
The Synergistic Effects of the Combination of L-Carnitine and Lycopene on the Lycopene Bioavailability and Duodenal Health of Roosters

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Sample Collection
2.3. Intestinal Morphology Analysis
2.4. Serum Lycopene Levels and Biochemical Indexes
2.5. Total RNA Extraction and mRNA Quantification
2.6. Statistical Analysis
3. Results
3.1. Intestinal Morphology
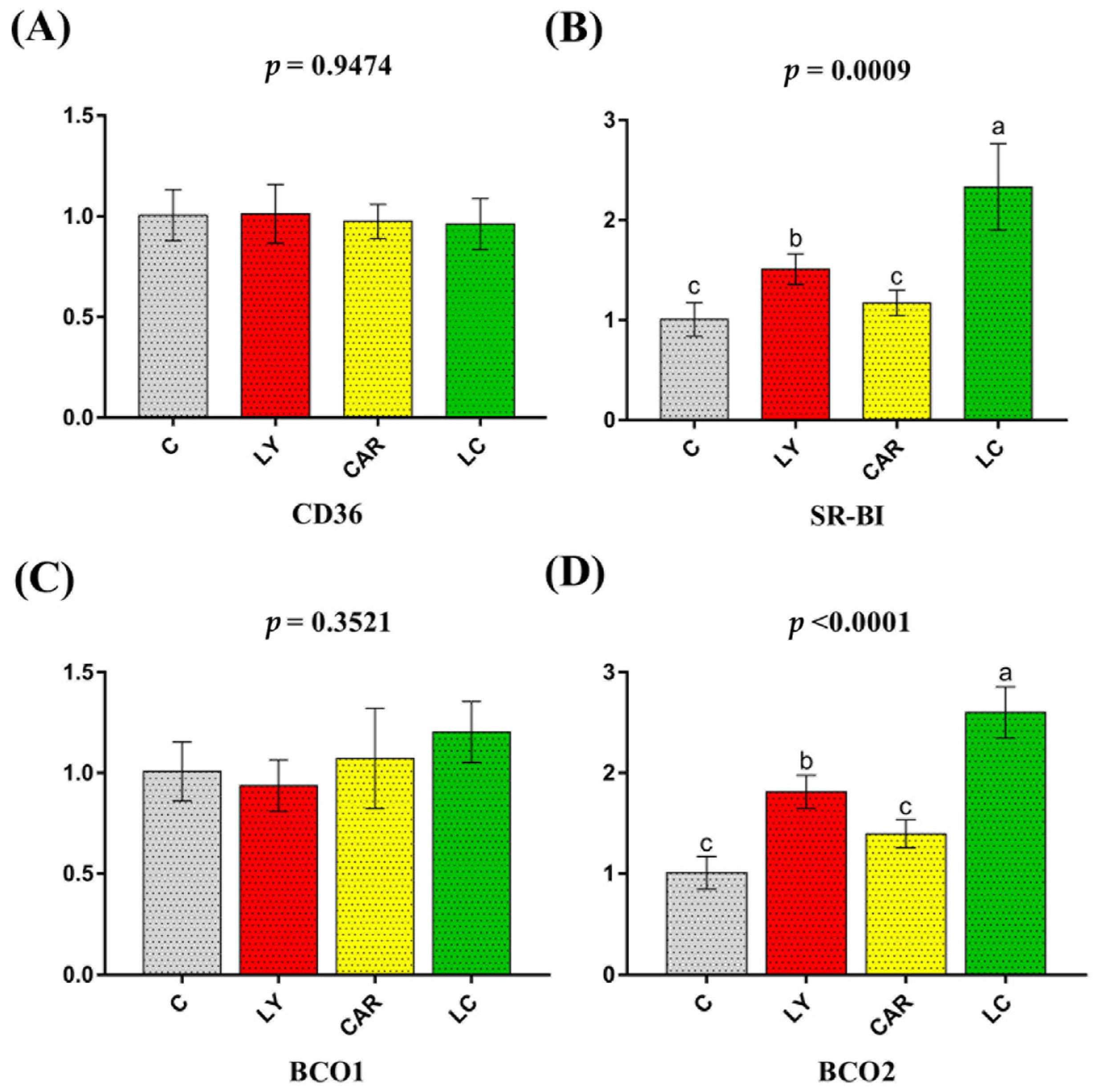
3.2. Relative mRNA Expression of Duodenal Related to Lycopene Uptake
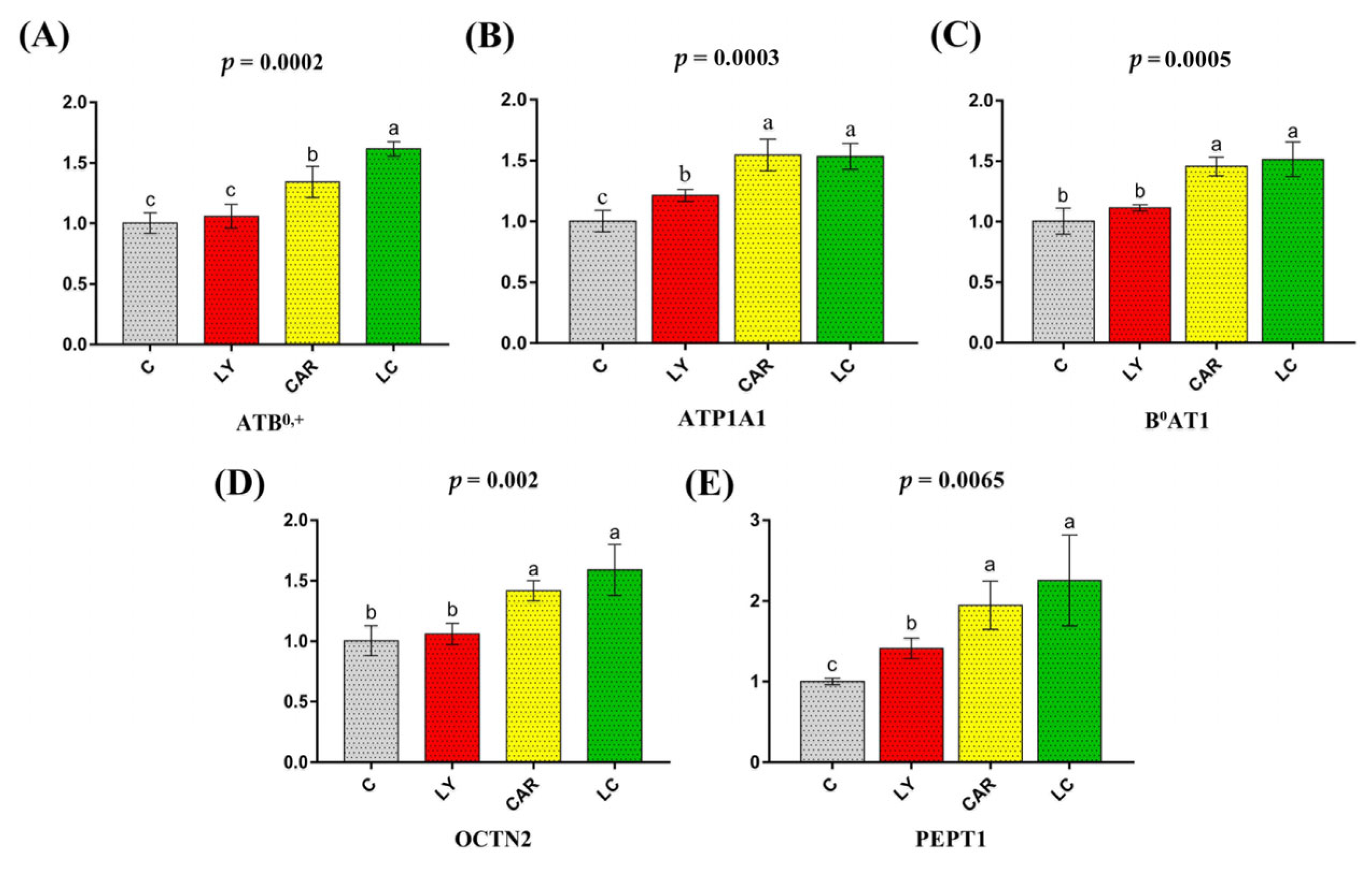
3.3. Relative mRNA Expression of Candidate Nutritional Transport Genes in the Duodenum
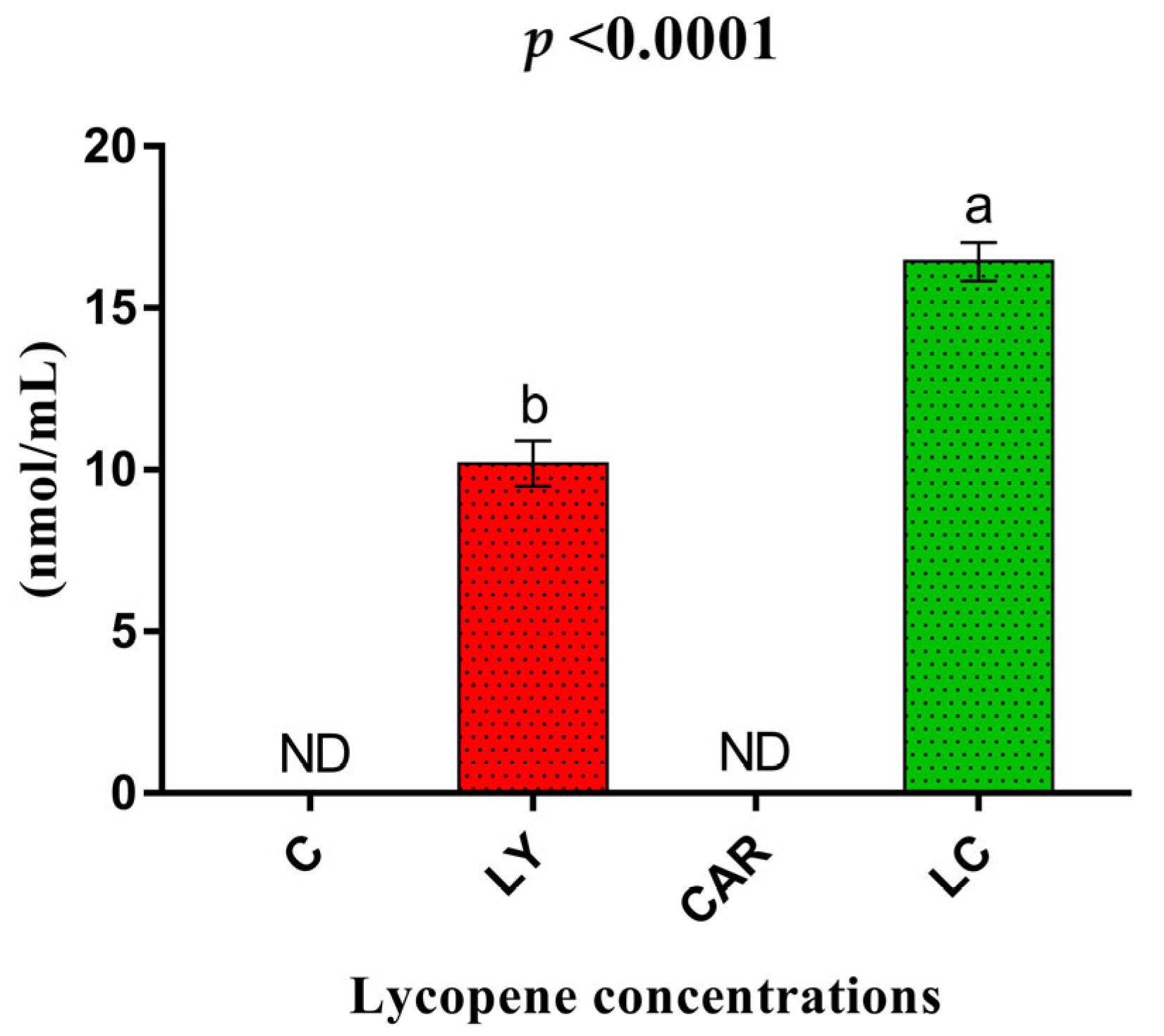
3.4. Serum Lycopene Levels
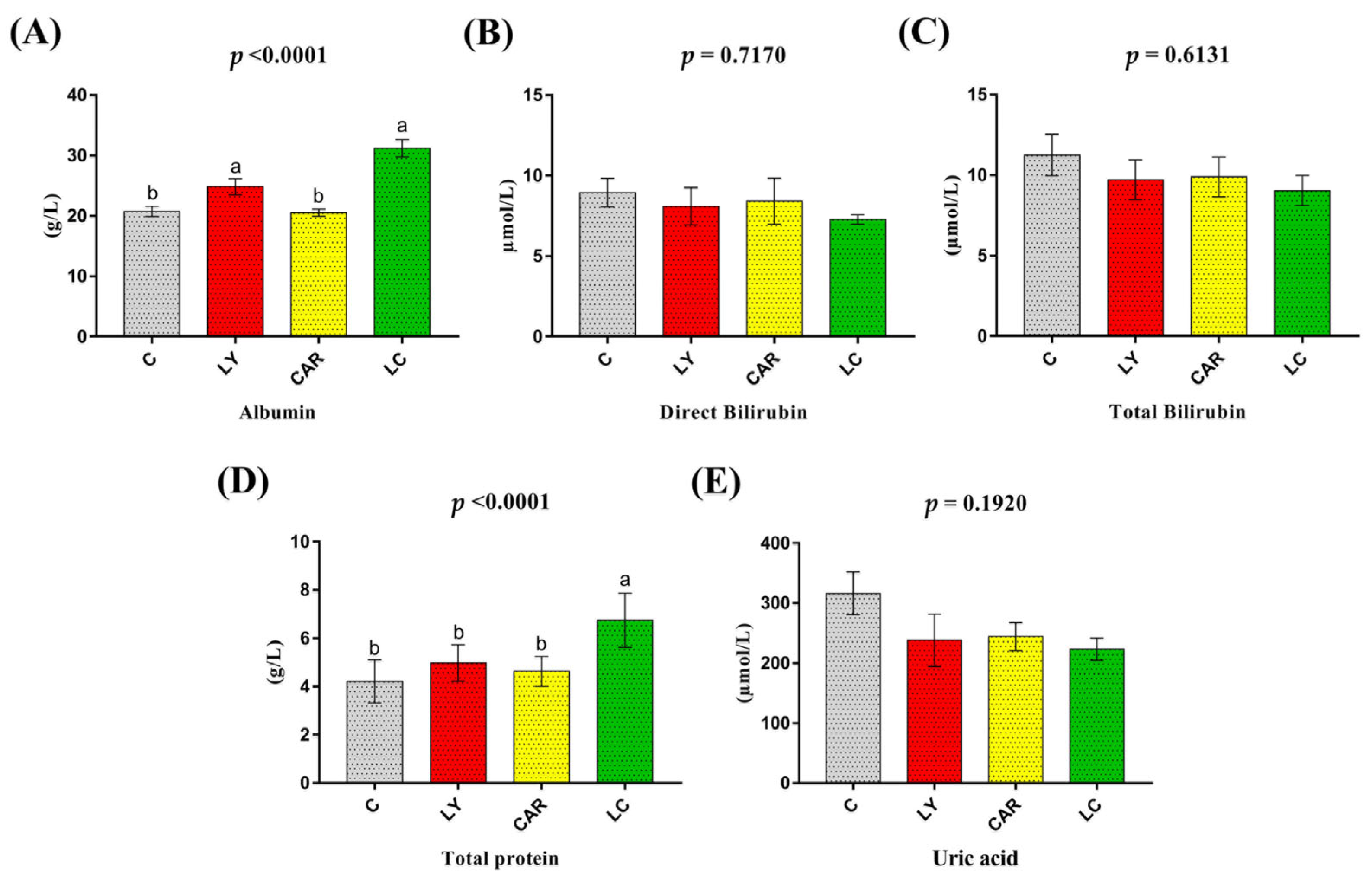
3.5. Serum Biochemical Indexes
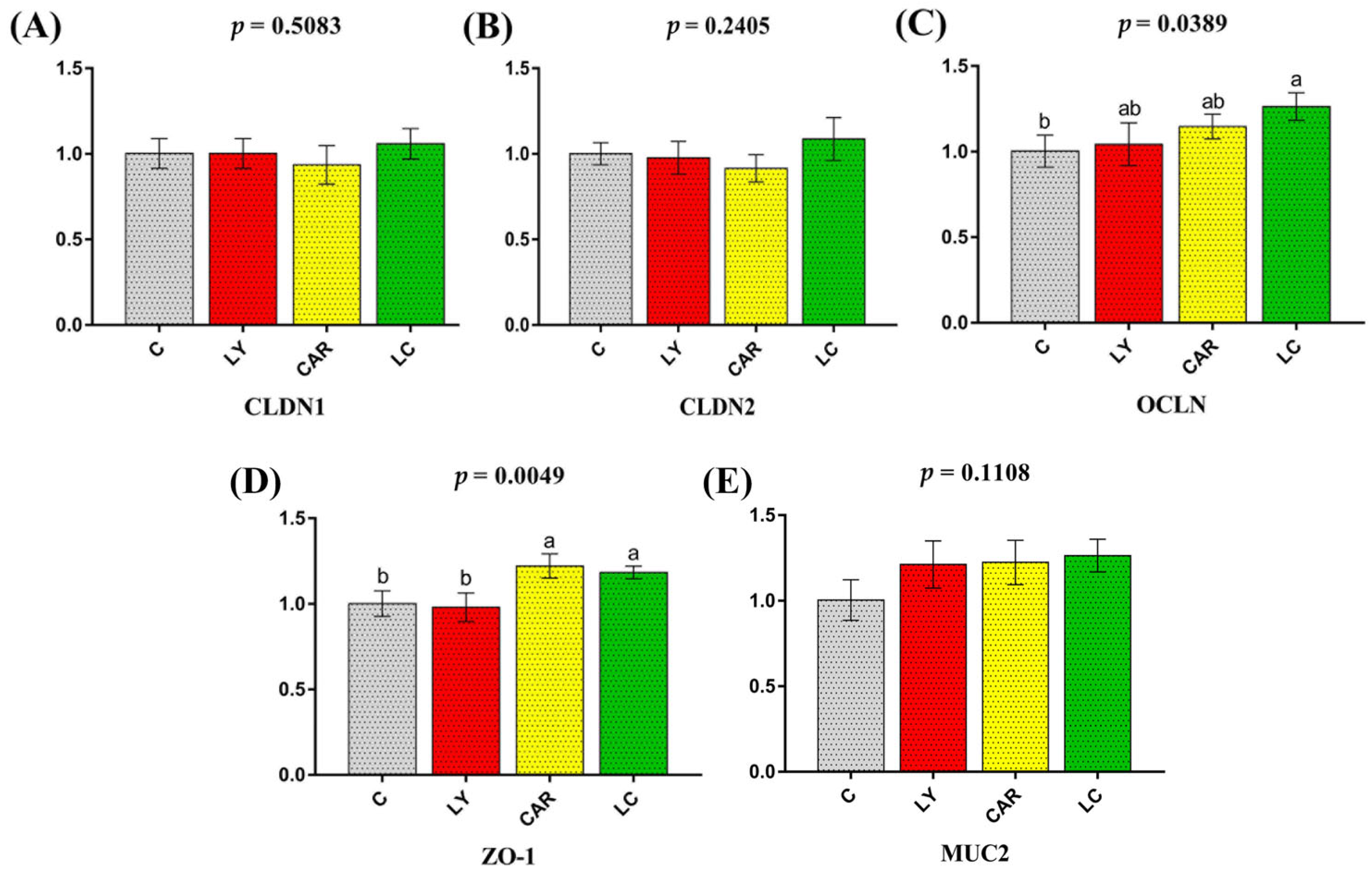
3.6. mRNA Expression of Tight Junction Genes and Mucin in the Duodenum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdel-Moneim, A.M.E.; Shehata, A.M.; Khidr, R.E.; Paswan, V.K.; Ibrahim, N.S.; El-Ghoul, A.A.; Aldhumri, S.A.; Gabr, S.A.; Mesalam, N.M.; Elbaz, A.M.; et al. Nutritional manipulation to combat heat stress in poultry—A comprehensive review. J. Therm. Biol. 2021, 98, 102915. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.; Fisinin, V. Vitagenes in poultry production: Part 2. Nutritional and internal stresses. World’s Poult. Sci. J. 2016, 72, 761–772. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidants in Poultry Nutrition and Reproduction: An Update. Antioxidants 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, Y.; Zhou, Z.; Shu, Y.; Zhai, C.; Zhu, Y.; Gong, S.; Cui, Y.; Wang, J.-F. Chronic unpredictable stress impairs endogenous antioxidant defense in rat brain. Neurosci. Lett. 2015, 584, 208–213. [Google Scholar] [CrossRef]
- Habashy, W.S.; Milfort, M.C.; Adomako, K.; Attia, Y.A.; Rekaya, R.; Aggrey, S.E. Effect of heat stress on amino acid digestibility and transporters in meat-type chickens. Poult. Sci. 2017, 96, 2312–2319. [Google Scholar] [CrossRef]
- Li, Y.; Cai, H.Y.; Liu, G.H.; Dong, X.L.; Chang, W.H.; Zhang, S.; Zheng, A.J.; Chen, G.L. Effects of stress simulated by dexamethasone on jejunal glucose transport in broilers. Poult. Sci. 2009, 88, 330–337. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sakai, M.; Sá, L.; Ferreira, A.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef]
- Ziegler, A.L.; Blikslager, A.T. Impaired intestinal barrier function and relapsing digestive disease: Lessons from a porcine model of early life stress. Neurogastroenterol. Motil. 2017, 29, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Al-Mutary, G.M. Use of antioxidants to augment semen efficiency during liquid storage and cryopreservation in livestock animals: A review. J. King Saud Univ. Sci. 2020, 33, 101226. [Google Scholar] [CrossRef]
- Alnadari, F.; Abdin, M.; Ennab, W.; Mohedein, A.; Nasiru, M.M. Metabolism of Anthocyanins and Modulation of Gut Microbiome in Inflammatory Bowel Disease. J. Food. Chem. Nanotechnol. 2020, 6, 207–217. [Google Scholar] [CrossRef]
