2.3. Primary Endometrial Epithelial Cell Culture
2.4. Viability Assay
2.5. Statistical Analysis
3. Results
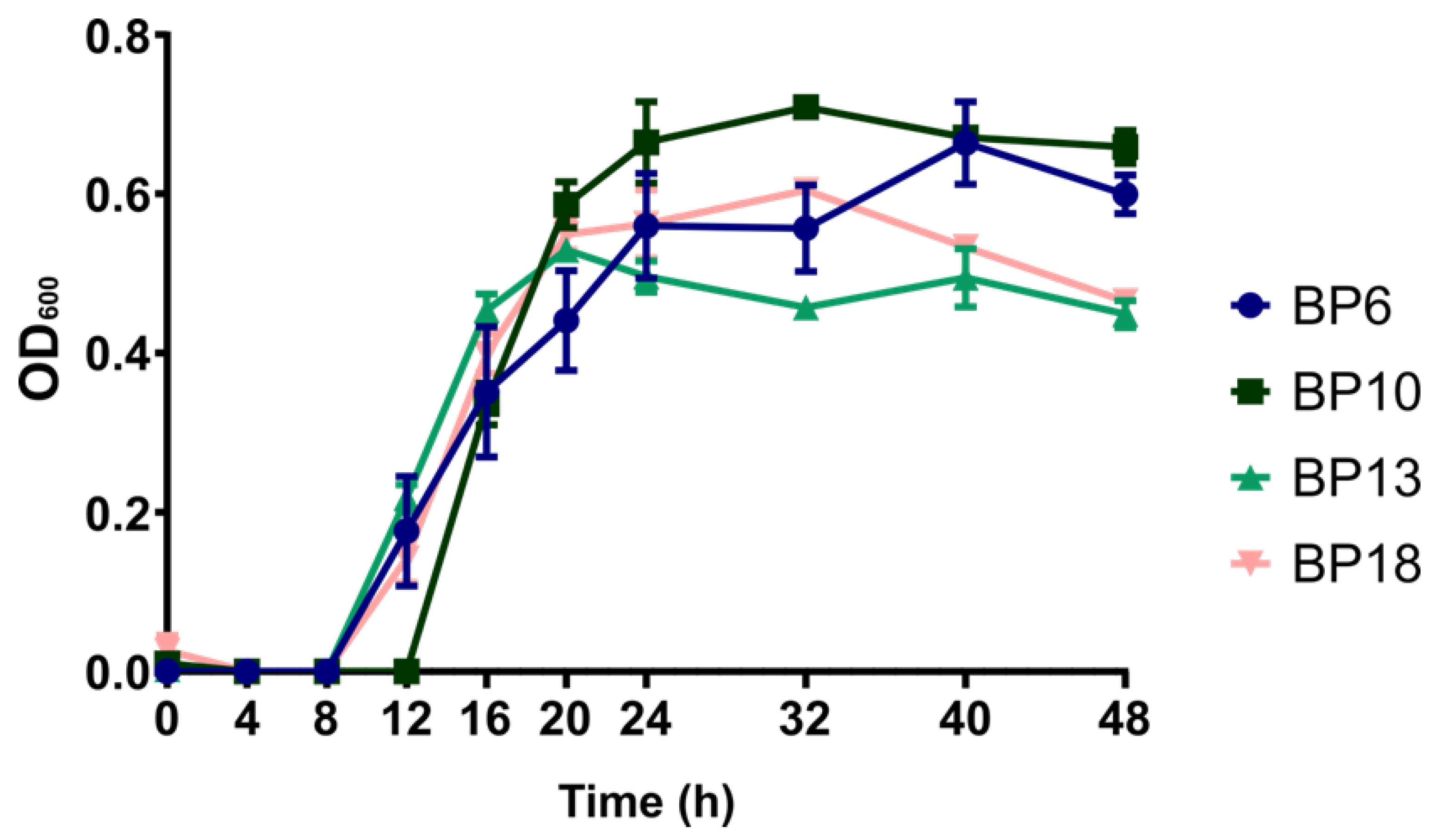
3.1. Keratinase Gene Screening, and Growth Kinetics
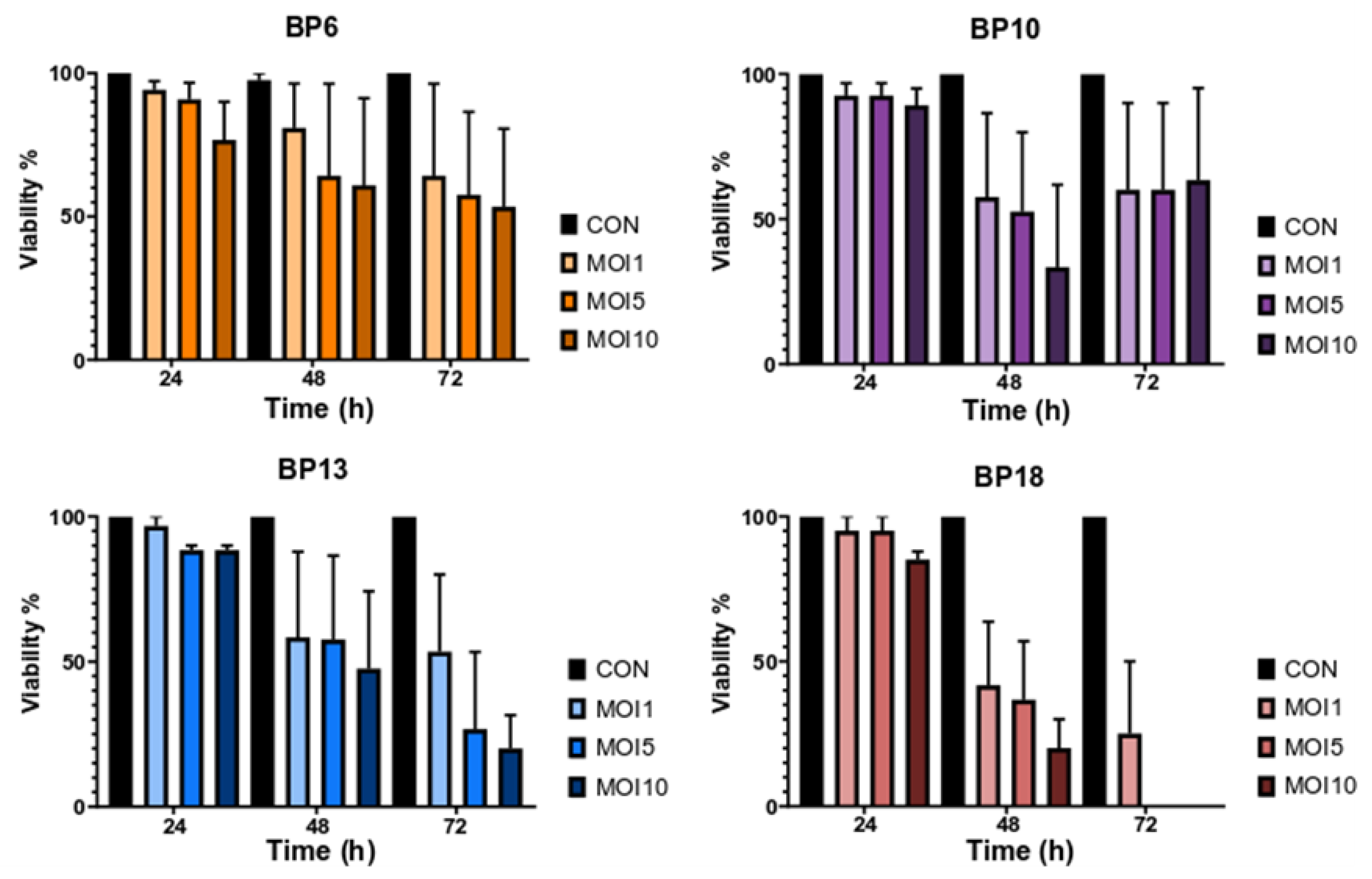
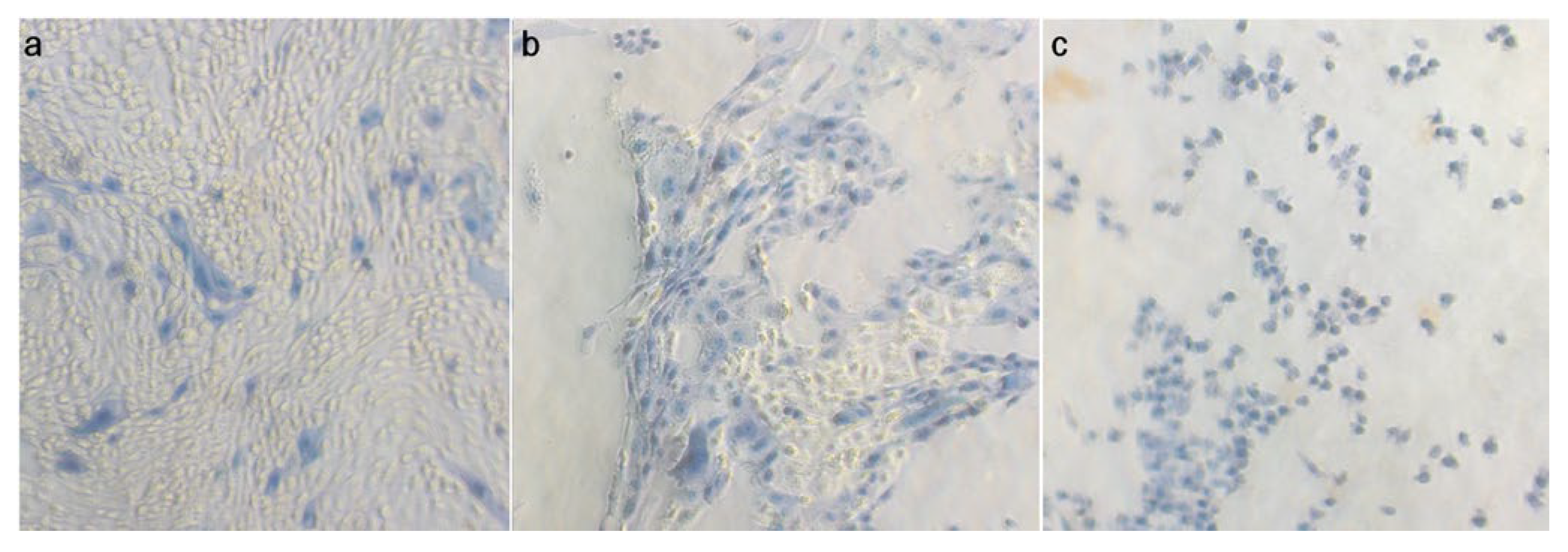
3.2. Effect of B. pumilus Isolates on Uterine Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheldon, I.M.; Molinari, P.C.C.; Ormsby, T.J.R.; Bromfield, J.J. Preventing Postpartum Uterine Disease in Dairy Cattle Depends on Avoiding, Tolerating and Resisting Pathogenic Bacteria. Theriogenology 2020, 150, 158–165. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Noakes, D.E.; Rycroft, A.N.; Pfeiffer, D.U.; Dobson, H. Influence of Uterine Bacterial Contamination after Parturition on Ovarian Dominant Follicle Selection and Follicle Growth and Function in Cattle. Reproduction 2002, 123, 837–845. [Google Scholar] [CrossRef]
- Moore, S.G.; Ericsson, A.C.; Poock, S.E.; Melendez, P.; Lucy, M.C. Hot Topic: 16S rRNA Gene Sequencing Reveals the Microbiome of the Virgin and Pregnant Bovine Uterus. J. Dairy Sci. 2017, 100, 4953–4960. [Google Scholar] [CrossRef] [Green Version]
- Ballas, P.; Reinländer, U.; Schlegl, R.; Ehling-Schulz, M.; Drillich, M.; Wagener, K. Characterization of Intrauterine Cultivable Aerobic Microbiota at the Time of Insemination in Dairy Cows with and without Mild Endometritis. Theriogenology 2021, 159, 28–34. [Google Scholar] [CrossRef]
- Santos, T.M.A.; Gilbert, R.O.; Bicalho, R.C. Metagenomic Analysis of the Uterine Bacterial Microbiota in Healthy and Metritic Postpartum Dairy Cows. J. Dairy Sci. 2011, 94, 291–302. [Google Scholar] [CrossRef]
