

Review of the Effects of Enclosure Complexity and Design on the Behaviour and Physiology of Zoo Animals
 and
and {kind=link}
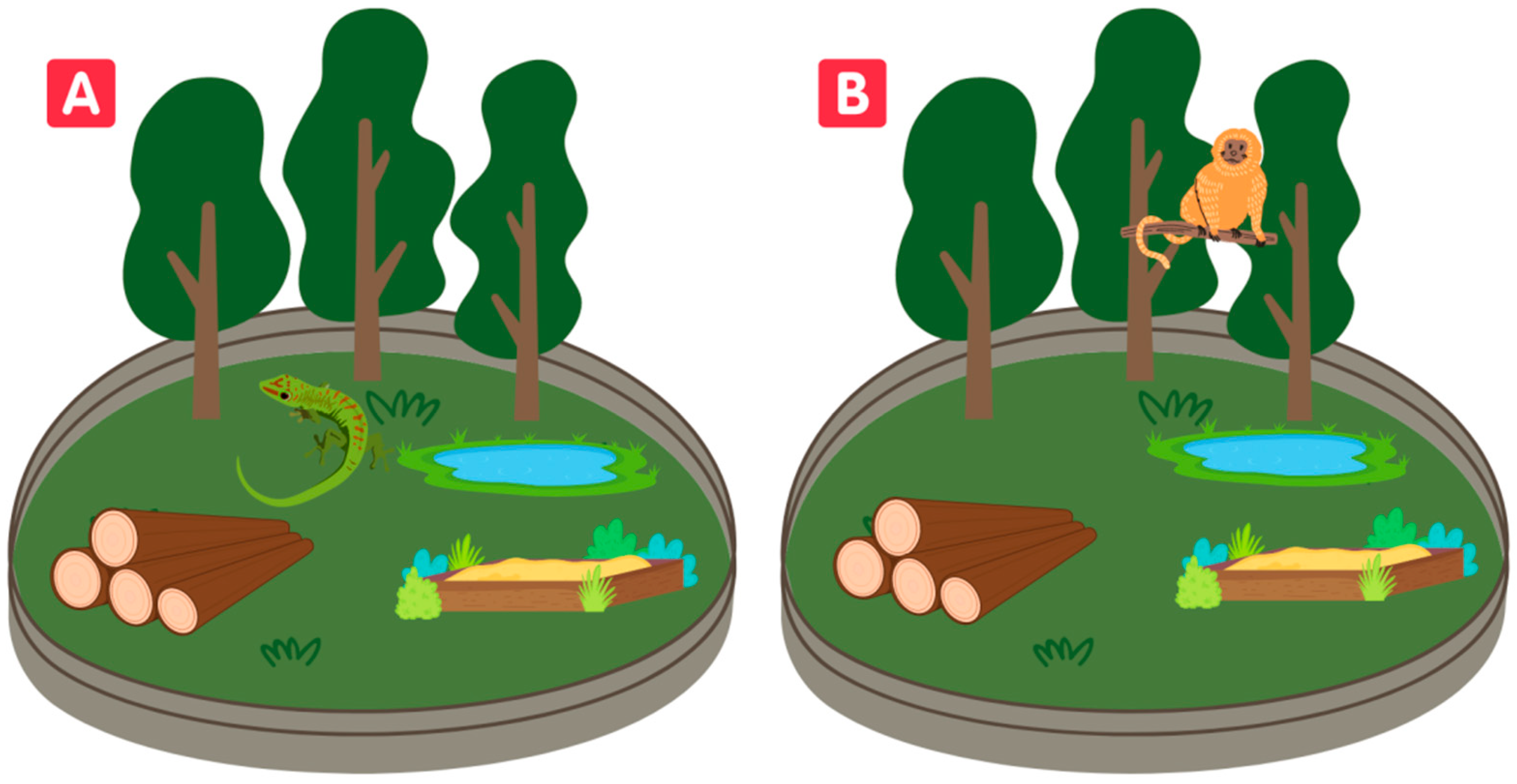{kind=link}
Abstract
:Simple Summary
Abstract
1. Habitat Complexity: Definition, Temporal Variation, and Measures
2. Why Is It Important to Live in a Complex Habitat?
3. How Does Complexity Affects Species?
4. Exhibit Renovations, Environmental Enrichment, and Concepts Aiming at Habitat Complexity
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat Complexity: Approaches and Future Directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Bracewell, S.A.; Clark, G.F.; Johnston, E.L. Habitat Complexity Effects on Diversity and Abundance Differ with Latitude: An Experimental Study over 20 Degrees. Ecology 2018, 99, 1964–1974. [Google Scholar] [CrossRef] [PubMed]
- Loke, L.H.L.; Chisholm, R.A. Measuring Habitat Complexity and Spatial Heterogeneity in Ecology. Ecol. Lett. 2022, 25, 2269–2288. [Google Scholar] [CrossRef] [PubMed]
- Strain, E.M.A.; Steinberg, P.D.; Vozzo, M.; Johnston, E.L.; Abbiati, M.; Aguilera, M.A.; Airoldi, L.; Aguirre, J.D.; Ashton, G.; Bernardi, M.; et al. A Global Analysis of Complexity–Biodiversity Relationships on Marine Artificial Structures. Glob. Ecol. Biogeogr. 2021, 30, 140–153. [Google Scholar] [CrossRef]
- Hurlbert, A.H. Species-Energy Relationships and Habitat Complexity in Bird Communities. Ecol. Lett. 2004, 7, 714–720. [Google Scholar] [CrossRef]
- Pruitt, J.N.; Cote, J.; Ferrari, M.C.O. Behavioural Trait Variants in a Habitat-Forming Species Dictate the Nature of Its Interactions with and among Heterospecifics. Funct. Ecol. 2012, 26, 29–36. [Google Scholar] [CrossRef]
- Oliveira, R.F.; Bshary, R. Expanding the Concept of Social Behavior to Interspecific Interactions. Ethology 2021, 127, 758–773. [Google Scholar] [CrossRef]
- Veasey, J.S.; Waran, N.K.; Young, R.J. On Comparing the Behaviour of Zoo Housed Animals with Wild Conspecifics as a Welfare Indicator. Anim. Welf. 1996, 5, 13–24. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Rylands, A.B.; Wilson, D.E. Handbook of the Mammals of the World; Lynx Editions: Barcelona, Spain, 2013; Volume 3. [Google Scholar]
- Healy, K.; McNally, L.; Ruxton, G.D.; Cooper, N.; Jackson, A.L. Metabolic Rate and Body Size Are Linked with Perception of Temporal Information. Anim. Behav. 2013, 86, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Bonan, G. Ecological Climatology: Concepts and Applications, 3rd ed.; Cambridge University Press: Cambridge, UK, 2015; ISBN 9781107619050. [Google Scholar]
- Pincebourde, S.; Murdock, C.C.; Vickers, M.; Sears, M.W. Fine-Scale Microclimatic Variation Can Shape the Responses of Organisms to Global Change in Both Natural and Urban Environments. Integr. Comp. Biol. 2016, 56, 45–61. [Google Scholar] [CrossRef] [Green Version]
- Milling, C.R.; Rachlow, J.L.; Olsoy, P.J.; Chappell, M.A.; Johnson, T.R.; Forbey, J.S.; Shipley, L.A.; Thornton, D.H. Habitat Structure Modifies Microclimate: An Approach for Mapping Fine-Scale Thermal Refuge. Methods Ecol. Evol. 2018, 9, 1648–1657. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Gardner, A.S.; Gaston, K.J. Diel Niche Variation in Mammals Associated with Expanded Trait Space. Nat. Commun. 2021, 12, 1753. [Google Scholar] [CrossRef]
- Dawson, M.N. Species Richness, Habitable Volume, and Species Densities in Freshwater, the Sea, and on Land. Front. Biogeogr. 2012, 4, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J. Plant Diversity in Tropical Forests: A Review of Mechanisms of Species Coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef]
- Ingram, T.; Costa-Pereira, R.; Araújo, M.S. The Dimensionality of Individual Niche Variation. Ecology 2018, 99, 536–549. [Google Scholar] [CrossRef]
- Myers, N. Biodiversity Hotspot Revisited. Bioscience 2003, 53, 916–917. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermeier2, R.A.; Mittermeier2, C.G.; Da Fonseca3, G.A.B.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Killacky, M.S. Home Is Where the Ice Is: Polar Bears Teach Us about the Arctic. Conserv. Physiol. 2021, 9, coab013. [Google Scholar] [CrossRef]
- Garneau, D.E.; Boudreau, T.; Keech, M.; Post, E. Habitat Use by Black Bears in Relation to Conspecifics and Competitors. Mamm. Biol. 2008, 73, 48–57. [Google Scholar] [CrossRef]
- van Bommel, J.K.; Sun, C.; Ford, A.T.; Todd, M.; Burton, A.C. Coexistence or Conflict: Black Bear Habitat Use along an Urban-Wildland Gradient. PLoS ONE 2022, 17, e0276448. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Clutton-Brock, T.; Shultz, S.; Lukas, D. Social Complexity: Patterns, Processes, and Evolution. Behav. Ecol. Sociobiol. 2019, 73, 5. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. The Social Brain Hypothesis. Evol. Anthropol. 1998, 6, 178–190. [Google Scholar] [CrossRef]