- Ennab, W.; Mustafa, S.; Wei, Q.; Lv, Z.; Kavita, N.M.X.; Ullah, S.; Shi, F. Resveratrol protects against restraint stress effects on stomach and spleen in adult male mice. Animals 2019, 9, 736. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, S.; Ennab, W.; Nazar, K.; Wei, Q.; Lv, Z.; Shi, Z.; Shi, F. Positive roles of resveratrol in early development of testicular germ cells against maternal restraint stress in mice. Animals 2020, 10, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Alesci, S.; Manoli, I.; Costello, R.; Coates, P.; Gold, P.W.; Chrousos, G.P.; Blackman, M.R. Carnitine: Lessons from one hundred years of research. In Annual the New York Academy of Sciences; Wiley-Blackwell: Hoboken, NJ, USA, 2004; Volume 1033, pp. ix–xi. [Google Scholar]
- Rebouche, C.J. Carnitine. In Modern Nutrition in Health and Disease, 9th ed.; Shils, M.E., Olson, J.A., Shike, M., Eds.; Williams & Wilkins: Baltimore, MD, USA, 1999; Volume 1, pp. 334–338. [Google Scholar]
- Ribas, G.S.; Vargas, C.R.; Wajner, M. L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene 2014, 533, 469–476. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidant action of carnitine: Molecular mechanisms and practical applications. EC Vet. Sci. 2015, 2, 66–84. [Google Scholar]
- Bohn, T.; Desmarchelier, C.; Dragsted, L.O.; Nielsen, C.S.; Stahl, W.; Rühl, R.; Keijer, J.; Borel, P. Host-related factors explaining interindividual variability of carotenoid bioavailability and tissue concentrations in humans. Mol. Nutr. Food Res. 2017, 61, 1600685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.; Rodríguez-Gironés, M.A.; Barbosa, A.; Costas, M. Donator acceptor map for carotenoids, melatonin and vitamins. J. Phys. Chem. A 2008, 112, 9037–9042. [Google Scholar] [CrossRef]
- Liang, X.; Ma, C.; Yan, X.; Liu, X.; Liu, F. Advances in research on bioactivity, metabolism, stability and delivery systems of lycopene. Trends Food Sci. Technol. 2019, 93, 185–196. [Google Scholar] [CrossRef]
- Bhuvaneswari, V.; Nagini, S. Lycopene: A review of its potential as an anticancer agent. Curr. Med. Chem. Anti-Cancer Agents 2005, 5, 627–635. [Google Scholar] [CrossRef]
- Bramley, P.M. Is lycopene beneficial to human health? Phytochemistry 2000, 54, 233–236. [Google Scholar] [CrossRef]
- Caseiro, M.; Ascenso, A.; Costa, A.; Creagh-Flynn, J.; Johnson, M.; Simões, S. Lycopene in human health. LWT 2020, 127, 109323. [Google Scholar] [CrossRef]
- Gann, P.H.; Ma, J.; Giovannucci, E.; Willett, W.; Sacks, F.M.; Hennekens, C.H.; Stampfer, M.J. Lower prostate cancer risk in men with elevated plasma lycopene levels: Results of a prospective analysis. Cancer Res. 1999, 59, 1225–1230. [Google Scholar]
- Karas, M.; Amir, H.; Fishman, D.; Danilenko, M.; Segal, S.; Nahum, A.; Koifmann, A.; Giat, Y.; Levy, J.; Sharoni, Y. Lycopene interferes with cell cycle progression and insulin-like growth factor I signaling in mammary cancer cells. Nutr. Cancer 2000, 36, 101–111. [Google Scholar] [CrossRef]
- Bárdos, L.; Kiss, Z.; Gregosits, B.; Réthy, K.; Kerti, A.; Szabo, C. Studies on the effects of lycopene in poultry (hen and quail). In International Seminar Animal Health; International Society for Animal Hygiene: Warsaw, Poland, 2005. [Google Scholar]