- Prunner, I.; Pothmann, H.; Wagener, K.; Giuliodori, M.; Huber, J.; Ehling-Schulz, M.; Drillich, M. Dynamics of Bacteriologic and Cytologic Changes in the Uterus of Postpartum Dairy Cows. Theriogenology 2014, 82, 1316–1322. [Google Scholar] [CrossRef]
- Jeon, S.J.; Vieira-Neto, A.; Gobikrushanth, M.; Daetz, R.; Mingoti, R.D.; Parize, A.C.B.; de Freitas, S.L.; da Costa, A.N.L.; Bicalho, R.C.; Lima, S.; et al. Uterine Microbiota Progression from Calving until Establishment of Metritis in Dairy Cows. Appl. Environ. Microbiol. 2015, 81, 6324–6332. [Google Scholar] [CrossRef] [Green Version]
- Wagener, K.; Prunner, I.; Pothmann, H.; Drillich, M.; Ehling-Schulz, M. Diversity and Health Status Specific Fluctuations of Intrauterine Microbial Communities in Postpartum Dairy Cows. Vet. Microbiol. 2015, 175, 286–293. [Google Scholar] [CrossRef]
- Bicalho, M.L.S.; Lima, S.; Higgins, C.H.; Machado, V.S.; Lima, F.S.; Bicalho, R.C. Genetic and Functional Analysis of the Bovine Uterine Microbiota. Part II: Purulent Vaginal Discharge versus Healthy Cows. J. Dairy Sci. 2017, 100, 3863–3874. [Google Scholar] [CrossRef] [Green Version]
- Paiano, R.B.; Moreno, L.Z.; Gomes, V.T.M.; Parra, B.M.; Barbosa, M.R.; Sato, M.I.Z.; Bonilla, J.; Pugliesi, G.; Baruselli, P.S.; Moreno, A.M. Assessment of the Main Pathogens Associated with Clinical and Subclinical Endometritis in Cows by Culture and MALDI-TOF Mass Spectrometry Identification. J. Dairy Sci. 2022, 105, 3367–3376. [Google Scholar] [CrossRef]