- Dunbar, R.I.M. The Social Brain Hypothesis and Its Implications for Social Evolution. Ann. Hum. Biol. 2009, 36, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; McShea, D.W. Individual versus Social Complexity, with Particular Reference to Ant Colonies. Biol. Rev. Camb. Philos. Soc. 2001, 76, 211–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taborsky, M.; Wong, M. Sociality in Fishes. In Comparative Social Evolution; Cambridge University Press: Cambridge, UK, 2017; pp. 354–389. [Google Scholar]
- Baker, K.C.; Bloomsmith, M.; Neu, K.; Griffis, C.; Maloney, M.; Oettinger, B.; Schoof, V.A.M.; Martinez, M. Positive Reinforcement Training Moderates Only High Levels of Abnormal Behavior in Singly Housed Rhesus Macaques. J. Appl. Anim. Welf. Sci. 2009, 12, 236–252. [Google Scholar] [CrossRef] [Green Version]
- Osorio, D.; Vorobyev, M. A Review of the Evolution of Animal Colour Vision and Visual Communication Signals. Vision Res. 2008, 48, 2042–2051. [Google Scholar] [CrossRef] [Green Version]
- Kelber, A. Birds Perceive Colours in Categories. Nature 2018, 560, 211–212. [Google Scholar] [CrossRef] [Green Version]
- Page, R.A.; ter Hofstede, H.M. Sensory and Cognitive Ecology of Bats. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 541–562. [Google Scholar] [CrossRef]
- Fieschi-Méric, L.; Ellis, C.; Servini, F.; Tapley, B.; Michaels, C.J. An Improvement in Enclosure Design Can Positively Impact Welfare, Reduce Aggressiveness and Stabilise Hierarchy in Captive Galapagos Giant Tortoises. J. Zool. Bot. Gard. 2022, 3, 499–512. [Google Scholar] [CrossRef]
- Young, R.J. Environmental Enrichment for Captive Animals, 1st ed.; Blackwell Science Ltda.: Oxford, UK, 2003. [Google Scholar]
- Azevedo, C.S.; Cipreste, C.F.; Pizzutto, C.S. Fundamentos Do Enriquecimento Ambiental, 1st ed.; Payá: São Paulo, Brazil, 2022. [Google Scholar]
- O’Connor, A.M.; Burton, T.J.; Leamey, C.A.; Sawatari, A. The Use of the Puzzle Box as a Means of Assessing the Efficacy of Environmental Enrichment. J. Vis. Exp. 2014, 94, e52225. [Google Scholar] [CrossRef] [Green Version]
- De Groot, B.; Cheyne, S.M. Does Mirror Enrichment Improve Primate Well-Being? Anim. Welf. 2016, 25, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Coe, J. Embedding Environmental Enrichment into Zoo Animal Facility Design. In Proceedings of the Zoo Design Conference, Wroclaw, Poland, 7 April 2017; pp. 1–21. [Google Scholar]
- Kistler, C.; Hegglin, D.; Würbel, H.; König, B. Structural Enrichment and Enclosure Use in an Opportunistic Carnivore: The Red Fox (Vulpes Vulpes). Anim. Welf. 2010, 19, 391–400. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Nuñez-Velazquez, S.; Diaz-Gil, C.; Follana-Berná, G.; Saraiva, J.L. Suspended Structures Reduce Variability of Group Risk-Taking Responses of Dicentrarchus Labrax Juvenile Reared in Tanks. Fishes 2022, 7, 126. [Google Scholar] [CrossRef]
- Dancer, A.M.M.; Díez-león, M.; Bizley, J.K.; Burn, C.C. Housing and Environmental Enrichment of the Domestic Ferret: A Multi-Sector Survey. Animals 2022, 12, 1065. [Google Scholar] [CrossRef]
- Orban, D.A.; Soltis, J.; Perkins, L.; Mellen, J.D. Sound at the Zoo: Using Animal Monitoring, Sound Measurement, and Noise Reduction in Zoo Animal Management. Zoo Biol. 2017, 36, 231–236. [Google Scholar] [CrossRef]
- Chiew, S.J.; Butler, K.L.; Sherwen, S.L.; Coleman, G.J.; Melfi, V.; Burns, A.; Hemsworth, P.H. Effect of Covering a Visitor Viewing Area Window on the Behaviour of Zoo-Housed Little Penguins (Eudyptula Minor). Animals 2020, 10, 1224. [Google Scholar] [CrossRef]
- Davey, G. Visitors’ Effects on the Welfare of Animals in the Zoo: A Review. J. Appl. Anim. Welf. Sci. 2007, 10, 169–183. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Hemsworth, P.H. The Visitor Effect on Zoo Animals: Implications and Opportunities for Zoo Animal Welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Wark, J.D.; Wierzal, N.K.; Cronin, K.A. Mapping Shade Availability and Use in Zoo Environments: A Tool for Evaluating Thermal Comfort. Animals 2020, 10, 1189. [Google Scholar] [CrossRef]
- Jones, N.; Sherwen, S.L.; Robbins, R.; McLelland, D.J.; Whittaker, A.L. Welfare Assessment Tools in Zoos: From Theory to Practice. Vet. Sci. 2022, 9, 170. [Google Scholar] [CrossRef]
- Buchanan-Smith, H.M. Environmental Enrichment for Primates in Laboratories. Adv. Sci. Res. 2011, 5, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Näslund, J.; Johnsson, J.I. Environmental Enrichment for Fish in Captive Environments: Effects of Physical Structures and Substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Woods, J.M.; Eyer, A.; Miller, L.J. Bird Welfare in Zoos and Aquariums: General Insights across Industries. J. Zool. Bot. Gard. 2022, 3, 198–222. [Google Scholar] [CrossRef]