- Murphy, M.P. Antioxidants as therapies: Can we improve on nature? Free Radic. Biol. Med. 2014, 66, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Pashtetsky, V.; Ostapchuk, P.; Il’yazov, R.; Zubochenko, D.; Kuevda, T. Use of antioxidants in poultry farming. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019. [Google Scholar]
- Surai, P.F. Natural antioxidants in poultry nutrition: New developments. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; World Poultry Science Association: Beekbergen, The Netherlands, 2007. [Google Scholar]
- Amini, F.G.; Rafieian-Kopaei, M.; Nematbakhsh, M.; Baradaran, A.; Nasri, H. Ameliorative effects of metformin on renal histologic and biochemical alterations of gentamicin-induced renal toxicity in Wistar rats. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2012, 17, 621. [Google Scholar]
- Bruckbauer, A.; Zemel, M.B. Synergistic effects of metformin, resveratrol, and hydroxymethylbutyrate on insulin sensitivity. Diabetes. Diabetes Metab. Syndr. Obes. Targets Ther. 2013, 6, 93. [Google Scholar]
- Morales, A.I.; Detaille, D.; Prieto, M.; Puente, A.; Briones, E.; Arévalo, M.; Leverve, X.; López-Novoa, J.M.; El-Mir, M.-Y. Metformin prevents experimental gentamicin-induced nephropathy by a mitochondria-dependent pathway. Kidney Int. 2010, 77, 861–869. [Google Scholar] [CrossRef]
- Rafieian-Kopaei, M.; Baradaran, A.; Merrikhi, A.; Nematbakhsh, M.; Madihi, Y.; Nasri, H. Efficacy of Co-administration of Garlic Extract and Metformin for Prevention of Gentamicin–Renal Toxicity in Wistar Rats: A Biochemical Study. Int. J. Prev. Med. 2013, 4, 258–264. [Google Scholar]
- Simmons, C.F., Jr.; Bogusky, R.T.; Humes, H.D. Inhibitory effects of gentamicin on renal mitochondrial oxidative phosphorylation. J. Pharmacol. Exp. Ther. 1980, 214, 709–715. [Google Scholar] [PubMed]
- Surai, P.F. Selenium-Vitamin E Interactions: Does 1+ 1 Equal More Than 2. In Nutritional Biotechnology in the Feed and Food Industries; Lyons, T.P., Jacques, K.A., Eds.; Nottingham University Press: Nottingham, UK, 2003; pp. 47–51. [Google Scholar]
- Abdollahi, M.; Tabatabaei-Malazy, O.; Larijani, B. A systematic review of in vitro studies conducted on effect of herbal products on secretion of insulin from Langerhans islets. J. Pharm. Pharm. Sci. 2012, 15, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Tabatabaei-Malazy, O.; Larijani, B.; Abdollahi, M. A novel management of diabetes by means of strong antioxidants’ combination. J. Med. Hypotheses Ideas 2013, 7, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Gentile, V.; Vicini, P.; Prigiotti, G.; Koverech, A.; Di Silverio, F. Preliminary observations on the use of propionyl-L-carnitine in combination with sildenafil in patients with erectile dysfunction and diabetes. Curr. Med. Res. Opin. 2004, 20, 1377–1384. [Google Scholar] [CrossRef]