- Wang, M.L.; Liu, M.C.; Xu, J.; An, L.G.; Wang, J.F.; Zhu, Y.H. Uterine Microbiota of Dairy Cows with Clinical and Subclinical Endometritis. Front. Microbiol. 2018, 9, 2691. [Google Scholar] [CrossRef] [PubMed]
- Pascottini, O.B.; Van Schyndel, S.J.; Spricigo, J.F.W.; Rousseau, J.; Weese, J.S.; LeBlanc, S.J. Dynamics of Uterine Microbiota in Postpartum Dairy Cows with Clinical or Subclinical Endometritis. Sci. Rep. 2020, 10, 12353. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, L.C.; Cronin, J.G.; Sheldon, I.M. Mechanisms Linking Bacterial Infections of the Bovine Endometrium to Disease and Infertility. Reprod. Biol. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicalho, R.C.; Machado, V.S.; Bicalho, M.L.S.; Gilbert, R.O.; Teixeira, A.G.V.; Caixeta, L.S.; Pereira, R.V.V. Molecular and Epidemiological Characterization of Bovine Intrauterine Escherichia Coli. J. Dairy Sci. 2010, 93, 5818–5830. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Rycroft, A.N.; Dogan, B.; Craven, M.; Bromfield, J.J.; Chandler, A.; Roberts, M.H.; Price, S.B.; Gilbert, R.O.; Simpson, K.W. Specific Strains of Escherichia Coli Are Pathogenic for the Endometrium of Cattle and Cause Pelvic Inflammatory Disease in Cattle and Mice. PLoS ONE 2010, 5, e9192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, M.; Peter, S.; Wagener, K.; Drillich, M.; Ehling-Schulz, M.; Einspanier, R.; Gabler, C. Bovine Endometrial Epithelial Cells Scale Their pro-Inflammatory Response in Vitro to Pathogenic Trueperella Pyogenes Isolated from the Bovine Uterus in a Strain-Specific Manner. Front. Cell. Infect. Microbiol. 