- Shumway, C.A. Habitat Complexity, Brain, and Behavior. Brain Behav. Evol. 2008, 72, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, K.; Rennstam Rubbmark, O.; Birkhofer, K.; Malsher, G.; Sint, D.; Jonsson, M.; Traugott, M. Habitat Heterogeneity Induces Rapid Changes in the Feeding Behaviour of Generalist Arthropod Predators. Funct. Ecol. 2018, 32, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, D.S. Catecholamines and Stress. Endocr. Regul. 2003, 37, 69–80. [Google Scholar]
- Bayazit, V. Evaluation of Cortisol and Stress in Captive Animals. Aust. J. Basic Appl. Sci. 2009, 3, 1022–1031. [Google Scholar]
- Fahey, G.C.; Barry, K.A.; Swanson, K.S. Age-Related Changes in Nutrient Utilization by Companion Animals. Annu. Rev. Nutr. 2008, 28, 425–445. [Google Scholar] [CrossRef]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 Approach to Promoting Optimal Welfare for Captive Wild Animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Young, R.J. Animal Personality and Conservation: Basics for Inspiring New Research. Animals 2021, 11, 1019. [Google Scholar] [CrossRef]
- Schirmer, A.; Herde, A.; Eccard, J.A.; Dammhahn, M. Individuals in Space: Personality-Dependent Space Use, Movement and Microhabitat Use Facilitate Individual Spatial Niche Specialization. Oecologia 2019, 189, 647–660. [Google Scholar] [CrossRef]
- Webster, M.M.; Atton, N.; Hoppitt, W.J.E.; Laland, K.N. Environmental Complexity Influences Association Network Structure and Network-Based Diffusion of Foraging Information in Fish Shoals. Am. Nat. 2013, 181, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Briffa, M.; Sneddon, L.U.; Wilson, A.J. Animal Personality as a Cause and Consequence of Contest Behaviour. Biol. Lett. 2015, 11, 20141007. [Google Scholar] [CrossRef] [Green Version]
- Henry, M.; Thomas, D.W.; Al Vaudry, R.É.; Carrier, M. Foraging Distances and Home Range of Pregnant and Lactating Little Brown Bats (Myotis Lucifugus). J. Mammal. 2002, 83, 767–774. [Google Scholar]
- Sohel Khan, A.; Lea, S.E.G.; Chand, P.; Rai, U.; Baskaran, N. Predictors of Psychological Stress and Behavioural Diversity among Captive Red Panda in Indian Zoos and Their Implications for Global Captive Management. Sci. Rep. 2022, 12, 14034. [Google Scholar] [CrossRef]
- He, P.; Maldonado-Chaparro, A.A.; Farine, D.R. The Role of Habitat Configuration in Shaping Social Structure: A Gap in Studies of Animal Social Complexity. Behav. Ecol. Sociobiol. 2019, 73, 9. [Google Scholar] [CrossRef] [Green Version]
- Kappeler, P.M. A Framework for Studying Social Complexity. Behav. Ecol. Sociobiol. 2019, 73, 13. [Google Scholar] [CrossRef]
- Palagi, E.; Bergman, T.J. Bridging Captive and Wild Studies: Behavioral Plasticity and Social Complexity in Theropithecus Gelada. Animals 2021, 11, 3003. [Google Scholar] [CrossRef]
- Galardi, E.G.; Fabbroni, M.; Rausa, F.A.; Preziosi, R.; Brereton, J.E.; Pastorino, G.Q. An Investigation into the Behavior, Sociality and Enclosure Use of Group-Housed Lions and Tigers. J. Vet. Med. Anim. Sci. 2021, 4, 1068. [Google Scholar]
- Truelove, M.A.; Martin, A.L.; Perlman, J.E.; Wood, J.S.; Bloomsmith, M.A. Pair Housing of Macaques: A Review of Partner Selection, Introduction Techniques, Monitoring for Compatibility, and Methods for Long-Term Maintenance of Pairs. Am. J. Primatol. 2017, 79, e22485. [Google Scholar] [CrossRef]
- Aureli, F.; Schino, G. Social Complexity from within: How Individuals Experience the Structure and Organization of Their Groups. Behav. Ecol. Sociobiol. 2019, 73, 6. [Google Scholar] [CrossRef]
- Hopper, L.M. Leveraging Social Learning to Enhance Captive Animal Care and Welfare. J. Zool. Bot. Gard. 2021, 2, 21–40. [Google Scholar] [CrossRef]
- Kleinhappel, T.K.; John, E.A.; Pike, T.W.; Wilkinson, A.; Burman, O.H.P. Animal Welfare: A Social Networks Perspective. Sci. Prog. 2016, 99, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Serres, A.; Hao, Y.; Wang, D. Body Contacts and Social Interactions in Captive Odontocetes Are Influenced by the Context: An Implication for Welfare Assessment. Animals 2020, 10, 924. [Google Scholar] [CrossRef]
- Hobson, E.A.; Mønster, D.; Dedeo, S.; Strassmann, J.E. Aggression Heuristics Underlie Animal Dominance Hierarchies and Provide Evidence of Group-Level Social Information. Proc. Natl. Acad. Sci. USA 2021, 118, e2022912118. [Google Scholar] [CrossRef]
- Pinheiro, T.; Ferrari, S.F.; Lopes, M.A. Polyspecific Associations between Squirrel Monkeys (Saimiri Sciureus) and Other Primates in Eastern Amazonia. Am. J. Primatol. 2011, 73, 1145–1151. [Google Scholar] [CrossRef]
- Sanz, C.M.; Strait, D.; Eyana Ayina, C.; Massamba, J.M.; Ebombi, T.F.; Ndassoba Kialiema, S.; Ngoteni, D.; Mbebouti, G.; Koni Boue, D.R.; Brogan, S.; et al. Interspecific Interactions between Sympatric Apes. iScience 2022, 25, 105059. [Google Scholar] [CrossRef]
- Kaplan, G. Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork. Animals 2022, 12, 31. [Google Scholar] [CrossRef]
- Svábik, K. Mixed-Species Exhibits with Carnivores (Carnivora)—Summary. Available online: https://zoolex.info/media/uploads/2021/05/02/2020_svabik_mixed_dogs.pdf (accessed on 27 January 2023).