- Sleem, M.; Taye, A.; El-Moselhy, M.A.; Mangoura, S.A. Combination therapy with losartan and L-carnitine protects against endothelial dysfunction of streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2014, 744, 10–17. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H.; Zhao, D.; Ding, D.; Sun, M.; Kou, L.; Luo, C.; Zhang, D.; Yi, X.; Dong, J.; et al. Combination of l-carnitine with lipophilic linkage-donating gemcitabine derivatives as intestinal novel organic cation transporter 2-targeting oral prodrugs. J. Med. Chem. 2017, 60, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Feng, J.; Zhang, Q.; Wu, W.; Mo, H.; Huang, L.; Zhang, W. l-Carnitine conjugated chitosan-stearic acid polymeric micelles for improving the oral bioavailability of paclitaxel. Drug Deliv. 2020, 27, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuoti Al-Rekabi, B.K.; Al-Shwilly, H.A. Comparative Study on the Protective Effect of L-Carnitine in Combination with Beta-Carotene against Overdose Acetaminophen Induced Hepatotoxicity and Nephrotoxicity in Adults Male Rats. Indian J. Public Health Res. Dev. 2019, 10, 897–901. [Google Scholar] [CrossRef]
- Bernard, A.; Rigault, C.; Mazue, F.; Le Borgne, F.; Demarquoy, J. L-carnitine supplementation and physical exercise restore age-associated decline in some mitochondrial functions in the rat. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2008, 63, 1027–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, W.; Noh, S.K.; Owen, K.Q.; Koo, S.I. Dietary L-carnitine enhances the lymphatic absorption of fat and α-tocopherol in ovariectomized rats. J. Nutr. 2005, 135, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, K.; Pittaway, J.K.; Ball, M.J. Effects of olive oil and tomato lycopene combination on serum lycopene, lipid profile, and lipid oxidation. Nutrition 2006, 22, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Assis, R.P.; Arcaro, C.A.; Gutierres, V.O.; Oliveira, J.O.; Costa, P.I.; Baviera, A.M.; Brunetti, I.L. Combined effects of curcumin and lycopene or bixin in yoghurt on inhibition of LDL oxidation and increases in HDL and paraoxonase levels in streptozotocin-diabetic rats. Int. J. Mol. Sci. 2017, 18, 332. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Feng, T.; Dai, G.; Wang, Y.; Zhu, H.; Hu, J. Lycopene and alpha-lipoic acid improve semen antioxidant enzymes activity and cashmere goat sperm function after cryopreservation. Cryobiology 2018, 84, 27–32. [Google Scholar] [CrossRef]
- Shi, J.; Qu, Q.; Kakùda, Y.; Yeung, D.; Jiang, Y. Stability and synergistic effect of antioxidative properties of lycopene and other active components. Crit. Rev. Food Sci. Nutr. 2005, 44, 559–573. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Poultry; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Ghods-Alavi, B.; Samie, H.; Jahanian, R. Effects of supplementary dietary L-carnitine on performance and egg quality of laying hens fed diets different in fat level. Ital. J. Anim. Sci. 2010, 9, e3. [Google Scholar] [CrossRef] [Green Version]
- Sahin, K.; Yenice, E.; Tuzcu, M.; Orhan, C.; Mizrak, C.; Ozercan, I.H.; Sahin, N.; Yilmaz, B.; Bilir, B.; Ozpolat, B.; et al. Lycopene protects against spontaneous ovarian cancer formation in laying hens. J. Cancer Prev. 2018, 23, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekeli, A. Effects of L-carnitine supplementation on ascites syndrome in the broilers grown at high altitude. Rev. MVZ Córdoba 2019, 24, 7127–7136. [Google Scholar]