2017, 7, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiano, R.B.; Bonilla, J.; Moreno, A.M.; Baruselli, P.S. Clinical Endometritis with Trueperella Pyogenes Reduces Reproductive Performance and Milk Production in Dairy Cows. Reprod. Domest. Anim. 2021, 56, 1536–1542. [Google Scholar] [CrossRef]
- Wagener, K.; Grunert, T.; Prunner, I.; Ehling-Schulz, M.; Drillich, M. Dynamics of Uterine Infections with Escherichia coli, Streptococcus uberis and Trueperella pyogenes in Post-Partum Dairy Cows and Their Association with Clinical Endometritis. Vet. J. 2014, 202, 527–532. [Google Scholar] [CrossRef]
- Ballas, P.; Gabler, C.; Wagener, K.; Drillich, M.; Ehling-Schulz, M. Streptococcus Uberis Strains Originating from Bovine Uteri Provoke Upregulation of pro-Inflammatory Factors mRNA Expression of Endometrial Epithelial Cells in Vitro. Vet. Microbiol. 2020, 245, 108710. [Google Scholar] [CrossRef]
- Gärtner, M.A.; Peter, S.; Jung, M.; Drillich, M.; Einspanier, R.; Gabler, C. Increased mRNA Expression of Selected pro-Inflammatory Factors in Inflamed Bovine Endometrium in Vivo as Well as in Endometrial Epithelial Cells Exposed to Bacillus pumilus in Vitro. Reprod. Fertil. Dev. 2016, 28, 982–994. [Google Scholar] [CrossRef]
- Fellahi, S.; Chibani, A.; Feuk-Lagerstedt, E.; Taherzadeh, M.J. Identification of Two New Keratinolytic Proteases from a Bacillus Pumilus Strain Using Protein Analysis and Gene Sequencing. AMB Express 2016, 6, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasimanickam, R.; Duffield, T.F.; Foster, R.A.; Gartley, C.J.; Leslie, K.E.; Walton, J.S.; Johnson, W.H. A Comparison of the Cytobrush and Uterine Lavage Techniques to Evaluate Endometrial Cytology in Clinically Normal Postpartum Dairy Cows. Can. Vet. J. 2005, 46, 255–259. [Google Scholar] [PubMed]