- Foulds-Davis, Y.L. To Mix or Not to Mix? Evaluating Breeding Performance in Mixed Species Bird Enclosures within European Zoos. Master’s Thesis, Liverpool John Moores University, Liverpool, UK, 2015. [Google Scholar]
- Kaandorp, J. Veterinary Challenges of Mixed Species Exhibits. In Fowler’s Zoo and Wild Animal Medicine; Elsevier: Amsterdam, The Netherlands, 2012; pp. 24–31. [Google Scholar]
- Serra, M.; Patricia, C.; Wolkers, B.; Urbinati, E.C. Physiological Indicators of Animal Welfare. Rev. Bras. Zoociências 2018, 19, 70–96. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, A.B.; Vigors, B.; Sandøe, P. What Is so Positive about Positive Animal Welfare?—A Critical Review of the Literature. Animals 2019, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Ritzler, C.P.; Lukas, K.E.; Bernstein-Kurtycz, L.M.; Koester, D.C. The Effects of Choice-Based Design and Management on the Behavior and Space Use of Zoo-Housed Amur Tigers (Panthera Tigris Altaica). J. Appl. Anim. Welf. Sci. 2021, 6, 1–14. [Google Scholar] [CrossRef]
- Nicol, C.J.; Caplen, G.; Edgar, J.; Browne, W.J. Associations between Welfare Indicators and Environmental Choice in Laying Hens. Anim. Behav. 2009, 78, 413–424. [Google Scholar] [CrossRef]
- Hutchinson, J.M.C. Is More Choice Always Desirable? Evidence and Arguments from Leks, Food Selection, and Environmental Enrichment. Biol. Rev. Camb. Philos. Soc. 2005, 80, 73–92. [Google Scholar] [CrossRef]
- Allard, S.; Fuller, G.; Torgerson-White, L.; Starking, M.D.; Yoder-Nowak, T. Personality in Zoo-Hatched Blanding’s Turtles Affects Behavior and Survival after Reintroduction into the Wild. Front. Psychol. 2019, 10, 2324. [Google Scholar] [CrossRef]
- Kitchen, A.M.; Martin, A.A. The Effects of Cage Size and Complexity on the Behaviour of Captive Common Marmosets, Callithrix Jacchus Jacchus. Lab. Anim. 1996, 30, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.A. Minimum Standards for Wildlife Rehabilitation, 4th ed.; National Wildlife Rehabilitators Association: Minnesota, MN, USA, 2012; ISBN 9781931439282. [Google Scholar]
- Marino, L.; Rose, N.A.; Visser, I.N.; Rally, H.; Ferdowsian, H.; Slootsky, V. The Harmful Effects of Captivity and Chronic Stress on the Well-Being of Orcas (Orcinus Orca). J. Vet. Behav. 2020, 35, 69–82. [Google Scholar] [CrossRef]
- Scrine, J.; Jochum, M.; Ólafsson, J.S.; O’Gorman, E.J. Interactive Effects of Temperature and Habitat Complexity on Freshwater Communities. Ecol. Evol. 2017, 7, 9333–9346. [Google Scholar] [CrossRef]
- Nay, T.J.; Johansen, J.L.; Rummer, J.L.; Steffensen, J.F.; Pratchett, M.S.; Hoey, A.S. Habitat Complexity Influences Selection of Thermal Environment in a Common Coral Reef Fish. Conserv. Physiol. 2020, 8, coaa070. [Google Scholar] [CrossRef]
- Keskitalo, J.; Bergquist, G.; Gardeström, P.; Jansson, S. A Cellular Timetable of Autumn Senescence. Plant Physiol. 2005, 139, 1635–1648. [Google Scholar] [CrossRef] [Green Version]
- Grogan, J.; Schulze, M. The Impact of Annual and Seasonal Rainfall Patterns on Growth and Phenology of Emergent Tree Species in Southeastern Amazonia, Brazil. Biotropica 2012, 44, 331–340. [Google Scholar] [CrossRef]
- Cissé, S.; Eymard, L.; Ottlé, C.; Ndione, J.A.; Gaye, A.T.; Pinsard, F. Rainfall Intra-Seasonal Variability and Vegetation Growth in the Ferlo Basin (Senegal). Remote Sens. 2016, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Bånkestad, D.; Hoch, G. Reaching Natural Growth: The Significance of Light and Temperature Fluctuations in Plant Performance in Indoor Growth Facilities. Plants 2020, 9, 1312. [Google Scholar] [CrossRef] [PubMed]
- Valdez, D.J.; Vera Cortez, M.; Della Costa, N.S.; Lèche, A.; Hansen, C.; Navarro, J.L.; Martella, M.B. Seasonal Changes in Plasma Levels of Sex Hormones in the Greater Rhea (Rhea Americana), a South American Ratite with a Complex Mating System. PLoS ONE 2014, 9, e97334. [Google Scholar] [CrossRef] [PubMed]
- Young, R.J. The Importance of Food Presentation for Animal Welfare and Conservation. Proc. Nutr. Soc. 1997, 56, 1095–1104. [Google Scholar] [CrossRef] [Green Version]
- Cenni, F.; Parisi, G.; Gherardi, F. Effects of Habitat Complexity on the Aggressive Behaviour of the American Lobster (Homarus Americanus) in Captivity. Appl. Anim. Behav. Sci. 2010, 122, 63–70. [Google Scholar] [CrossRef]
- Xiaofeng, L.; Changjie, Z.; Cenyi, H.; Xiuxiang, M. Seasonal Variation in the Behavior of Captive Alpine Musk Deer, Moschus Sifanicus, in Xinglongshan Musk Deer Farm, of China. Zoologia 2010, 27, 848–852. [Google Scholar] [CrossRef]
- Versteeg, E.J.; Fernandes, T.; Guzzo, M.M.; Laberge, F.; Middel, T.; Ridgway, M.; McMeans, B.C. Seasonal Variation of Behavior and Brain Size in a Freshwater Fish. Ecol. Evol. 2021, 11, 14950–14959. [Google Scholar] [CrossRef]