- Tufan, T.; Arslan, C.; Durna, Ö.; Önk, K.; Sari, M.; Erman, H. Effects of chito-oligosaccharides and L-carnitine supplementation in diets for Japanese quails on performance, carcass traits and some blood parameters. Arq. Bras. Med. Vet. Zootec. 2015, 67, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Sahin, K.; Onderci, M.; Sahin, N.; Gursu, M.F.; Khachik, F.; Kucuk, O. Effects of lycopene supplementation on antioxidant status, oxidative stress, performance and carcass characteristics in heat-stressed Japanese quail. J. Therm. Biol. 2006, 31, 307–312. [Google Scholar] [CrossRef]
- Mustafa, S.; Wei, Q.; Ennab, W.; Lv, Z.; Nazar, K.; Siyal, F.A.; Rodeni, S.; Kavita, N.M.X.; Shi, F. Resveratrol ameliorates testicular histopathology of mice exposed to restraint stress. Animals 2019, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Ngekure MX, K.; Jiang, J.; Enayatullah, H.; Ennab, W.; Mustafa, S.; Rodeni, S.; Wei, Q.; Shi, F. Sweet taste receptor agonists alter ovarian functions and ovarian cycles in aged mice. Reprod. Biol. 2019, 19, 230–236. [Google Scholar] [CrossRef]
- Ullah, S.; Sheeraz, M.; Wael, E.; Jan, M.; Shafiq, M.; Kavita, N.M.X.; Lü, Z.-P.; Mao, D.-G.; Shi, F.-X. A protective role of resveratrol against the effects of immobilization stress in corpora lutea of mice in early pregnancy. J. Integr. Agric. 2020, 19, 1857–1866. [Google Scholar] [CrossRef]
- Rasband, W.S. Imagej, US National Institutes of Health, Bethesda, Maryland, USA. Available online: https://imagej.nih.gov/ij/download.html (accessed on 1 November 2022).
- Liu, A.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; He, J.; Yan, H.; Zheng, P.; Yu, J.; et al. Effects of dietary lycopene supplementation on intestinal morphology, antioxidant capability and inflammatory response in finishing pigs. Anim. Biotechnol. 2021, 33, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Lycopene: A biologically important carotenoid for humans? Arch. Biochem. Biophys. 1996, 336, 1–9. [Google Scholar] [CrossRef]
- Mezbani, A.; Kavan, B.P.; Kiani, A.; Masouri, B. Effect of dietary lycopene supplementation on growth performance, blood parameters and antioxidant enzymes status in broiler chickens. Livest. Res. Rural. Dev. 2019, 31, 1. [Google Scholar]
- An, B.-K.; Choo, W.D.; Kang, C.W.; Lee, J.; Lee, K.W. Effects of dietary lycopene or tomato paste on laying performance and serum lipids in laying hens and on malondialdehyde content in egg yolk upon storage. J. Poult. Sci. 2019, 56, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Luo, Y.; Ma, L.; Hu, X.; Simal-Gandara, J.; Wang, L.S.; Bajpai, V.K.; Xiao, J.; Chen, F. Recent trends and advances in the epidemiology, synergism, and delivery system of lycopene as an anti-cancer agent. Semin. Cancer Biol. 2021, 73, 331–346. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Karimzadeh, I.; Sagheb, M.M.; Khalili, H. The renal safety of L-carnitine, L-arginine, and glutamine in athletes and bodybuilders. J. Ren. Nutr. 2019, 29, 221–234. [Google Scholar] [CrossRef] [PubMed]