- Betts, J.G.; Hansen, P.J. Regulation of Prostaglandin Secretion from Epithelial and Stromal Cells of the Bovine Endometrium by Interleukin-1β, Interleukin-2, Granulocyte-Macrophage Colony Stimulating Factor and Tumor Necrosis Factor-α. Life Sci. 1992, 51, 1171–1176. [Google Scholar] [CrossRef]
- Viani, F.C.; Dos Santos, J.I.; Paula, C.R.; Larsony, C.E.; Gambale, W. Production of Extracellular Enzymes by Microsporum Canis and Their Role in Its Virulence. Med. Mycol. 2001, 39, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhao, H.; Wang, X.; Cui, X.; Jin, L. Keratin 86 Is up-Regulated in the Uterus during Implantation, Induced by Oestradiol. BMC Dev. Biol. 2020, 20, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballas, P.; Gabler, C.; Wagener, K.; Drillich, M.; Ehling-Schulz, M. Characterization of Bacillus pumilus Strains Isolated from Bovine Uteri. Animals 2023, 13, 1297. https://doi.org/10.3390/ani13081297
Ballas P, Gabler C, Wagener K, Drillich M, Ehling-Schulz M. Characterization of Bacillus pumilus Strains Isolated from Bovine Uteri. Animals. 2023; 13(8):1297. https://doi.org/10.3390/ani13081297
Chicago/Turabian StyleBallas, Panagiotis, Christoph Gabler, Karen Wagener, Marc Drillich, and Monika Ehling-Schulz. 2023. "Characterization of Bacillus pumilus Strains Isolated from Bovine Uteri" Animals 13, no. 8: 1297. https://doi.org/10.3390/ani13081297