- Eguizábal, G.V.; Superina, M.; Palme, R.; Asencio, C.J.; Villarreal, D.P.; Borrelli, L.; Busso, J.M. Non-Invasive Assessment of the Seasonal Stress Response to Veterinary Procedures and Transportation of Zoo-Housed Lesser Anteater (Tamandua Tetradactyla). Animals 2022, 12, 75. [Google Scholar] [CrossRef]
- Coloma-García, W.; Mehaba, N.; Such, X.; Caja, G.; Salama, A.A.K. Effects of Cold Exposure on Some Physiological, Productive, and Metabolic Variables in Lactating Dairy Goats. Animals 2020, 10, 2383. [Google Scholar] [CrossRef]
- Tochihara, Y.; Wakabayashi, H.; Lee, J.Y.; Wijayanto, T.; Hashiguchi, N.; Saat, M. How Humans Adapt to Hot Climates Learned from the Recent Research on Tropical Indigenes. J. Physiol. Anthropol. 2022, 41, 27. [Google Scholar] [CrossRef]
- Bauert, M.R.; Furrer, S.C.; Zingg, R.; Steinmetz, H.W. Three Years of Experience Running the Masoala Rainforest Ecosystem at Zurich Zoo, Switzerland. Int. Zoo Yearb. 2007, 41, 203–216. [Google Scholar] [CrossRef]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s New from the Zoo? An Analysis of Ten Years of Zoo-Themed Research Output. Palgrave Commun. 2019, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Brereton, J.; Fernandez, E. Which Index Should I Use? A Comparison of Indices for Enclosure Use Studies. Anim. Behav. Cogn. 2022, 9, 119–132. [Google Scholar] [CrossRef]
- Plowman, A.B. A Note on a Modification of the Spread of Participation Index Allowing for Unequal Zones. Appl. Anim. Behav. Sci. 2003, 83, 331–336. [Google Scholar] [CrossRef]
- Prystupczuk, L.; Leynaud, G.C.; Labaque, C. Evaluating Behavior and Enclosure Use of Zoo-Housed Broad-Snouted Caiman (Caiman Latirostris): A Case Study in the Zoo of Córdoba (Argentina). Herpetol. Conserv. Biol. 2019, 14, 601–614. [Google Scholar]
- Rose, P.; Robert, R. Evaluating the Activity Patterns and Enclosure Usage of a Little-Studied Zoo Species, the Sitatunga (Tragelaphus Spekii). J. Zoo Aquar. Res. 2013, 1, 14–19. [Google Scholar] [CrossRef]
- Clark, F.E.; Melfi, V.A. Environmental Enrichment for a Mixed-Species Nocturnal Mammal Exhibit. Zoo Biol. 2012, 31, 397–413. [Google Scholar] [CrossRef]
- Checon, C.T.; Rosenfield, D.A.; Jorge-Neto, P.N.; Pizzutto, C.S. Influence of Environmental Enrichment on the Behavioral Variables of Caged Hyacinth Macaws (Anodorhynchus Hyacinthinus). Ornithol. Res. 2020, 28, 125–132. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behaviour as a Tool in the Assessment of Animal Welfare. Zoology 2003, 106, 383–387. [Google Scholar] [CrossRef]
- Broom, D.M. Welfare of Animals: Behavior as a Basis for Decisions. In Encyclopedia of Animal Behavior; Breed, M.D., Moore, J., Eds.; Academic Press: Oxford, UK, 2010; Volume 3, pp. 580–584. [Google Scholar]
- Wemelsfelder, F.; Haskell, M.; Mendl, M.T.; Calvert, S.; Lawrence, A.B. Diversity of Behaviour during Novel Object Tests Is Reduced in Pigs Housed in Substrate-Impoverished Conditions. Anim. Behav. 2000, 60, 385–394. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.R.; Sloman, K.A. The Effects of Environmental Pollutants on Complex Fish Behaviour: Integrating Behavioural and Physiological Indicators of Toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Cochet, H.; Byrne, R.W. Complexity in Animal Behaviour: Towards Common Ground. Acta Ethol. 2015, 18, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Relative Importance of Spatial and Temporal Scales in a Patchy Environment. Theor. Popul Biol 1992, 41, 300–314. [Google Scholar] [CrossRef]
- McPhee, M.E. Effects of Captivity on Response to a Novel Environment in the Oldfield Mouse (Peromyscus Polionotus Subgriseus). Int. J. Comp. Psychol. 2003, 16, 85–94. [Google Scholar] [CrossRef]
- Bacon, H. Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors. Animals 2018, 8, 103. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.J.; Latham, N.R. Can’t Stop, Won’t Stop: Is Stereotypy a Reliable Animal Welfare Indicator? Animal Welfare 2004, 13, S57–S69. [Google Scholar] [CrossRef]
- Yeates, J. Naturalness and Animal Welfare. Animals 2018, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Browning, H. The Natural Behavior Debate: Two Conceptions of Animal Welfare. J. Appl. Anim. Welf. Sci. 2020, 23, 325–337. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific Approaches to Enrichment and Stereotypies in Zoo Animals: What’s Been Done and Where Should We Go Next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Ball, N.J.; Mercado, E.; Orduña, I. Enriched Environments as a Potential Treatment for Developmental Disorders: A Critical Assessment. Front. Psychol. 2019, 10, 466. [Google Scholar] [CrossRef] [Green Version]
- Chudeau, K.R.; Johnson, S.P.; Caine, N.G. Enrichment Reduces Stereotypical Behaviors and Improves Foraging Development in Rehabilitating Eastern Pacific Harbor Seals (Phoca Vitulina Richardii). Appl. Anim. Behav. Sci. 2019, 219, 104830. [Google Scholar] [CrossRef]
- Lima, M.F.F.; de Azevedo, C.S.; Young, R.J.; Viau, P. Impacts of Food-Based Enrichment on Behaviour and Physiology of Male Greater Rheas (Rhea Americana, Rheidae, Aves). Pap. Avulsos Zool. 2019, 59. [Google Scholar] [CrossRef]