- McClain, R.M.; Bausch, J. Summary of safety studies conducted with synthetic lycopene. Regul. Toxicol. Pharmacol. 2003, 37, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, V.; Passantino, L.; Perillo, A.; Lopresti, G.; Passantino, A.; Khan, R.U.; Tufarelli, V. Productive performance and histological features of intestinal mucosa of broiler chickens fed different dietary protein levels. Poult. Sci. 2012, 91, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.-E. Review on chicken intestinal villus histological alterations related with intestinal function. J. Poult. Sci. 2002, 39, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Azizi-Chekosari, M.; Bouyeh, M.; Seidavi, A.; Ventura, M.R. Effect of dietary supplementation with L-Carnitine and fenofibrate on broiler chickens. S. Afr. J. Anim. Sci. 2021, 51, 587–603. [Google Scholar] [CrossRef]
- Sun, B.; Chen, C.; Wang, W.; Ma, J.; Xie, Q.; Gao, Y.; Chen, F.; Zhang, X.; Bi, Y. Effects of lycopene supplementation in both maternal and offspring diets on growth performance, antioxidant capacity and biochemical parameters in chicks. J. Anim. Physiol. Anim. Nutr. 2015, 99, 42–49. [Google Scholar] [CrossRef]
- Li, J.; Qian, H.; Pi, F.; Wang, B.-X. Bioavailability evaluation of the intestinal absorption and liver accumulation of torularhodin using a rat postprandial model. Food Funct. 2022, 13, 5946–5952. [Google Scholar] [CrossRef]
- Lowe, G.M.; Graham, D.L.; Young, A.J. Lycopene: Chemistry, Metabolism, and Bioavailability. In Lycopene and Tomatoes in Human Nutrition and Health; CRC Press: Boca Raton, FL, USA, 2018; pp. 1–20. [Google Scholar]
- Arballo, J.; Amengual, J.; Erdman, J.W., Jr. Lycopene: A Critical Review of Digestion, Absorption, Metabolism, and Excretion. Antioxidants 2021, 10, 342. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E.; Klein, A.; Bietrix, F.; Gleize, B.; Malezet-Desmoulins, C.; Schneider, M.; Margotat, A.; Lagrost, L.; Collet, X.; Borel, P. Scavenger receptor class B type I (SR-BI) is involved in vitamin E transport across the enterocyte. J. Biol. Chem. 2006, 281, 4739–4745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.E.; Erdman, J.W., Jr.; Clinton, S.K. Complex interactions between dietary and genetic factors impact lycopene metabolism and distribution. Arch. Biochem. Biophys. 2013, 539, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, M.; Landrier, J.; Reboul, E.; Ghiringhelli, O.; Comera, C.; Collet, X.; Borel, P. Lycopene absorption in human intestinal cells and in mice involves scavenger receptor class B type I but not Niemann-Pick C1-like 1. J. Nutr. 2008, 138, 1432–1436. [Google Scholar] [CrossRef] [Green Version]
- Reboul, E.; Borel, P. Proteins involved in uptake, intracellular transport and basolateral secretion of fat-soluble vitamins and carotenoids by mammalian enterocytes. Prog. Lipid Res. 2011, 50, 388–402. [Google Scholar] [CrossRef]
- Böhmer, C.; Bröer, A.; Munzinger, M.; Kowalczuk, S.; Rasko, J.E.; Lang, F.; Bröer, S. Characterization of mouse amino acid transporter B0AT1 (slc6a19). Biochem. J. 2005, 389, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Nałęcz, K.A. Amino acid transporter SLC6A14 (ATB0,+)—A target in combined anti-cancer therapy. Front. Cell Dev. Biol. 2020, 8, 594464. [Google Scholar] [CrossRef]
- Srinivas, S.R.; Prasad, P.D.; Umapathy, N.S.; Ganapathy, V.; Shekhawat, P.S. Transport of butyryl-l-carnitine, a potential prodrug, via the carnitine transporter OCTN2 and the amino acid transporter (ATB0,+). Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G1046–G1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, T.; Tavraz, N.N.; Junghans, C. ATP1A2 mutations in migraine: Seeing through the facets of an ion pump onto the neurobiology of disease. Front. Physiol. 2016, 7, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigault, C.; Bernard, A.; Georges, B.; Kandel, A.; Pfützner, E.; Le Borgne, F.; Demarquoy, J. Extracellular ATP increases L-carnitine transport and content in C2C12 cells. Pharmacology 2008, 81, 246–250. [Google Scholar] [CrossRef]
- Capecchi, P.L.; Laghi Pasini, F.; Ouartarolo, E.; Di Perri, T. Carnitines increase plasma levels of adenosine and ATP in humans. Vasc. Med. 1997, 2, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, H. Molecular and integrative physiology of intestinal peptide transport. Annu. Rev. Physiol. 2004, 66, 361. [Google Scholar] [CrossRef]
- Miska, K.; Fetterer, R. The mRNA expression of amino acid and sugar transporters, aminopeptidase, as well as the di-and tri-peptide transporter PepT1 in the intestines of Eimeria infected broiler chickens. Poult. Sci. 2017, 96, 465–473. [Google Scholar] [CrossRef]
- Sweadner, K.J.; Arystarkhova, E.; Penniston, J.T.; Swoboda, K.J.; Brashear, A.; Ozelius, L.J. Genotype-structure-phenotype relationships diverge in paralogs ATP1A1, ATP1A2, and ATP1A3. Neurol. Genet. 2019, 5, e303. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Liu, S.; Jamal, T.; Ding, T.; Qi, L.; Lv, Z.; Yu, D.; Shi, F. Effects of dietary sweeteners supplementation on growth performance, serum biochemicals, and jejunal physiological functions of broiler chickens. Poult. Sci. 2020, 99, 3948–3958. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.; Drake, V.J.; Delage, B.; Johnson, E.J.; Mayer, J. Carotenoids. α-Carotene, β-Carotene, β-Cryptoxanthin, Lycopene, Lutein, and Zeaxanthin; Micronutrient Information Center, Linus Pauling Institute, Oregon State University: Corvallis, OR, USA, 2016. [Google Scholar]
- İkiz, Ö.; Kahramansoy, N.; Erkol, H.; Koçoğlu, E.; Fırat, T. Effects of lycopene in intestinal ischemia reperfusion injury via intestinal immunoglobulin A. J. Surg. Res. 2021, 267, 63–70. [Google Scholar] [CrossRef]
- Gurer, H.; Ercal, N. Can antioxidants be beneficial in the treatment of lead poisoning? Free Radic. Biol. Med. 2000, 29, 927–945. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Meng, Q.; He, S.; Qu, Z.; Nie, S.; Shi, B.; Shan, A. Effects of l-carnitine in the distillers dried grains with solubles diet of sows on reproductive performance and antioxidant status of sows and their offspring. Animal 2019, 13, 1448–1457. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | 20 to 28 Weeks |
|---|---|
| Corn | 55.86 |
| Soybean meal | 19.10 |
| Soybean oil | 4.12 |
| Wheat bran | 15.12 |
| NaCL | 0.35 |
| DL-methionine | 0.15 |
| Limestone | 3.50 |
| Dicalcium phosphate | 1.52 |
| Mineral premix | 0.30 |
| Vitamin premix | 0.02 |
| Choline chloride (50%) | 0.23 |
| Total | 100 |
| Calculation of nutrients | |
| Metabolizable energy MC/kg | 12.05 |
| Crude protein | 14.5 |
| Calcium | 2.5 |
| Methionine | 0.32 |
| Lysine | 0.7 |
| Available phosphorus | 0.5 |
| Gene Name | Sequences | Amplicon Size (bp) | Accession Number |
|---|---|---|---|