- Mkwanazi, M.V.; Ncobela, C.N.; Kanengoni, A.T.; Chimonyo, M. Effects of Environmental Enrichment on Behaviour, Physiology and Performance of Pigs—A Review. Asian-Australas. J. Anim. Sci. 2019, 32, 1–13. [Google Scholar] [CrossRef]
- Mellor, D.J. Moving beyond the “Five Freedoms” by Updating the “Five Provisions” and Introducing Aligned “Animal Welfare Aims”. Animals 2016, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, M.S. Animal Welfare and Efficient Farming: Is Conflict Inevitable? Anim. Prod. Sci. 2017, 57, 201–208. [Google Scholar] [CrossRef]
- Greggor, A.L.; Vicino, G.A.; Swaisgood, R.R.; Fidgett, A.; Brenner, D.; Kinney, M.E.; Farabaugh, S.; Masuda, B.; Lamberski, N. Animal Welfare in Conservation Breeding: Applications and Challenges. Front. Vet. Sci. 2018, 5, 323. [Google Scholar] [CrossRef]
- Maryam Jamilah, I.; Darsono, A.; Fathurrahman, I.; Sonia, M. Animal Welfare As Stress Management to Improve Beef Cattle Reproduction. KnE Life Sci. 2019, 4, 200. [Google Scholar] [CrossRef] [Green Version]
- Moura, D.J.; Naas, I.A.; Pereira, D.F.; Silva, R.B.T.R.; Camargo, G.A. Animal Welfare Concepts and Strategy for Poultry Production: A Review. Braz. J. Poltry Sci. 2006, 8, 137–148. [Google Scholar] [CrossRef]
- Eizaguirre, C.; Baltazar-Soares, M. Evolutionary Conservation-Evaluating the Adaptive Potential of Species. Evol. Appl. 2014, 7, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Rey, O.; Eizaguirre, C.; Angers, B.; Baltazar-Soares, M.; Sagonas, K.; Prunier, J.G.; Blanchet, S. Linking Epigenetics and Biological Conservation: Towards a Conservation Epigenetics Perspective. Funct. Ecol. 2020, 34, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Lamka, G.F.; Harder, A.M.; Sundaram, M.; Schwartz, T.S.; Christie, M.R.; DeWoody, J.A.; Willoughby, J.R. Epigenetics in Ecology, Evolution, and Conservation. Front. Ecol. Evol. 2022, 10, 871791. [Google Scholar] [CrossRef]
- Johnstone, S.E.; Baylin, S.B. Stress and the Epigenetic Landscape: A Link to the Pathobiology of Human Diseases? Nat. Rev. Genet. 2010, 11, 806–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, B.; Rally, H.; Doyle, C.; O’Brien, L.; Tennison, M.; Marino, L. Putative Neural Consequences of Captivity for Elephants and Cetaceans. Rev. Neurosci. 2022, 33, 439–465. [Google Scholar] [CrossRef] [PubMed]
- Holt, W.V.; Comizzoli, P. Conservation Biology and Reproduction in a Time of Developmental Plasticity. Biomolecules 2022, 12, 1297. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, A.; Guerrero-Altamirano, T.P.; Yasar, S.; Czirják, G.; Wachter, B.; Fickel, J. First Steps towards the Development of Epigenetic Biomarkers in Female Cheetahs (Acinonyx Jubatus). Life 2022, 12, 920. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.L.; Edes, A.N.; Brown, J.L. Stress, Well-Being and Reproductive Success. Adv. Exp. Med. Biol. 2019, 1200, 91–162. [Google Scholar] [CrossRef]
- Pizzutto, C.S.; Colbachini, H.; Jorge-Neto, P.N. One Conservation: The Integrated View of Biodiversity Conservation. Anim. Reprod. 2021, 18, e20210024. [Google Scholar] [CrossRef]
- Conde, D.A.; Colchero, F.; Gusset, M.; Pearce-Kelly, P.; Byers, O.; Flesness, N.; Browne, R.K.; Jones, O.R. Zoos through the Lens of the IUCN Red List: A Global Metapopulation Approach to Support Conservation Breeding Programs. PLoS ONE 2013, 8, e80311. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D.J.; Reid, C.S.W. Concepts of Animal Well-Being and Predicting the Impact of procedures on Experimental Animals. In Improving the Well-Being of Animals in the Research Environment; WellBeing International: Potomac, MD, USA, 1994. [Google Scholar]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- von der Emde, G.W.E. The Ecology of Animal Senses: Matched Filters for Economical Sensing, 1st ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Schmidt-Rhaesa, A. The Evolution of Organ Systems, 1st ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Veilleux, C.C.; Kirk, E.C. Visual Acuity in Mammals: Effects of Eye Size and Ecology. Brain Behav. Evol. 2014, 83, 43–53. [Google Scholar] [CrossRef]
- Boström, J.E.; Dimitrova, M.; Canton, C.; Håstad, O.; Qvarnström, A.; Ödeen, A. Ultra-Rapid Vision in Birds. PLoS ONE 2016, 11, e0151099. [Google Scholar] [CrossRef] [Green Version]
- Debjani, R. Development of Hearing in Vertebrates with Special Reference to Anuran Acoustic Communication. J. Biosci. 1994, 19, 629–644. [Google Scholar] [CrossRef]
- Dooling, R.J.; Prior, N.H. Do We Hear What Birds Hear in Birdsong? Anim. Behav. 2017, 124, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Heffner, H.E.; Heffner, R.S. The Evolution of Mammalian Hearing. AIP Conf. Proc. 2018, 1965, 130001. [Google Scholar]
- Willemart, R.H.; Gnaspini, P. Comparative Density of Hair Sensilla on the Legs of Cavernicolous and Epigean Harvestmen (Arachnida: Opiliones). Zool. Anz. 2004, 242, 353–365. [Google Scholar] [CrossRef] [Green Version]
- Kasumyan, A.O. Tactile Reception and Behavior of Fish. J. Ichthyol. 2011, 51, 1035–1103. [Google Scholar] [CrossRef]
- Kelley, J.L.; Chapuis, L.; Davies, W.I.L.; Collin, S.P. Sensory System Responses to Human-Induced Environmental Change. Front. Ecol. Evol. 2018, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Hale, R.; Piggott, J.J.; Swearer, S.E. Describing and Understanding Behavioral Responses to Multiple Stressors and Multiple Stimuli. Ecol. Evol. 2017, 7, 38–47. [Google Scholar] [CrossRef]
- Stephan, H.; Andy, O.J. Quantitative Comparisons of Brain Structures from Insectivores to Primates. Am. Zool. 1964, 4, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Northcutt, R.G. Understanding Vertebrate Brain Evolution. Integr. Comp. Biol. 2002, 42, 743–756. [Google Scholar] [CrossRef]
- Corfield, J.R.; Price, K.; Iwaniuk, A.N.; Gutiérrez-Ibáñez, C.; Birkhead, T.; Wylie, D.R. Diversity in Olfactory Bulb Size in Birds Reflects Allometry, Ecology, and Phylogeny. Front. Neuroanat. 2015, 9, 102. [Google Scholar] [CrossRef] [PubMed]
- Ubeda-Bañon, I.; Pro-Sistiaga, P.; Mohedano-Moriano, A.; Saiz-Sanchez, D.; de la Rosa-Prieto, C.; Gutierrez-Castellanos, N.; Lanuza, E.; Martinez-Garcia, F.; Martinez-Marcos, A. Cladistic Analysis of Olfactory and Vomeronasal Systems. Front. Neuroanat. 2011, 5, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, S.J.; Schneider, D.C.; Schaefer, J.A.; Mahoney, S.P. Habitat Selection at Multiple Scales. Ecoscience 2009, 16, 238–247. [Google Scholar] [CrossRef]
- Buskirk, S.W.; Millspaugh, J.J. Metrics for Studies of Resource Selection. J. Wildl. Manag. 2006, 70, 358–366. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Pfaus, J.G.; Orihuela, A.; Domínguez-Oliva, A.; José-Pérez, N.; Hernández, L.A.; Mota-Rojas, D. The Neurobiology of Behavior and Its Applicability for Animal Welfare: A Review. Animals 2022, 12, 928. [Google Scholar] [CrossRef]
- Singh, S.V.; Kumar, S. Circadian Rhythm and Their Significance in Relation to Physiological Functions of Animals: A Review. J. Entomol. Zool. Stud. 2018, 6, 1861–1866. [Google Scholar]
- Helm, B.; Ben-Shlomo, R.; Sheriff, M.J.; Hut, R.A.; Foster, R.; Barnes, B.M.; Dominoni, D. Annual Rhythms That Underlie Phenology: Biological Time-Keeping Meets Environmental Change. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, G.H. Ultraviolet Vision in Vertebrates. Am. Zool. 1992, 32, 544–554. [Google Scholar] [CrossRef] [Green Version]
- McGann, J.P. Poor Human Olfaction Is a 19th-Century Myth. Science 2017, 356, eaam7263. [Google Scholar] [CrossRef] [Green Version]
- Heffner, H. Hearing Ranges of Laboratory Animals. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 20–22. [Google Scholar]
- Poole, T.B. Meeting a Mammal’s Psychological Needs: Basic Principles. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Books: Washington, DC, USA, 1998; pp. 83–94. [Google Scholar]
- Hancocks, D. The History and Principles of Zoo Exhibition. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management; Kleiman, D.G., Thompson, K.V., Baer, C.K., Eds.; The University of Chicago Press: Chicago, IL, USA, 2010; pp. 121–136. [Google Scholar]
- Hosey, G.; Melfi, V.; Pankhurst, S. Zoo Animals: Behaviour, Management, and Welfare, 2nd ed.; Hosey, G., Melfi, V., Pankhurst, S., Eds.; Oxford University Press: Oxford, UK, 2013. [Google Scholar]
- Rajchard, J. Ultraviolet (UV) Light Perception by Birds: A Review. Vet. Med. 2009, 54, 351–359. [Google Scholar] [CrossRef] [Green Version]
- House, G.M.; Sobotik, E.B.; Nelson, J.R.; Archer, G.S. Effects of Ultraviolet Light Supplementation on Pekin Duck Production, Behavior, and Welfare. Animals 2020, 10, 833. [Google Scholar] [CrossRef]
- Ross, M.R.; Gillespie, K.L.; Hopper, L.M.; Bloomsmith, M.A.; Maple, T.L. Differential Preference for Ultraviolet Light among Captive Birds from Three Ecological Habitats. Appl. Anim. Behav. Sci. 2013, 147, 278–285. [Google Scholar] [CrossRef]
- Stevens, E.F. Flamingo Breeding: The Role of Group Display. Zoo Biol. 1991, 10, 53–63. [Google Scholar] [CrossRef]
- Rose, P.E.; Brereton, J.E.; Gardner, L. Developing Flamingo Husbandry Practices through Workshop Communication. J. Zoo Aquar. Res. 2016, 4, 115–121. [Google Scholar] [CrossRef]
- Lawrence, K.; Sherwen, S.L.; Larsen, H. Natural Habitat Design for Zoo-housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit. Animals 2021, 11, 2979. [Google Scholar] [CrossRef]
- Jameson, K.A.; Winkler, A.D.; Goldfarb, K. Art, Interpersonal Comparisons of Color Experience, and Potential Tetrachromacy. Hum. Vis. Electron. Imaging 2016, 145, 354–365. [Google Scholar] [CrossRef] [Green Version]
- BBC. The Women with Superhuman Vision. 2023. Available online: https://www.bbc.com/future/article/20140905-the-women-with-super-human-vision (accessed on 12 February 2023).