| CLDN1 | Forward: GAGGATGACCAGGTGAAGAAGA Reverse: GCTGATCCAAACTCAAATCTGGTG | 126 | NM_001013611.2 |
| CLDN2 | Forward: ATCTCCAGCCATCTCTGTAACC Reverse: TCTCCCGCACGTTTACCTTT | 150 | NM_001277622.1 |
| OCLN | Forward: AGCCCTCAATACCAGGATGTG Reverse: TGCTTCTTGCTTTGGTAGTCTG | 83 | NM_205128.1 |
| ZO-1 | Forward: AGGAGACCAAATTCTCAGGGTTA Reverse: AGACTCAACAATGCGACGATAAA | 151 | XM_040706827.1 |
| MUC2 | Forward: TACTCTTACCACCATAGTTACCAC Reverse: ATCAAACCACTCAGACCAATCAC | 88 | XM_040701667.1 |
| ATP1A1 | Forward: TTGAGCACTTTATCCACCTCATC Reverse: AGTGTCAGACATACCGTTACAG | 175 | NM_205521.1 |
| B0AT1 | Forward: TTTGGAACCCTAAATACGAGGAAT Reverse: CAATAATAGCATAGACCCAGCCA | 75 | XM_419056.7 |
| ATB0,+ | Forward: ATCGTCTACATTTACGGAGGAAAC Reverse: CCCATGTTGGATATAGCACTGA | 196 | XM_015278436.3 |
| PEPT1 | Forward: CAGACAGTCAACATCACTATGGG Reverse: GTCACATCTCCAGAACACTCATTA | 203 | NM_204365.1 |
| OCTN2 | Forward: GATTGAGGTTCCAGCCTACATTA Reverse: TAGATAGGGCACGAAGATGTGAA | 143 | NM_001045828.1 |
| CD36 | Forward: GGACCTTACACATACAGGGTCAG Reverse: GCATGTACGATATTGTGCCATTAG | 79 | NM_001030731.1 |
| SR-BI | Forward: CAATGTTCATCTCCCACCCTC Reverse: GAATCTTCCCTGTTTGAAGGATACC | 204 | XM_415106.5 |
| BCO1 | Forward: TGGAGACACTAGATAAGGTAGAC Reverse: TGGTACAGAGGAAGGGATCTTA | 164 | NM_001364902.1 |
| BCO2 | Forward: TTGAGTTTGGTGAGGAGAAATACA Reverse: TGTGCTGGTTGTTAATCAGGTAG | 138 | XM_417929.7 |
| β-actin | Forward: CATGGATGATGATATTGCTGCG Reverse: TACCAACCATCACACCCTGAT | 140 | NM_205518.1 |
| Items | Experimental Groups | |||||
|---|---|---|---|---|---|---|
| Duodenum | C | LY | CAR | LC | SEM | p-Value |
| VH (µm) | 952.7 c | 1054 b | 1027 b | 1141 a | 14.5 | <0.0001 |
| CD (µm) | 165.1 | 149.3 | 156.6 | 160.1 | 4.02 | 0.0955 |
| VCR (µm) | 5.787 b | 7.064 a | 6.574 a | 7.173 a | 0.16 | <0.001 |
| Jejunum | ||||||
| VH (µm) | 980.4 d | 1141 b | 1066 c | 1211 a | 15.24 | <0.0001 |
| CD (µm) | 157.6 | 158.5 | 148 | 155.6 | 4.69 | 0.4162 |
| VCR (µm) | 6.25 b | 7.22 a | 7.22 a | 7.824 a | 0.22 | <0.0008 |
| Ileum | ||||||
| VH (µm) | 777.5 | 837.9 | 829.6 | 834.2 | 14.65 | 0.0360 |
| CD (µm) | 124.3 | 125.7 | 122.7 | 121 | 4.87 | 0.9159 |
| VCR (µm) | 6.295 | 6.705 | 6.827 | 6.925 | 0.24 | 0.3053 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ennab, W.; Ye, N.; Wu, H.; Ullah, S.; Hadi, T.; Bassey, A.P.; Mustafa, S.; Jiang, J.; Wei, Q.; Shi, F. The Synergistic Effects of the Combination of L-Carnitine and Lycopene on the Lycopene Bioavailability and Duodenal Health of Roosters. Animals 2023, 13, 1274. https://doi.org/10.3390/ani13081274
Ennab W, Ye N, Wu H, Ullah S, Hadi T, Bassey AP, Mustafa S, Jiang J, Wei Q, Shi F. The Synergistic Effects of the Combination of L-Carnitine and Lycopene on the Lycopene Bioavailability and Duodenal Health of Roosters. Animals. 2023; 13(8):1274. https://doi.org/10.3390/ani13081274
Chicago/Turabian StyleEnnab, Wael, Nanwei Ye, Haoze Wu, Saif Ullah, Tavakolikazerooni Hadi, Anthony Pius Bassey, Sheeraz Mustafa, Jingle Jiang, Quanwei Wei, and Fangxiong Shi. 2023. "The Synergistic Effects of the Combination of L-Carnitine and Lycopene on the Lycopene Bioavailability and Duodenal Health of Roosters" Animals 13, no. 8: 1274. https://doi.org/10.3390/ani13081274