- Fàbregas, M.C.; Guillén-Salazar, F.; Garcés-Narro, C. Do Naturalistic Enclosures Provide Suitable Environments for Zoo Animals? Zoo Biol. 2012, 31, 362–373. [Google Scholar] [CrossRef]
- Finch, K.; Waterman, J.O.; Cowl, V.B.; Marshall, A.; Underwood, L.; Williams, L.J.; Davis, N.; Holmes, L. Island Life: Use of Activity Budgets and Visibility to Evaluate a Multi-Species Within-Zoo Exhibit Move. Animals 2022, 12, 2123. [Google Scholar] [CrossRef]
- Learmonth, M.J. Dilemmas for Natural Living Concepts of Zoo Animal Welfare. Animals 2019, 9, 318. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.E.; Rowe, C.; Mackay, F.; Matthews, C.; Matthews, C.G.G. Aquarium Tank Design Is Integral to the Elimination of Mantle Abrasion in the Captive Curled Octopus (Eledone Cirrhosa): A Case Study at Macduff Marine Aquarium. J. Appl. Anim. Welf. Sci. 2022, 25, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.M.L.; Jajou, S.; Stellato, A.C.; Niel, L. Perspectives of Canadian and American Cat Owners on Provision of Uncontrolled Outdoor Access for Owned Domestic Cats. Front. Vet. Sci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.M.; Ong, S.M.; Langat, G.; Bahaman, A.R.; Sharma, R.S.K.; Sumita, S. The Influence of Enclosure Design on Diurnal Activity and Stereotypic Behaviour in Captive Malayan Sun Bears (Helarctos Malayanus). Res. Vet. Sci. 2013, 94, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Warwick, C.; Grant, R.; Steedman, C.; Howell, T.J.; Arena, P.C.; Lambiris, A.J.L.; Nash, A.E.; Jessop, M.; Pilny, A.; Amarello, M.; et al. Getting It Straight: Accommodating Rectilinear Behavior in Captive Snakes—A Review of Recommendations and Their Evidence Base. Animals 2021, 11, 1459. [Google Scholar] [CrossRef] [PubMed]
- Jensvold, M.L.A.; Sanz, C.M.; Fouts, R.S.; Fouts, D.H. Effect of Enclosure Size and Complexity on the Behaviors of Captive Chimpanzees (Pan Troglodytes). J. Appl. Anim. Welf. Sci. 2001, 4, 53–69. [Google Scholar] [CrossRef]
- Webb, S.J.N.; Hau, J.; Schapiro, S.J. Captive Chimpanzee (Pan Troglodytes) Behavior as a Function of Space per Animal and Enclosure Type. Am. J. Primatol. 2018, 80, e22749. [Google Scholar] [CrossRef] [Green Version]
- Philadelphia Zoo Philadelphia Magazine. 2016. Available online: https://www.phillymag.com/sponsor-content/philadelphia-zoo-animal-trails/ (accessed on 12 February 2023).
- Bisgrove, D. Zooscape Ecology: A Conceptual Analysis of Zoos and Landscape Ecology. Landsc. Ecol. 2022, 37, 1733–1745. [Google Scholar] [CrossRef]
- Belyaeva, T.V. Dynamic Architecture. Interaction with City, Nature, Man. IOP Conf. Ser. Mater. Sci. Eng. 2019, 687, 055015. [Google Scholar] [CrossRef] [Green Version]
- Karanouh, A.; Kerber, E. Innovations in Dynamic Architecture. J. Facade Des. Eng. 2015, 3, 185–221. [Google Scholar] [CrossRef] [Green Version]
- Maulana, R. Architecture for Animals: The Expanding Challenges of Sustainable Development. IOP Conf Ser Earth Environ Sci 2018, 195, 012079. [Google Scholar] [CrossRef]
- Sade, C. Visitor Effects on Zoo Animals. Plymouth Stud. Sci. 2013, 6, 423–433. [Google Scholar]
- Fernandez, E.J.; Chiew, S.J. Animal-Visitor Interactions: Effects, Experiences, and Welfare. Anim. Behav. Cogn. 2021, 8, 462–467. [Google Scholar] [CrossRef]
- Hoy, R.A.; Brereton, J.E. Does Observer Presence Modify the Behavior and Enclosure Use of Captive Edwards’ Pheasants? J. Zool. Bot. Gard. 2022, 3, 147–157. [Google Scholar] [CrossRef]
- Suárez, P.; Recuerda, P.; Arias-De-Reyna, L. Behaviour and Welfare: The Visitor Effect in Captive Felids. Anim. Welf. 2017, 26, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, M.B.; Young, R.J. The Different Physical and Behavioural Characteristics of Zoo Mammals That Influence Their Response to Visitors. Animals 2018, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Christakis, D.A.; Ramirez, J.S.B.; Ramirez, J.M. Overstimulation of Newborn Mice Leads to Behavioral Differences and Deficits in Cognitive Performance. Sci. Rep. 2012, 2, 546. [Google Scholar] [CrossRef] [Green Version]
- Townsend, L.; Gee, N.R. Recognizing and Mitigating Canine Stress during Animal Assisted Interventions. Vet. Sci. 2021, 8, 254. [Google Scholar] [CrossRef]
- San Diego Zoo Live Cameras. 2023. Available online: https://zoo.sandiegozoo.org/live-cameras (accessed on 3 March 2023).
- Camstreamer The Melbourne Zoo Is Using QR Codes in Their Live Stream, Forward People from Videos of Sleeping Animals to Other Content. Available online: https://camstreamer.com/resources/melbourne-zoo-pdf (accessed on 3 March 2023).
- Houston Zoo Webcams Houston Zoo. 2023. Available online: https://www.houstonzoo.org/explore/webcams/ (accessed on 3 March 2023).
- Tofield, S.; Coll, R.K.; Vyle, B.; Bolstad, R. Zoos as a Source of Free Choice Learning. Research in Science & Technological Education 2003, 21, 67–99. [Google Scholar] [CrossRef]
- Ivana, G.; Martin, H.; Jitka, Š. Environmental Enrichment in the Awareness of Zoo Visitors and the General Public. Annu Res. Rev. Biol. 2017, 20, 1–5. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Domingues, M.; Ström, E.; Berggren, L. Cognitive Foraging Enrichment (but Not Non-Cognitive Enrichment) Improved Several Longer-Term Welfare Indicators in Bottlenose Dolphins. Animals 2023, 13, 238. [Google Scholar] [CrossRef]
- Gusset, M.; Chin, S.A. Future of Zoo and Aquarium Design; WAZA Executive Office: Gland, Switzerland, 2016. [Google Scholar]
- Krebs, B.L.; Marrin, D.; Phelps, A.; Krol, L.; Watters, J.V. Managing Aged Animals in Zoos to Promote Positive Welfare: A Review and Future Directions. Animals 2018, 8, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Azevedo, C.S.; Cipreste, C.F.; Pizzutto, C.S.; Young, R.J. Review of the Effects of Enclosure Complexity and Design on the Behaviour and Physiology of Zoo Animals. Animals 2023, 13, 1277. https://doi.org/10.3390/ani13081277
de Azevedo CS, Cipreste CF, Pizzutto CS, Young RJ. Review of the Effects of Enclosure Complexity and Design on the Behaviour and Physiology of Zoo Animals. Animals. 2023; 13(8):1277. https://doi.org/10.3390/ani13081277
Chicago/Turabian Stylede Azevedo, Cristiano Schetini, Cynthia Fernandes Cipreste, Cristiane Schilbach Pizzutto, and Robert John Young. 2023. "Review of the Effects of Enclosure Complexity and Design on the Behaviour and Physiology of Zoo Animals" Animals 13, no. 8: 1277. https://doi.org/10.3390/ani13081277